A Newly Isolated Alcaligenes faecalis ANSA176 with the Capability of Alleviating Immune Injury and Inflammation through Efficiently Degrading Ochratoxin A

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
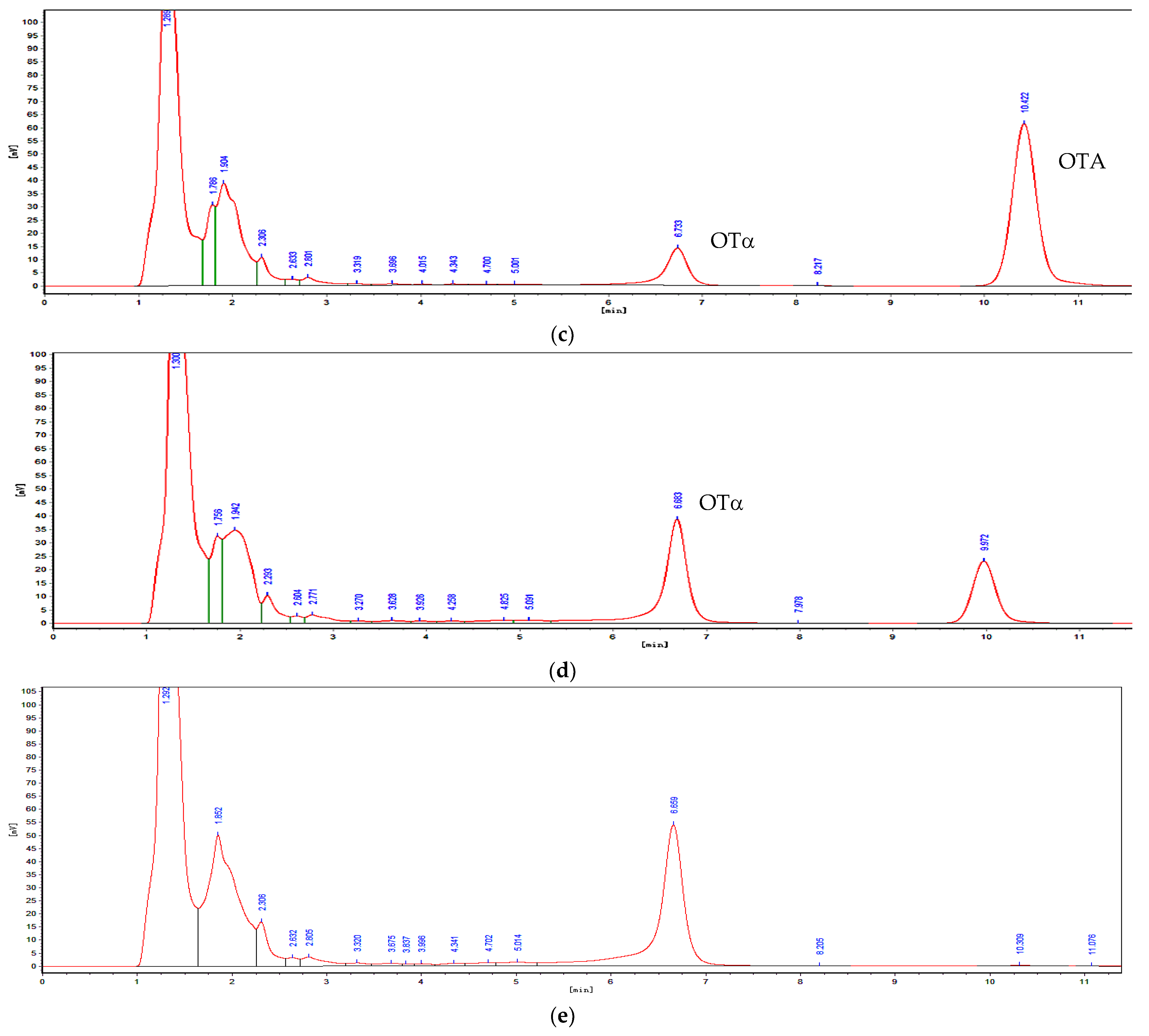
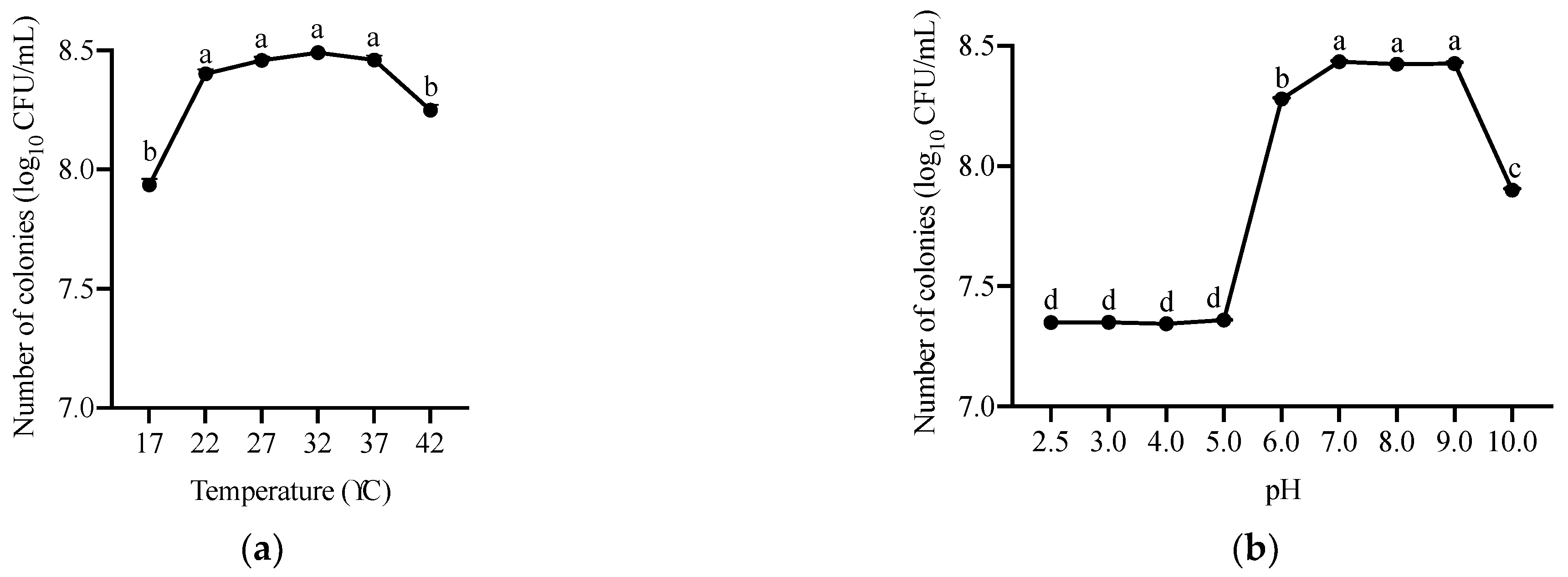
2.1. Isolation and Characterization of OTA-Degrading Strain
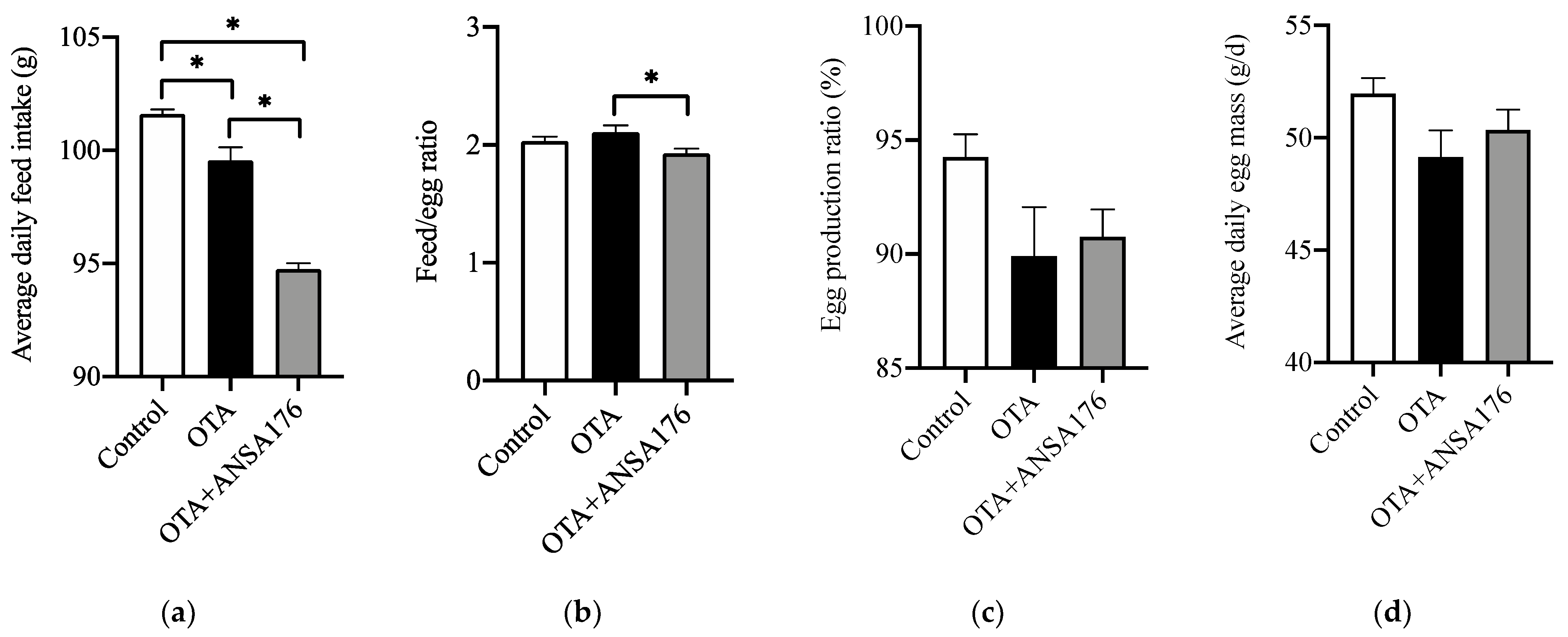
2.2. Protection of A. faecalis ANSA176 against OTA-Induced Damages in Laying Hens
2.2.1. Production Performance in Laying Hens
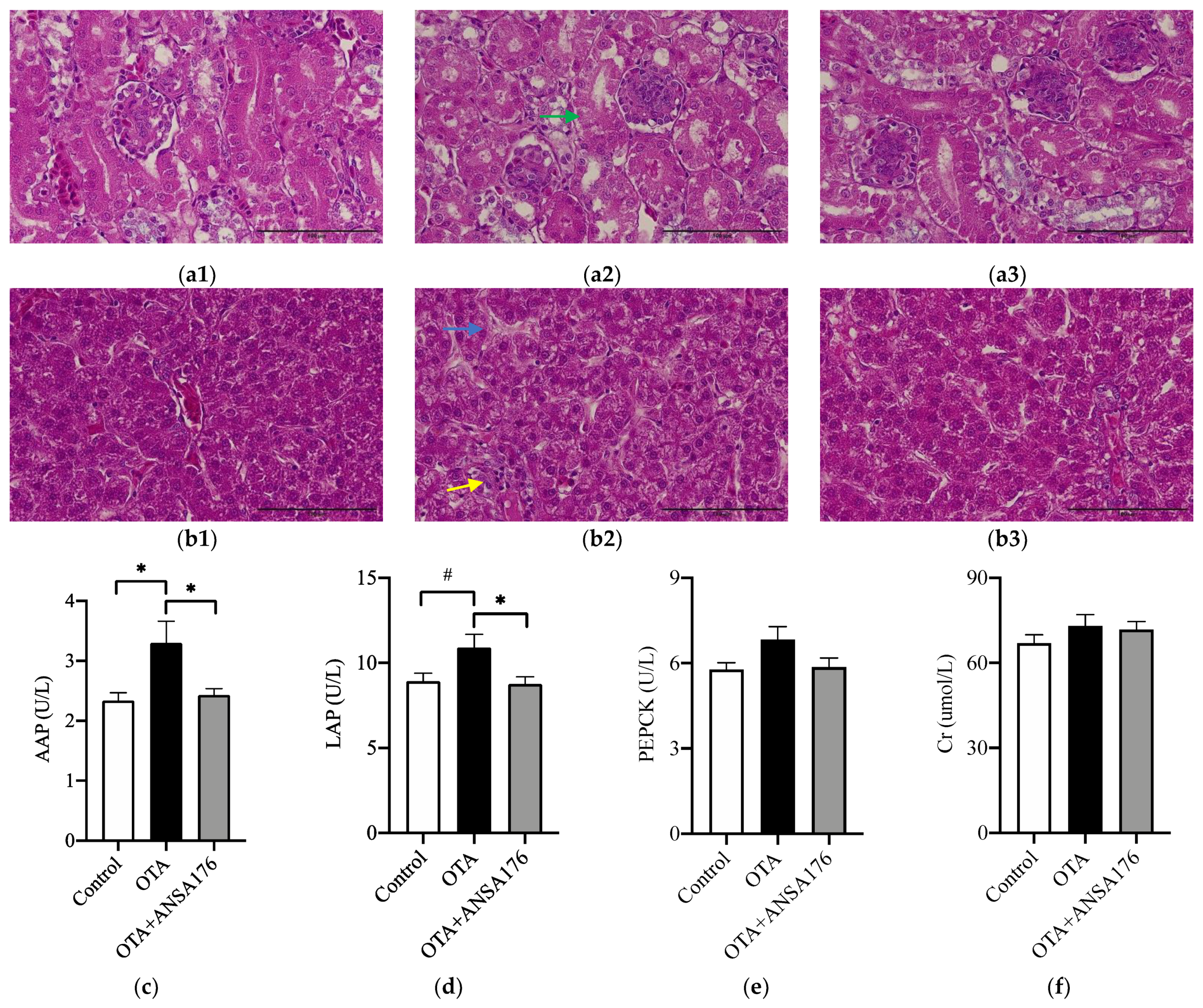
2.2.2. Kidney and Liver Damage Related Parameters in Laying Hens
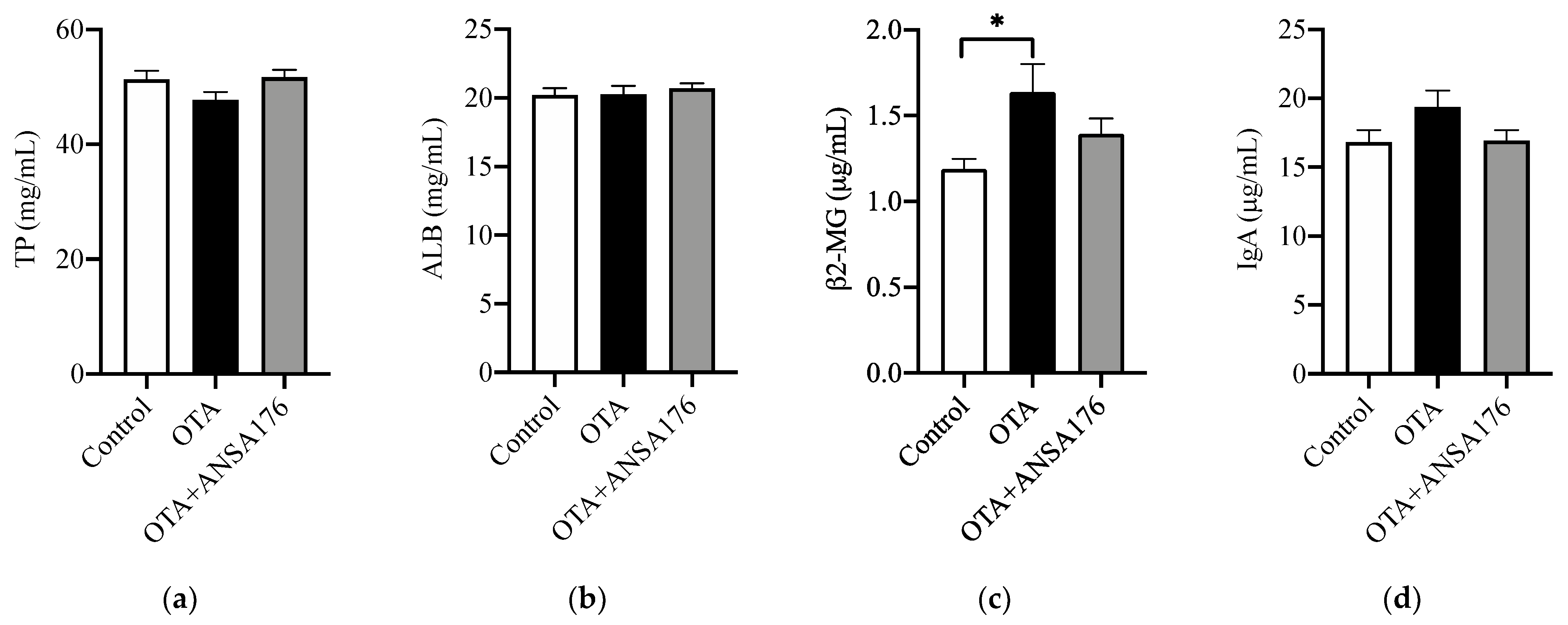
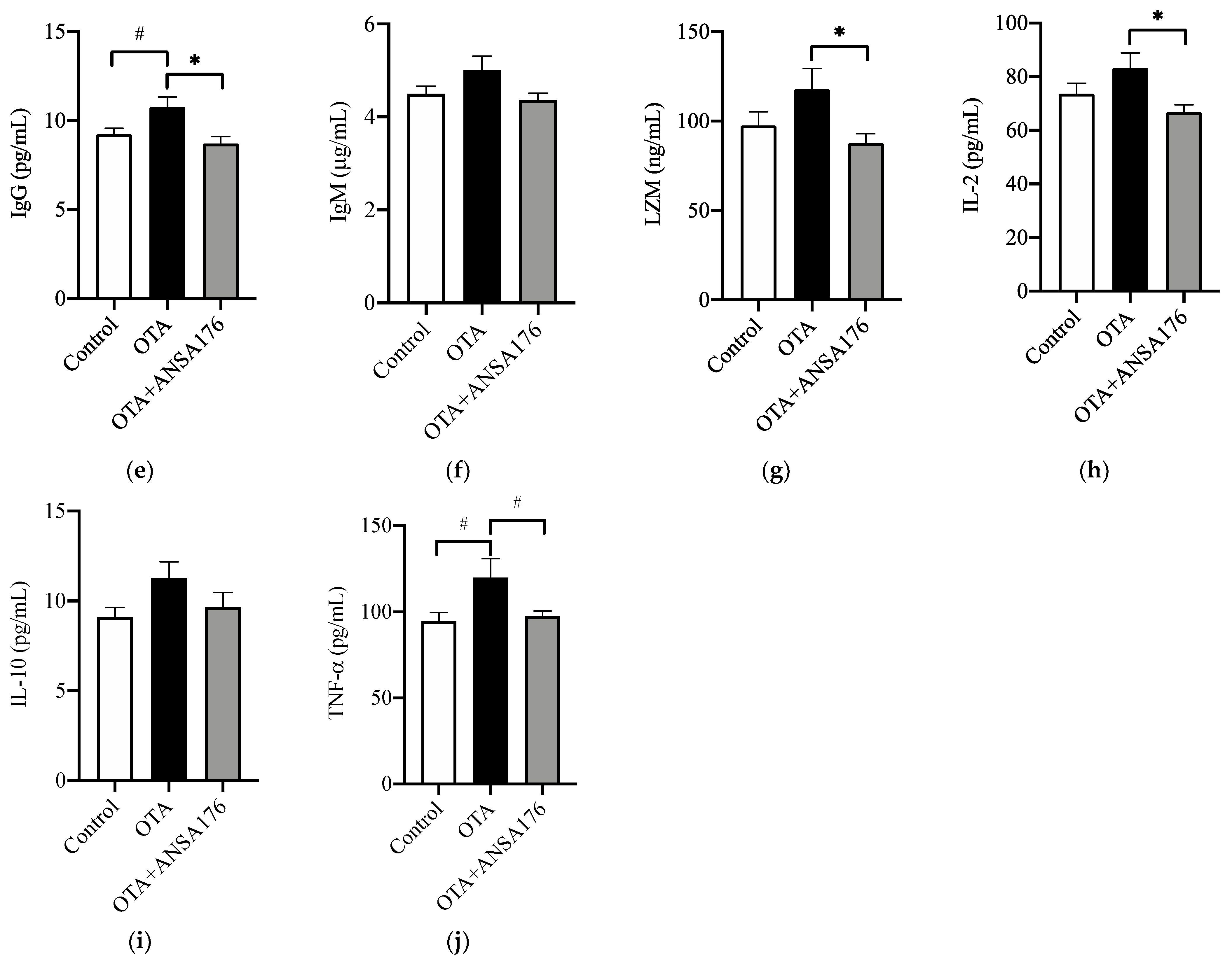
2.2.3. Immune and Inflammatory Response in Laying Hens
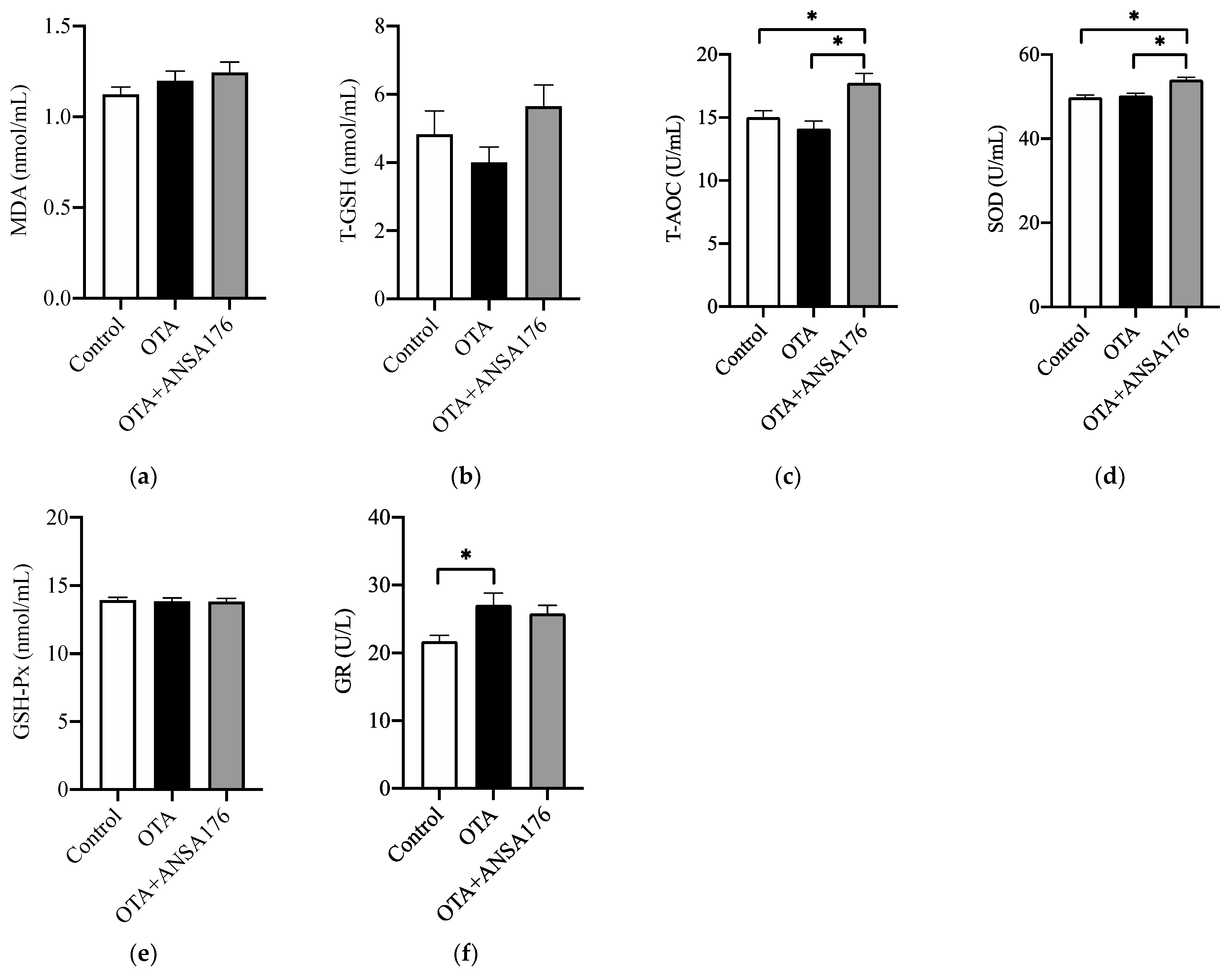
2.2.4. Oxidative Stress and Antioxidant Status in Laying Hens
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Isolation and Identification of the OTA-Degrading Strain
5.1.1. Culture Media and Reagents
5.1.2. Isolation of Microorganisms with OTA Degradation Ability
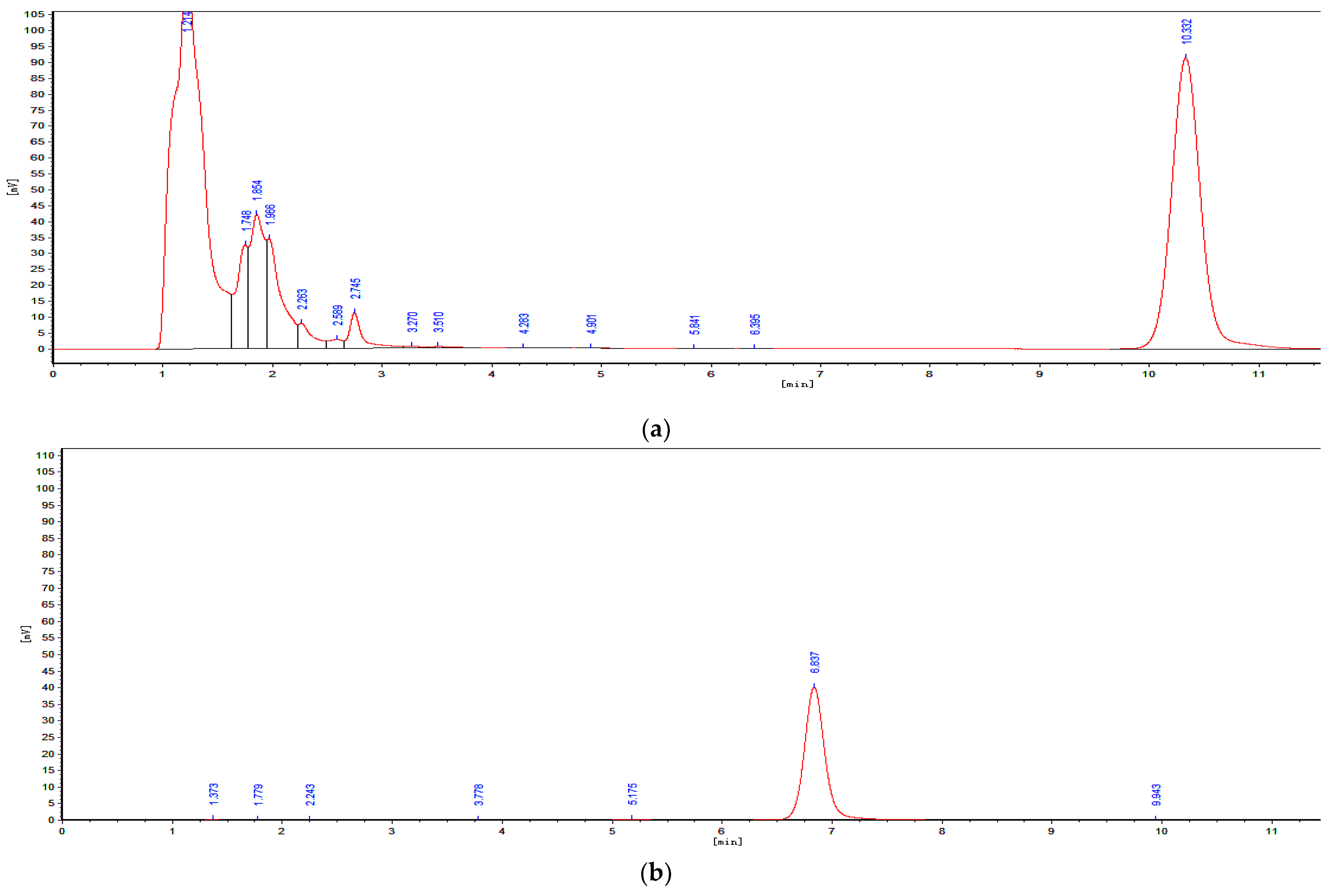
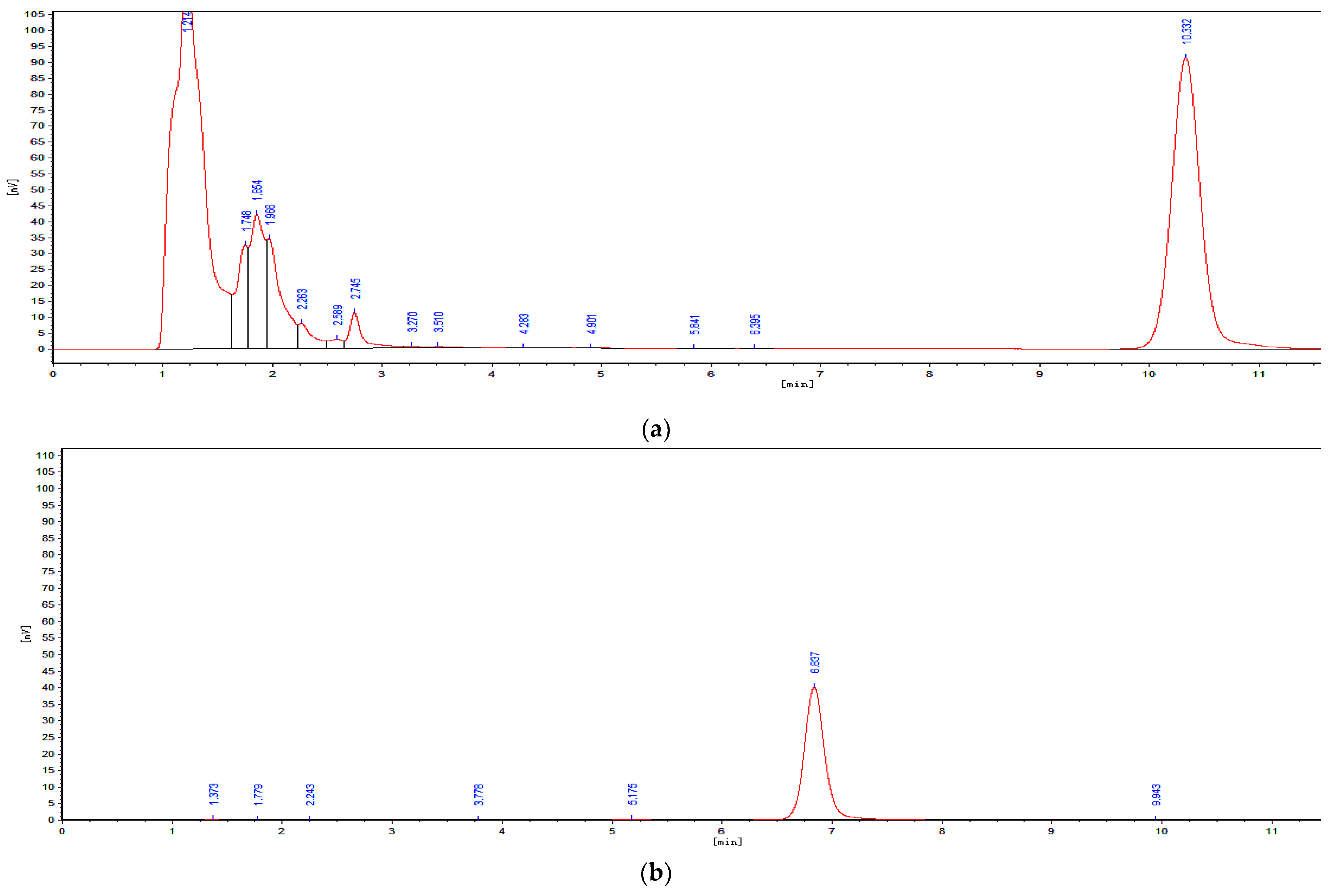
5.1.3. Determination of OTA and OTα by Using HPLC
5.1.4. Identification of the OTA-Degrading Strain
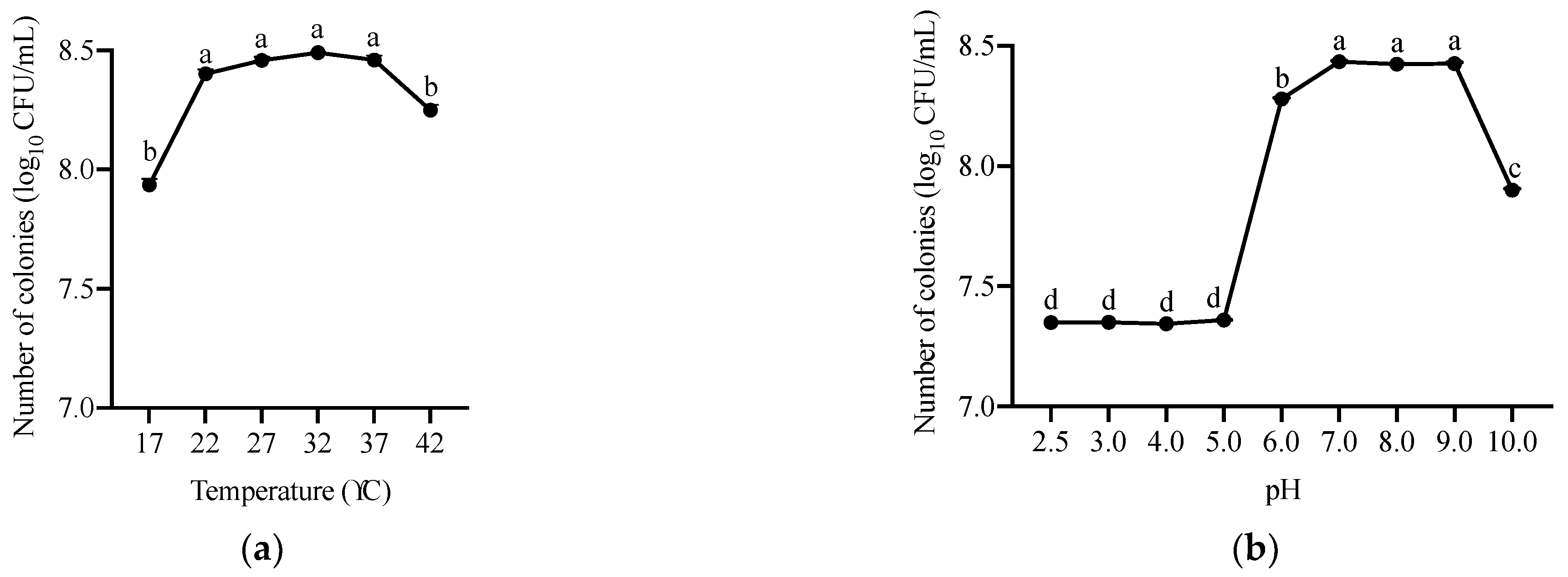
5.1.5. Characterization of A. faecalis ANSA176 Growth with Different Temperature and pH Levels
5.2. Animal Trial in Layer Hens
5.2.1. Dietary Treatments of Animal Trial
5.2.2. Design of Animal Experiments
5.2.3. Laying Performance and Egg Quality
5.2.4. Residues of OTA in Eggs
5.2.5. Blood Sampling and Serum Biochemical Analysis
5.2.6. Liver and Kidney Histological Assessment
5.3. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (%) | Basal Diet | Contaminated Diet |
|---|---|---|
| Normal corn | 65.05 | 47.05 |
| Contaminated corn 1 | 0 | 18 |
| Soybean meal (43% CP) | 24.20 | 24.20 |
| Limestone (38% Ca) | 8.20 | 8.20 |
| Calcium hydrophosphate (21% Ca, 16% P) | 1.70 | 1.70 |
| NaCl | 0.30 | 0.30 |
| DL-methionine | 0.12 | 0.12 |
| Mineral premix 2 | 0.30 | 0.30 |
| Vitamin premix 3 | 0.03 | 0.03 |
| Choline chloride | 0.10 | 0.10 |
| Total | 100.00 | 100.00 |
| Nutritional content (%) 4 | ||
| Crude protein | 16.04 | 16.04 |
| ME (MJ/kg) | 11.25 | 11.25 |
| Calcium | 3.60 | 3.60 |
| Total phosphorus | 0.65 | 0.65 |
| Non-phytin phosphorus | 0.39 | 0.39 |
| Methionine | 0.38 | 0.38 |
| Methionine + cystine | 0.65 | 0.65 |
| Lysine | 0.78 | 0.78 |
| Tryptophan | 0.16 | 0.16 |
| Threonine | 0.59 | 0.59 |
References
- Iqbal, S.Z.; Asi, M.R.; Hanif, U.; Zuber, M.; Jinap, S. The Presence of Aflatoxins and Ochratoxin A in Rice and Rice Products; and Evaluation of Dietary Intake. Food Chem. 2016, 210, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ma, W.; Ma, Z.; Zhang, Q.; Li, H. The Occurrence and Contamination Level of Ochratoxin A in Plant and Animal-Derived Food Commodities. Mol. Basel Switz. 2021, 26, 6928. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, C.; Zhang, B.; Zhou, Z.; Shen, Y.; Liao, X.; Yang, J.; Wang, Y.; Li, X.; Li, Y.; et al. Advances in Biodetoxification of Ochratoxin A-A Review of the Past Five Decades. Front. Microbiol. 2018, 9, 1386. [Google Scholar] [CrossRef] [PubMed]
- Pozzo, L.; Cavallarin, L.; Antoniazzi, S.; Guerre, P.; Biasibetti, E.; Capucchio, M.T.; Schiavone, A. Feeding a Diet Contaminated with Ochratoxin A for Broiler Chickens at the Maximum Level Recommended by the EU for Poultry Feeds (0.1 Mg/Kg). 2. Effects on Meat Quality, Oxidative Stress, Residues and Histological Traits. J. Anim. Physiol. Anim. Nutr. 2013, 97 (Suppl. 1), 23–31. [Google Scholar] [CrossRef]
- Tolosa, J.; Rodríguez-Carrasco, Y.; Ruiz, M.J.; Vila-Donat, P. Multi-Mycotoxin Occurrence in Feed, Metabolism and Carry-over to Animal-Derived Food Products: A Review. Food Chem. Toxicol. 2021, 158, 112661. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Some Traditional Herbal Medicines, Some Mycotoxins, Naphthalene and Styrene. IARC Monogr. Eval. Carcinog. Risks Hum. 2002, 82, 1–556. [Google Scholar]
- Pleadin, J. Mycotoxins in Grains and Feed: Contamination and Toxic Effect in Animals. Biotechnol. Anim. Husb. 2015, 31, 441–456. [Google Scholar] [CrossRef]
- Duarte, S.C.; Lino, C.M.; Pena, A. Food Safety Implications of Ochratoxin A in Animal-Derived Food Products. Vet. J. 2012, 192, 286–292. [Google Scholar] [CrossRef]
- European Commission. Commission Recommendation of 17 August 2006 on the Presence of Deoxynivalenol, Zearalenone, Ochratoxin A, T-2 and HT-2 and Fumonisins in Products Intended for Animal Feeding. Off. J. Eur. Union 2006, 229, 7–9. [Google Scholar]
- Hassan, Z.U.; Khan, M.Z.; Khan, A.; Javed, I.; Hussain, Z. Effects of Individual and Combined Administration of Ochratoxin A and Aflatoxin B1 in Tissues and Eggs of White Leghorn Breeder Hens. J. Sci. Food Agric. 2012, 92, 1540–1544. [Google Scholar] [CrossRef]
- Denli, M.; Blandon, J.C.; Guynot, M.E.; Salado, S.; Perez, J.F. Efficacy of a New Ochratoxin-Binding Agent (OcraTox) to Counteract the Deleterious Effects of Ochratoxin A in Laying Hens. Poult. Sci. 2008, 87, 2266–2272. [Google Scholar] [CrossRef]
- Denli, M.; Perez, J.F. Ochratoxins in Feed, a Risk for Animal and Human Health: Control Strategies. Toxins 2010, 2, 1065–1077. [Google Scholar] [CrossRef]
- Vasiljević, M.; Marinković, D.; Milićević, D.; Pleadin, J.; Stefanović, S.; Trialović, S.; Raj, J.; Petrujkić, B.; Trialović, J.N. Efficacy of a Modified Clinoptilolite Based Adsorbent in Reducing Detrimental Effects of Ochratoxin A in Laying Hens. Toxins 2021, 13, 469. [Google Scholar] [CrossRef]
- Tao, Y.; Xie, S.; Xu, F.; Liu, A.; Wang, Y.; Chen, D.; Pan, Y.; Huang, L.; Peng, D.; Wang, X.; et al. Ochratoxin A: Toxicity, Oxidative Stress and Metabolism. Food Chem. Toxicol. 2018, 112, 320–331. [Google Scholar] [CrossRef]
- Hameed, M.R.; Khan, M.Z.; Saleemi, M.K.; Khan, A.; Akhtar, M.; Hassan, Z.; Hussain, Z. Study of Ochratoxin A (OTA)-Induced Oxidative Stress Markers in Broiler Chicks. Toxin Rev. 2017, 36, 270–274. [Google Scholar] [CrossRef]
- Samuel, M.S.; Jeyaram, K.; Datta, S.; Chandrasekar, N.; Balaji, R.; Selvarajan, E. Detection, Contamination, Toxicity, and Prevention Methods of Ochratoxins: An Update Review. J. Agric. Food Chem. 2021, 69, 13974–13989. [Google Scholar] [CrossRef]
- Var, I.; Erginkaya, Z.; Kabak, B. Reduction of Ochratoxin A Levels in White Wine by Yeast Treatments. J. Inst. Brew. 2009, 115, 30–34. [Google Scholar] [CrossRef]
- Piotrowska, M.; Zakowska, Z. The Elimination of Ochratoxin A by Lactic Acid Bacteria Strains. Pol. J. Microbiol. 2005, 54, 279–286. [Google Scholar]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Biodegradation of Ochratoxin A by Aspergillus Section Nigri Species Isolated from French Grapes: A Potential Means of Ochratoxin A Decontamination in Grape Juices and Musts. FEMS Microbiol. Lett. 2006, 255, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Varga, J.; Péteri, Z.; Tábori, K.; Téren, J.; Vágvölgyi, C. Degradation of Ochratoxin A and Other Mycotoxins by Rhizopus Isolates. Int. J. Food Microbiol. 2005, 99, 321–328. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Serra, R.; Venâncio, A. Biodegradation of Ochratoxin A by Fungi Isolated from Grapes. J. Agric. Food Chem. 2002, 50, 7493–7496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luz, C.; Ferrer, J.; Mañes, J.; Meca, G. Toxicity Reduction of Ochratoxin A by Lactic Acid Bacteria. Food Chem. Toxicol. 2018, 112, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Wang, Y.; Zhao, C.; Wang, J.; Zhang, X.L. Biodegradation of Ochratoxin A by Alcaligenes Faecalis Isolated from Soil. J. Appl. Microbiol. 2017, 123, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Angioni, A.; Caboni, P.; Garau, A.; Farris, A.; Orro, D.; Budroni, M.; Cabras, P. In Vitro Interaction between Ochratoxin A and Different Strains of Saccharomyces Cerevisiae and Kloeckera Apiculata. J. Agric. Food Chem. 2007, 55, 2043–2048. [Google Scholar] [CrossRef]
- Xiong, K.; Wang, X.; Zhi, H.-W.; Sun, B.-G.; Li, X.-T. Identification and Safety Evaluation of a Product from the Biodegradation of Ochratoxin A by an Aspergillus Strain. J. Sci. Food Agric. 2017, 97, 434–443. [Google Scholar] [CrossRef]
- Bellis, P.; Tristezza, M.; Haidukowski, M.; Fanelli, F.; Sisto, A.; Mule, G.; Greco, F. Biodegradation of Ochratoxin A by Bacterial Strains Isolated from Vineyard Soils. Toxins 2015, 7, 5079–5093. [Google Scholar] [CrossRef]
- Loi, M.; Fanelli, F.; Liuzzi, V.C.; Logrieco, A.F.; Mulè, G. Mycotoxin Biotransformation by Native and Commercial Enzymes: Present and Future Perspectives. Toxins 2017, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, Y.; Yin, T.; Wang, J.; Zhang, X. Heterologous Expression and Characterization of A Novel Ochratoxin A Degrading Enzyme, N-Acyl-L-Amino Acid Amidohydrolase, from Alcaligenes Faecalis. Toxins 2019, 11, 518. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.; Wu, Z.; Wu, S.; Dai, Y.; Sun, C. Degradation of Ochratoxin A by Bacillus Amyloliquefaciens ASAG1. Food Addit. Contam. Part. Chem. Anal. Control. Expo. Risk Assess. 2015, 32, 564–571. [Google Scholar] [CrossRef]
- Liuzzi, V.C.; Fanelli, F.; Tristezza, M.; Haidukowski, M.; Picardi, E.; Manzari, C.; Lionetti, C.; Grieco, F.; Logrieco, A.F.; Thon, M.R.; et al. Transcriptional Analysis of Acinetobacter Sp. Neg1 Capable of Degrading Ochratoxin A. Front. Microbiol. 2017, 7, 2162. [Google Scholar] [CrossRef] [Green Version]
- Qing, H.; Huo, X.; Huang, S.; Zhao, L.; Zhang, J.; Ji, C.; Ma, Q. Bacillus subtilis ANSB168 Producing d-Alanyl-d-Alanine Carboxypeptidase Could Alleviate the Immune Injury and Inflammation Induced by Ochratoxin A. Int. J. Mol. Sci. 2021, 22, 12059. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Santos, L.; Venâncio, A. Degradation of Ochratoxin A by Proteases and by a Crude Enzyme of Aspergillus Niger. Food Biotechnol. 2006, 20, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Dobritzsch, D.; Wang, H.; Schneider, G.; Yu, S. Structural and Functional Characterization of Ochratoxinase, a Novel Mycotoxin-Degrading Enzyme. Biochem. J. 2014, 462, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Stander, M.A.; Bornscheuer, U.T.; Henke, E.; Steyn, P.S. Screening of Commercial Hydrolases for the Degradation of Ochratoxin A. J. Agric. Food Chem. 2000, 48, 5736–5739. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Venâncio, A.; Teixeira, J.A. Optimization of Process Parameters for the Production of an OTA-Hydrolyzing Enzyme from Aspergillus Niger under Solid-State Fermentation. J. Biosci. Bioeng. 2011, 112, 351–355. [Google Scholar] [CrossRef]
- Johri, T.S.; Swain, B. Effect of Varying Levels of Aflatoxin, Ochratoxin and Their Combinations on the Performance and Egg Quality Characteristics in Laying Hens. Asian-Australas. J. Anim. Sci. 2003, 16, 1015–1019. [Google Scholar] [CrossRef]
- Jovanović, M.; Zlatanović, S.; Micić, D.; Bacić, D.; Mitić-Ćulafić, D.; Đuriš, M.; Gorjanović, S. Functionality and Palatability of Yogurt Produced Using Beetroot Pomace Flour Granulated with Lactic Acid Bacteria. Foods Basel Switz. 2021, 10, 1696. [Google Scholar] [CrossRef]
- Shiferaw Terefe, N.; Augustin, M.A. Fermentation for Tailoring the Technological and Health Related Functionality of Food Products. Crit. Rev. Food Sci. Nutr. 2020, 60, 2887–2913. [Google Scholar] [CrossRef]
- Qing, H.; Huang, S.; Zhan, K.; Zhao, L.; Zhang, J.; Ji, C.; Ma, Q. Combined Toxicity Evaluation of Ochratoxin A and Aflatoxin B1 on Kidney and Liver Injury, Immune Inflammation, and Gut Microbiota Alteration Through Pair-Feeding Pullet Model. Front. Immunol. 2022, 13, 3539. [Google Scholar] [CrossRef]
- Emeigh Hart, S.G. 7.11–In Vivo Methodologies Used to Assess Renal Function and Injury. In Comprehensive Toxicology, 2nd ed.; McQueen, C.A., Ed.; Elsevier: Oxford, UK, 2010; pp. 263–303. ISBN 978-0-08-046884-6. [Google Scholar]
- Yordanova, P.; Wilfried, K.; Tsolova, S.; Dimitrov, P. Ochratoxin A and Β2-Microglobulin in BEN Patients and Controls. Toxins 2010, 2, 780–792. [Google Scholar] [CrossRef] [Green Version]
- Marin, D.; Taranu, I. Ochratoxin A and Its Effects on Immunity. Toxin Rev. 2015, 34, 11–20. [Google Scholar] [CrossRef]
- Darif, Y.; Mountassif, D.; Belkebir, A.; Zaid, Y.; Basu, K.; Mourad, W.; Oudghiri, M. Ochratoxin A Mediates MAPK Activation, Modulates IL-2 and TNF-α MRNA Expression and Induces Apoptosis by Mitochondria-Dependent and Mitochondria-Independent Pathways in Human H9 T Cells. J. Toxicol. Sci. 2016, 41, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Al-Anati, L.; Essid, E.; Stenius, U.; Beuerlein, K.; Schuh, K.; Petzinger, E. Differential Cell Sensitivity between OTA and LPS upon Releasing TNF-α. Toxins 2010, 2, 1279–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kőszegi, T.; Poór, M. Ochratoxin A: Molecular Interactions, Mechanisms of Toxicity and Prevention at the Molecular Level. Toxins 2016, 8, 111. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahhab, M.A.; Aljawish, A.; El-Nekeety, A.A.; Abdel-Aziem, S.H.; Hassan, N.S. Chitosan Nanoparticles plus Quercetin Suppress the Oxidative Stress, Modulate DNA Fragmentation and Gene Expression in the Kidney of Rats Fed Ochratoxin A-Contaminated Diet. Food Chem. Toxicol. 2017, 99, 209–221. [Google Scholar] [CrossRef] [PubMed]
- El-Shafie, M.; Elmasry, H.G.; Elsadek, M.F.B.; AlMajwal, A.M. Curative Effect of Orally Consumed Aloe Vera Juice on Ochratoxin A-Induced Nephrotoxicity in Rats. Prog. Nutr. 2015, 17, 128–136. [Google Scholar]
- Yang, Q.; Shi, L.; Huang, K.; Xu, W. Protective Effect of N-Acetylcysteine against DNA Damage and S-Phase Arrest Induced by Ochratoxin A in Human Embryonic Kidney Cells (HEK-293). Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2014, 70, 40–47. [Google Scholar] [CrossRef]
- Baudrimont, I.; Betbeder, A.M.; Gharbi, A.; Pfohl-Leszkowicz, A.; Dirheimer, G.; Creppy, E.E. Effect of Superoxide Dismutase and Catalase on the Nephrotoxicity Induced by Subchronical Administration of Ochratoxin A in Rats. Toxicology 1994, 89, 101–111. [Google Scholar] [CrossRef]
- Marin-Kuan, M.; Nestler, S.; Verguet, C.; Bezençon, C.; Piguet, D.; Mansourian, R.; Holzwarth, J.; Grigorov, M.; Delatour, T.; Mantle, P.; et al. A Toxicogenomics Approach to Identify New Plausible Epigenetic Mechanisms of Ochratoxin a Carcinogenicity in Rat. Toxicol. Sci. Off. J. Soc. Toxicol. 2006, 89, 120–134. [Google Scholar] [CrossRef]
- Geng, S.; Huang, S.; Ma, Q.; Li, F.; Gao, Y.; Zhao, L.; Zhang, J. Alterations and Correlations of the Gut Microbiome, Performance, Egg Quality, and Serum Biochemical Indexes in Laying Hens with Low-Protein Amino Acid-Deficient Diets. ACS Omega 2021, 6, 13094–13104. [Google Scholar] [CrossRef]
- Fan, Y.; Zhao, L.; Ji, C.; Li, X.; Jia, R.; Xi, L.; Zhang, J.; Ma, Q. Protective Effects of Bacillus Subtilis ANSB060 on Serum Biochemistry, Histopathological Changes and Antioxidant Enzyme Activities of Broilers Fed Moldy Peanut Meal Naturally Contaminated with Aflatoxins. Toxins 2015, 7, 3330–3343. [Google Scholar] [CrossRef]



| Substrate/Test | Result | Substrate/Test | Result | Substrate/Test | Result | Substrate/Test | Result |
|---|---|---|---|---|---|---|---|
| Cell shape | Rod | Gram Staining | − | Catalase | + | Oxidase | + |
| BIOLOG GEN III (carbon source utilization assays) | |||||||
| Negative control | − | α-D-glucose | − | Gelatin | − | D-raffinose | − |
| 3-methyl-D-glucose | − | D-mannose | − | Gentiobiose | − | α-D-lactose | − |
| D-glucose-6-PO4 | − | D-fructose | − | L-alanine | + | D-melibiose | − |
| Glucuronamide | + | D-galactose | − | L-arginine | − | L-lactic acid | + |
| D-cellobiose | − | Dextrin | − | L-glutamic acid | + | Citric acid | + |
| Glycyl-L-proline | − | L-rhamnose | − | L-histidine | + | α-ketoglutaric acid | + |
| L-aspartic acid | + | Inosine | − | L-pyroglutamic acid | + | D-malic acid | − |
| p-hydroxyphenyl acetic acid | + | D-sorbitol | − | L-serine | − | L-malic acid | + |
| Methyl pyruvate | + | D-mannitol | − | Pectin | − | Bromo succinic acid | + |
| D-lactic acid methyl ester | − | D-arabitol | − | Sucrose | − | γ-aminobutyric acid | − |
| β-methyl-D-glucoside | − | Myo-inositol | − | Turanose | − | α-hydroxybutyric acid | + |
| β-hydroxy-D,L-butyric acid | + | Glycerol | − | D-gluconic acid | − | D-salicin | − |
| N-acetyl-D-glucosamine | − | D-maltose | − | Stachyose | − | α-keto-butyric acid | + |
| N-acetyl-β-D-mannosamine | − | D-trehalose | − | Mucic acid | − | Propionic | + |
| N-acetyl-D-galactosamine | − | D-aspartic acid | − | Quinic acid | − | Acetic acid | + |
| N-neuraminic acid | − | D-serine | + | Saccharic acid | − | Formic acid | + |
| BIOLOG GEN III (chemical sensitivity assays; +, not sensitive; −, sensitive) | |||||||
| Positive control | + | Tetrazolium blue | + | pH 6.0 | + | Nalidixic acid | − |
| Na bromate | − | Fusidic acid | + | pH 5.0 | + | LiCl | + |
| Rifamycin SV | + | D-serine | + | 1% Na-lactase | + | K-tellurite | − |
| Minocycline | + | Lincomycin | + | 1% NaCl | + | Aztreonam | + |
| Vancomycin | + | Guanidine HCl | + | 4% NaCl | + | Na-butyrate | + |
| Niaproof 4 | + | Tetrazolium violet | + | 8% NaCl | + | ||
| Treatment | Control | OTA | OTA + ANSA176 | SEM 1 | p-Value |
|---|---|---|---|---|---|
| Egg weight (g) | 54.95 a | 46.64 b | 55.08 a | 1.20 | <0.01 |
| Shell percentage (%) | 9.92 b | 12.17 a | 10.23 b | 0.30 | <0.01 |
| Yolk percentage (%) | 25.90 b | 31.81 a | 25.35 b | 1.07 | <0.01 |
| Albumen percentage (%) | 64.18 a | 56.01 b | 64.43 a | 1.31 | <0.01 |
| Shell color | 58.03 | 58.01 | 57.69 | 1.31 | 0.98 |
| Shell thickness (mm) | 0.41 | 0.41 | 0.41 | 0.00 | 0.37 |
| Shell strength (N) | 40.75 | 38.77 | 39.90 | 1.12 | 0.46 |
| Haugh unit | 83.87 | 81.14 | 82.93 | 1.37 | 0.36 |
| Yolk color | 5.37 | 5.33 | 4.93 | 0.16 | 0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, R.; Qing, H.; Ma, Q.; Huo, X.; Huang, S.; Zhao, L.; Zhang, J.; Ji, C. A Newly Isolated Alcaligenes faecalis ANSA176 with the Capability of Alleviating Immune Injury and Inflammation through Efficiently Degrading Ochratoxin A. Toxins 2022, 14, 569. https://doi.org/10.3390/toxins14080569
Zheng R, Qing H, Ma Q, Huo X, Huang S, Zhao L, Zhang J, Ji C. A Newly Isolated Alcaligenes faecalis ANSA176 with the Capability of Alleviating Immune Injury and Inflammation through Efficiently Degrading Ochratoxin A. Toxins. 2022; 14(8):569. https://doi.org/10.3390/toxins14080569
Chicago/Turabian StyleZheng, Rui, Hanrui Qing, Qiugang Ma, Xueting Huo, Shimeng Huang, Lihong Zhao, Jianyun Zhang, and Cheng Ji. 2022. "A Newly Isolated Alcaligenes faecalis ANSA176 with the Capability of Alleviating Immune Injury and Inflammation through Efficiently Degrading Ochratoxin A" Toxins 14, no. 8: 569. https://doi.org/10.3390/toxins14080569
APA StyleZheng, R., Qing, H., Ma, Q., Huo, X., Huang, S., Zhao, L., Zhang, J., & Ji, C. (2022). A Newly Isolated Alcaligenes faecalis ANSA176 with the Capability of Alleviating Immune Injury and Inflammation through Efficiently Degrading Ochratoxin A. Toxins, 14(8), 569. https://doi.org/10.3390/toxins14080569