Development of a Monoclonal scFv against Cytotoxin to Neutralize Cytolytic Activity Induced by Naja atra Venom on Myoblast C2C12 Cells

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
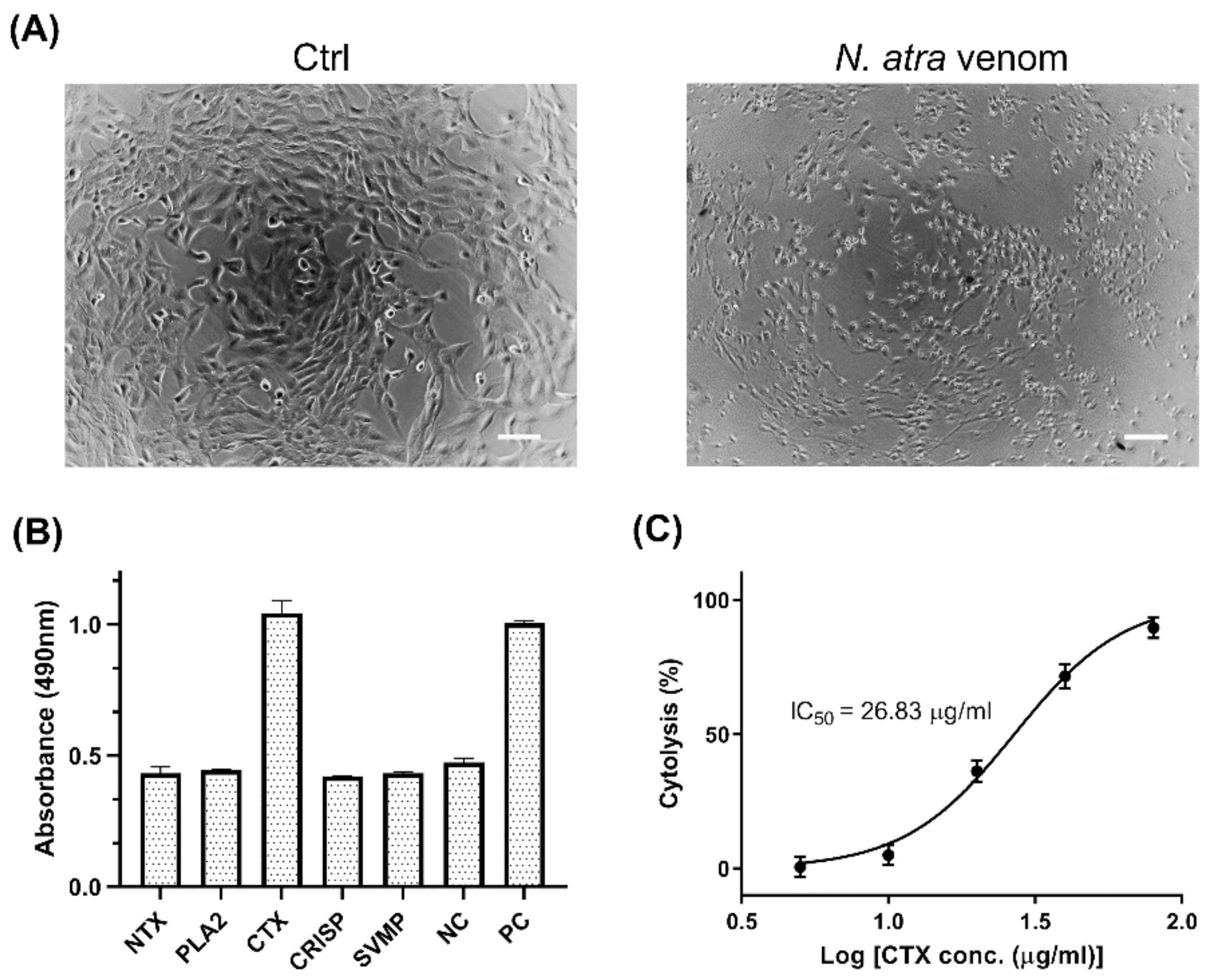
2.1. Cytotoxicity of N. atra Venom Proteins in C2C12 Cells
2.2. Production and Selection of Anti-CTX Antibodies
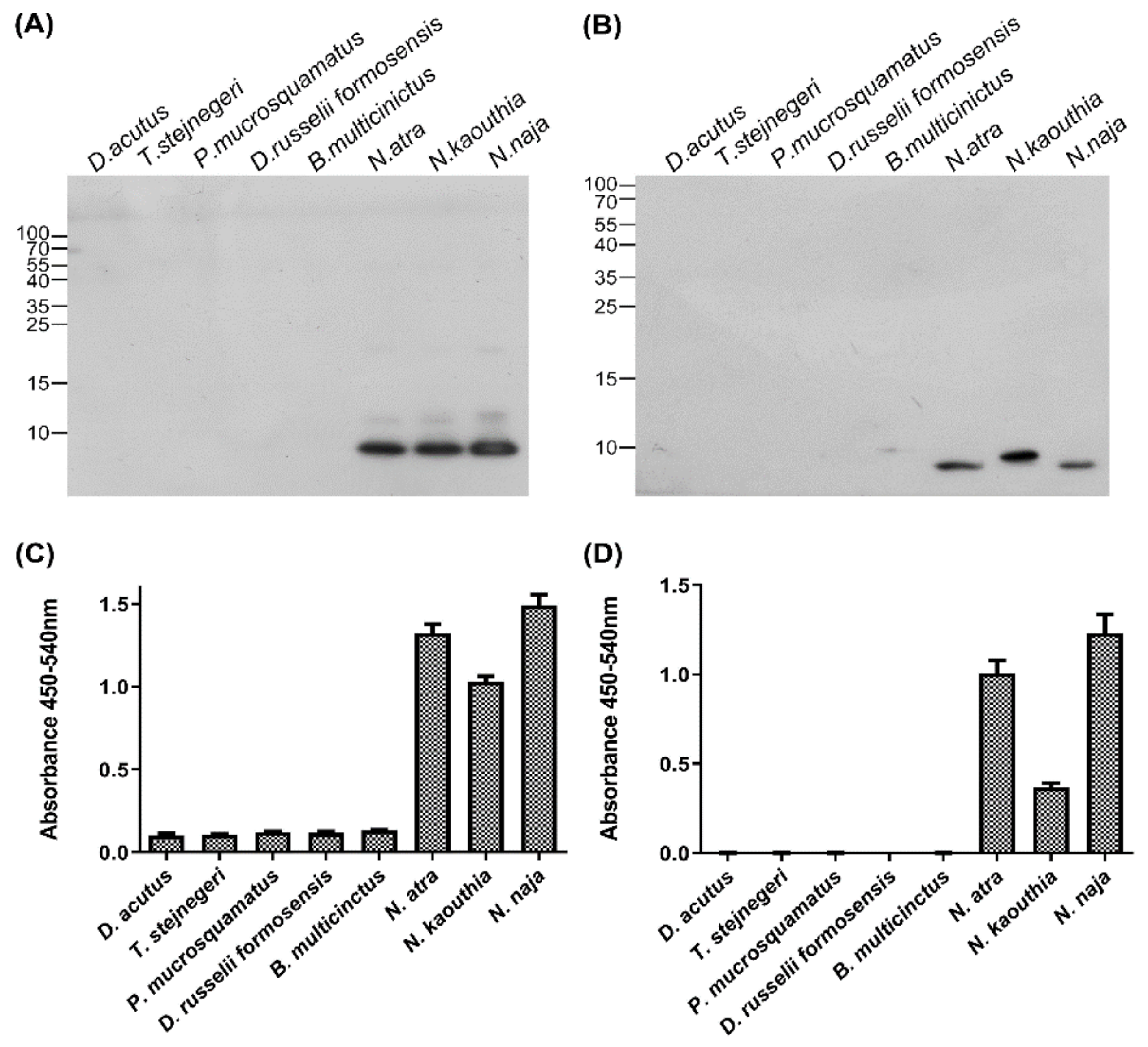
2.3. Specificity of Anti-CTX Antibodies
2.4. Abilities of Anti-CTX Antibodies and Conventional Antivenom to Neutralize CTX-Induced Cytotoxicity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Snake Venoms and Anivenom
5.2. Cytotoxicity Assay
5.3. Antigen Preparation and Hen Immunization
5.4. Library Construction and Biopanning of Anti-CTX scFv
5.5. Expression and Purification of Anti-CTX scFv
5.6. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Western Blotting
5.7. Indirect Enzyme-Linked Immunosorbent Assay (ELISA)
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chippaux, J.P. Snake-bites appraisal of the global situation. Bull. World Health Organ. 1998, 76, 515–524. [Google Scholar] [PubMed]
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [Green Version]
- Cruz, L.S.; Vargas, R.; Lopes, A.n.A. Snakebite envenomation and death in the developing world. Ethn. Dis. 2009, 19, 42–46. [Google Scholar]
- Gutierrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Primers 2017, 3, 17063. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Williams, D.; Fan, H.W.; Warrell, D.A. Snakebite envenoming from a global perspective: Towards an integrated approach. Toxicon 2010, 56, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.R.; Sashindran, V.K. Clinical Features And Management Of Snake Bite. Med. J. Armed Forces India 2002, 58, 247–249. [Google Scholar] [CrossRef] [Green Version]
- Gasanov, S. Snake Venom Cytotoxins, Phospholipase A2s, and Zn2+-dependent Metalloproteinases Mechanisms of Action and Pharmacological Relevance. J. Clin. Toxicol. 2014, 4, 1000181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, J.M.; Theakston, R.D.; Warrell, D.A. Confronting the neglected problem of snake bite envenoming: The need for a global partnership. PLoS Med. 2006, 3, e150. [Google Scholar] [CrossRef] [Green Version]
- HA, R.; Theakston, R.D. The management of snake bite. Bull. World Health Organ. 1983, 61, 885–895. [Google Scholar]
- Mao, Y.C.; Liu, P.Y.; Chiang, L.C.; Lai, C.S.; Lai, K.L.; Ho, C.H.; Wang, T.H.; Yang, C.C. Naja atra snakebite in Taiwan. Clin. Toxicol. 2018, 56, 273–280. [Google Scholar] [CrossRef]
- Hung, D.-Z. Taiwan’s venomous snakebite: Epidemiological, evolution and geographic differences. Trans. R. Soc. Trop. Med. Hyg. 2004, 98, 96–101. [Google Scholar] [CrossRef]
- Sanz, L.; Quesada-Bernat, S.; Chen, P.Y.; Lee, C.D.; Chiang, J.R.; Calvete, J.J. Translational Venomics: Third-Generation Antivenomics of Anti-Siamese Russell’s Viper, Daboia siamensis, Antivenom Manufactured in Taiwan CDC’s Vaccine Center. Trop. Med. Infect. Dis. 2018, 3, 66. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.C.; Chaou, C.H.; Tseng, C.Y. An investigation of snakebite antivenom usage in Taiwan. J. Formos. Med. Assoc. 2016, 115, 672–677. [Google Scholar] [CrossRef] [Green Version]
- Standker, L.; Harvey, A.L.; Furst, S.; Mathes, I.; Forssmann, W.G.; Escalona de Motta, G.; Beress, L. Improved method for the isolation, characterization and examination of neuromuscular and toxic properties of selected polypeptide fractions from the crude venom of the Taiwan cobra Naja naja atra. Toxicon 2012, 60, 623–631. [Google Scholar] [CrossRef]
- Mao, Y.C.; Liu, P.Y.; Hung, D.Z.; Lai, W.C.; Huang, S.T.; Hung, Y.M.; Yang, C.C. Bacteriology of Naja atra Snakebite Wound and Its Implications for Antibiotic Therapy. Am. J. Trop. Med. Hyg. 2016, 94, 1129–1135. [Google Scholar] [CrossRef]
- Liu, C.C.; Chou, Y.S.; Chen, C.Y.; Liu, K.L.; Huang, G.J.; Yu, J.S.; Wu, C.J.; Liaw, G.W.; Hsieh, C.H.; Chen, C.K. Pathogenesis of local necrosis induced by Naja atra venom: Assessment of the neutralization ability of Taiwanese freeze-dried neurotoxic antivenom in animal models. PLoS Negl. Trop. Dis. 2020, 14, e0008054. [Google Scholar] [CrossRef] [Green Version]
- Hiu, J.J.; Yap, M.K.K. The myth of cobra venom cytotoxin: More than just direct cytolytic actions. Toxicon X 2022, 14, 100123. [Google Scholar] [CrossRef]
- Lipman, N.S.; Jackson, L.R.; Trudel, L.J.; Weis-Garcia, F. Monoclonal versus polyclonal antibodies: Distinguishing characteristics, applications, and information resources. ILAR J. 2005, 46, 258–268. [Google Scholar] [CrossRef] [Green Version]
- Carter, P.J. Potent antibody therapeutics by design. Nat. Rev. Immunol. 2006, 6, 343–357. [Google Scholar] [CrossRef]
- Reichert, J.M.; Valge-Archer, V.E. Development trends for monoclonal antibody cancer therapeutics. Nat. Rev. Drug Discov. 2007, 6, 349–356. [Google Scholar] [CrossRef]
- Kulkeaw, K.; Sakolvaree, Y.; Srimanote, P.; Tongtawe, P.; Maneewatch, S.; Sookrung, N.; Tungtrongchitr, A.; Tapchaisri, P.; Kurazono, H.; Chaicumpa, W. Human monoclonal ScFv neutralize lethal Thai cobra, Naja kaouthia, neurotoxin. J. Proteom. 2009, 72, 270–282. [Google Scholar] [CrossRef] [PubMed]
- Tomita, M.; Tsumoto, K. Hybridoma technologies for antibody production. Immunotherapy 2011, 3, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Hammers, C.M.; Stanley, J.R. Antibody phage display: Technique and applications. J. Investig. Dermatol. 2014, 134, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frenzel, A.; Schirrmann, T.; Hust, M. Phage display-derived human antibodies in clinical development and therapy. MAbs 2016, 8, 1177–1194. [Google Scholar] [CrossRef] [Green Version]
- Laustsen, A.H.; Maria Gutierrez, J.; Knudsen, C.; Johansen, K.H.; Bermudez-Mendez, E.; Cerni, F.A.; Jurgensen, J.A.; Ledsgaard, L.; Martos-Esteban, A.; Ohlenschlaeger, M.; et al. Pros and cons of different therapeutic antibody formats for recombinant antivenom development. Toxicon 2018, 146, 151–175. [Google Scholar] [CrossRef]
- Negrin, A.; Battocletti, M.A.; Juanena, C.; Morais, V. Reports of Doses Administered and Adverse Reactions to Snake Antivenom Used in Uruguay in 2018. Front. Toxicol. 2021, 3, 690964. [Google Scholar] [CrossRef]
- De Silva, H.A.; Ryan, N.M.; de Silva, H.J. Adverse reactions to snake antivenom, and their prevention and treatment. Br. J. Clin. Pharmacol. 2016, 81, 446–452. [Google Scholar] [CrossRef] [Green Version]
- Gawarammana, I.; Keyler, D. Dealing with adverse reactions to snake antivenom. Ceylon Med. J. 2011, 56, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.J.; Jensen, S.D.; Nimorakiotakis, B.; Muller, R.; Winkel, K.D. Antivenom use, premedication and early adverse reactions in the management of snake bites in rural Papua New Guinea. Toxicon 2007, 49, 780–792. [Google Scholar] [CrossRef]
- Chakrabartty, S.; Alam, M.I.; Bhagat, S.; Alam, A.; Dhyani, N.; Khan, G.A.; Alam, M.S. Inhibition of snake venom induced sterile inflammation and PLA2 activity by Titanium dioxide Nanoparticles in experimental animals. Sci. Rep. 2019, 9, 11175. [Google Scholar] [CrossRef] [Green Version]
- Albulescu, L.O.; Xie, C.; Ainsworth, S.; Alsolaiss, J.; Crittenden, E.; Dawson, C.A.; Softley, R.; Bartlett, K.E.; Harrison, R.A.; Kool, J.; et al. A therapeutic combination of two small molecule toxin inhibitors provides broad preclinical efficacy against viper snakebite. Nat. Commun. 2020, 11, 6094. [Google Scholar] [CrossRef]
- Wei, S.; Duan, S.; Liu, X.; Wang, H.; Ding, S.; Chen, Y.; Xie, J.; Tian, J.; Yu, N.; Ge, P.; et al. Chicken Egg Yolk Antibodies (IgYs) block the binding of multiple SARS-CoV-2 spike protein variants to human ACE2. Int. Immunopharmacol. 2021, 90, 107172. [Google Scholar] [CrossRef]
- Lee, C.H.; Liu, C.I.; Leu, S.J.; Lee, Y.C.; Chiang, J.R.; Chiang, L.C.; Mao, Y.C.; Tsai, B.Y.; Hung, C.S.; Chen, C.C.; et al. Chicken antibodies against venom proteins of Trimeresurus stejnegeri in Taiwan. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200056. [Google Scholar] [CrossRef]
- Lee, C.H.; Lee, Y.C.; Leu, S.J.; Lin, L.T.; Chiang, J.R.; Hsu, W.J.; Yang, Y.Y. Production and Characterization of Neutralizing Antibodies against Bungarus multicinctus Snake Venom. Appl. Environ. Microbiol. 2016, 82, 6973–6982. [Google Scholar] [CrossRef] [Green Version]
- Laustsen, A.H.; Karatt-Vellatt, A.; Masters, E.W.; Arias, A.S.; Pus, U.; Knudsen, C.; Oscoz, S.; Slavny, P.; Griffiths, D.T.; Luther, A.M.; et al. In vivo neutralization of dendrotoxin-mediated neurotoxicity of black mamba venom by oligoclonal human IgG antibodies. Nat. Commun. 2018, 9, 3928. [Google Scholar] [CrossRef]
- Maynard, J.; Georgiou, G. Antibody engineering. Annu. Rev. Biomed. Eng. 2000, 2, 339–376. [Google Scholar] [CrossRef]
- Watkins, N.A.; Ouwehand, W.H. Introduction to antibody engineering and phage display. Vox Sang. 2000, 78, 72–79. [Google Scholar] [CrossRef]
- Andris-Widhopf, J.; Rader, C.; Steinberger, P.; Fuller, R.; Barbas, C.F., 3rd. Methods for the generation of chicken monoclonal antibody fragments by phage display. J. Immunol. Methods 2000, 242, 159–181. [Google Scholar] [CrossRef]
- Gutierrez, J.M. Understanding and confronting snakebite envenoming: The harvest of cooperation. Toxicon 2016, 109, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, J.M.; Solano, G.; Pla, D.; Herrera, M.; Segura, A.; Vargas, M.; Villalta, M.; Sanchez, A.; Sanz, L.; Lomonte, B.; et al. Preclinical Evaluation of the Efficacy of Antivenoms for Snakebite Envenoming: State-of-the-Art and Challenges Ahead. Toxins 2017, 9, 163. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Chanda, A.; Kalita, B.; Islam, T.; Patra, A.; Mukherjee, A.K. Proteomic analysis to unravel the complex venom proteome of eastern India Naja naja: Correlation of venom composition with its biochemical and pharmacological properties. J. Proteom. 2017, 156, 29–39. [Google Scholar] [CrossRef]
- Adamude, F.A.; Dingwoke, E.J.; Abubakar, M.S.; Ibrahim, S.; Mohamed, G.; Klein, A.; Sallau, A.B. Proteomic analysis of three medically important Nigerian Naja (Naja haje, Naja katiensis and Naja nigricollis) snake venoms. Toxicon 2021, 197, 24–32. [Google Scholar] [CrossRef]
- Chanda, A.; Patra, A.; Kalita, B.; Mukherjee, A.K. Proteomics analysis to compare the venom composition between Naja naja and Naja kaouthia from the same geographical location of eastern India: Correlation with pathophysiology of envenomation and immunological cross-reactivity towards commercial polyantivenom. Expert Rev. Proteom. 2018, 15, 949–961. [Google Scholar] [CrossRef]
- Laustsen, A.H.; Gutierrez, J.M.; Lohse, B.; Rasmussen, A.R.; Fernandez, J.; Milbo, C.; Lomonte, B. Snake venomics of monocled cobra (Naja kaouthia) and investigation of human IgG response against venom toxins. Toxicon 2015, 99, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauridsen, L.P.; Laustsen, A.H.; Lomonte, B.; Gutierrez, J.M. Exploring the venom of the forest cobra snake: Toxicovenomics and antivenom profiling of Naja melanoleuca. J. Proteom. 2017, 150, 98–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petras, D.; Sanz, L.; Segura, A.; Herrera, M.; Villalta, M.; Solano, D.; Vargas, M.; Leon, G.; Warrell, D.A.; Theakston, R.D.; et al. Snake venomics of African spitting cobras: Toxin composition and assessment of congeneric cross-reactivity of the pan-African EchiTAb-Plus-ICP antivenom by antivenomics and neutralization approaches. J. Proteome Res. 2011, 10, 1266–1280. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Rojas, E.; Quesada, L.; Leon, G.; Nunez, J.; Laing, G.D.; Sasa, M.; Renjifo, J.M.; Nasidi, A.; Warrell, D.A.; et al. Pan-African polyspecific antivenom produced by caprylic acid purification of horse IgG: An alternative to the antivenom crisis in Africa. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 468–475. [Google Scholar] [CrossRef]
- Rivel, M.; Solano, D.; Herrera, M.; Vargas, M.; Villalta, M.; Segura, A.; Arias, A.S.; Leon, G.; Gutierrez, J.M. Pathogenesis of dermonecrosis induced by venom of the spitting cobra, Naja nigricollis: An experimental study in mice. Toxicon 2016, 119, 171–179. [Google Scholar] [CrossRef]
- Akita, E.M.; Nakai, S. Comparison of four purification methods for the production of immunoglobulins from eggs laid by hens immunized with an enterotoxigenic E. coli strain. J. Immunol. Methods 1993, 160, 207–214. [Google Scholar] [CrossRef]
- Lee, C.H.; Leu, S.J.; Lee, Y.C.; Liu, C.I.; Lin, L.T.; Mwale, P.F.; Chiang, J.R.; Tsai, B.Y.; Chen, C.C.; Hung, C.S.; et al. Characterization of Chicken-Derived Single Chain Antibody Fragments against Venom of Naja Naja Atra. Toxins 2018, 10, 383. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Lee, Y.C.; Liang, M.H.; Leu, S.J.; Lin, L.T.; Chiang, J.R.; Yang, Y.Y. Antibodies against Venom of the Snake Deinagkistrodon acutus. Appl. Environ. Microbiol. 2016, 82, 71–80. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.-C.; Wu, C.-J.; Chou, T.-Y.; Liaw, G.-W.; Hsiao, Y.-C.; Chu, L.-J.; Lee, C.-H.; Wang, P.-J.; Hsieh, C.-H.; Chen, C.-K.; et al. Development of a Monoclonal scFv against Cytotoxin to Neutralize Cytolytic Activity Induced by Naja atra Venom on Myoblast C2C12 Cells. Toxins 2022, 14, 459. https://doi.org/10.3390/toxins14070459
Liu C-C, Wu C-J, Chou T-Y, Liaw G-W, Hsiao Y-C, Chu L-J, Lee C-H, Wang P-J, Hsieh C-H, Chen C-K, et al. Development of a Monoclonal scFv against Cytotoxin to Neutralize Cytolytic Activity Induced by Naja atra Venom on Myoblast C2C12 Cells. Toxins. 2022; 14(7):459. https://doi.org/10.3390/toxins14070459
Chicago/Turabian StyleLiu, Chien-Chun, Cho-Ju Wu, Tsai-Ying Chou, Geng-Wang Liaw, Yung-Chin Hsiao, Lichieh-Julie Chu, Chi-Hsin Lee, Po-Jung Wang, Cheng-Hsien Hsieh, Chun-Kuei Chen, and et al. 2022. "Development of a Monoclonal scFv against Cytotoxin to Neutralize Cytolytic Activity Induced by Naja atra Venom on Myoblast C2C12 Cells" Toxins 14, no. 7: 459. https://doi.org/10.3390/toxins14070459
APA StyleLiu, C.-C., Wu, C.-J., Chou, T.-Y., Liaw, G.-W., Hsiao, Y.-C., Chu, L.-J., Lee, C.-H., Wang, P.-J., Hsieh, C.-H., Chen, C.-K., & Yu, J.-S. (2022). Development of a Monoclonal scFv against Cytotoxin to Neutralize Cytolytic Activity Induced by Naja atra Venom on Myoblast C2C12 Cells. Toxins, 14(7), 459. https://doi.org/10.3390/toxins14070459