Analysis of Active Compounds Using Target Protein Cofilin―Cucurbitacins in Cytotoxic Plant Bryonia cretica

 ,
,  ,
,
Abstract
1. Introduction
2. Results and Discussions
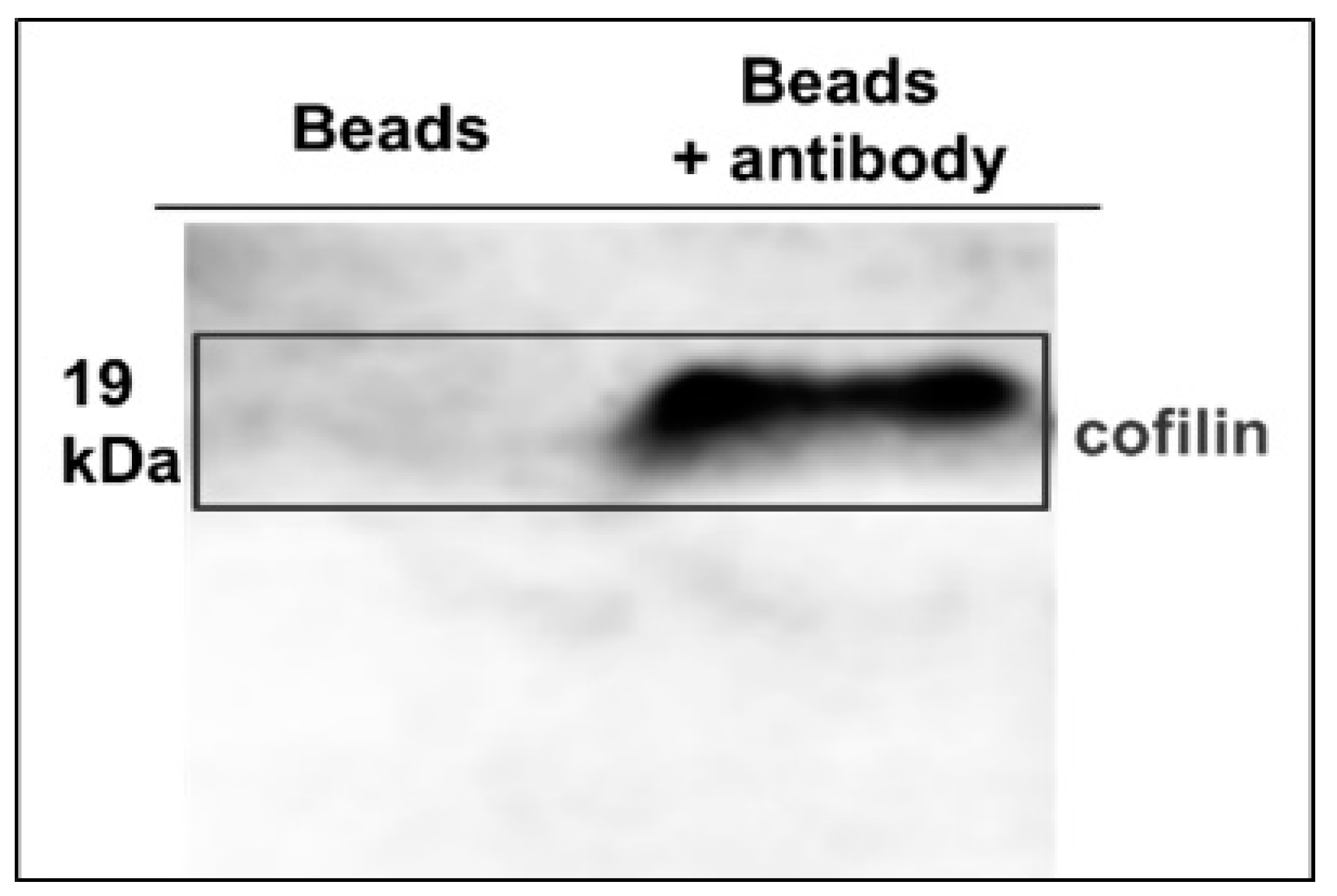
2.1. Preparation of Anti-Cofilin Antibody Binding Beads
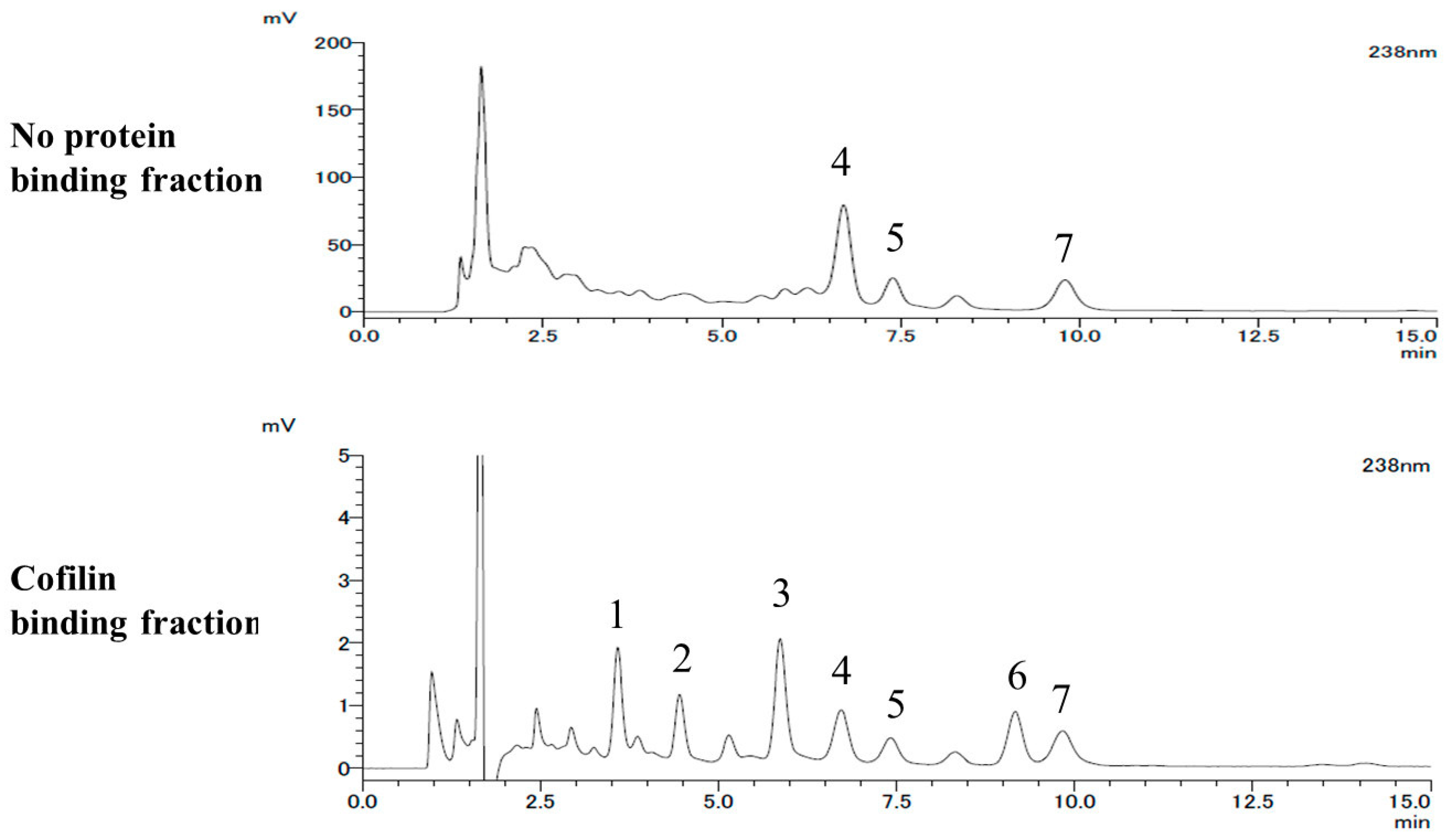
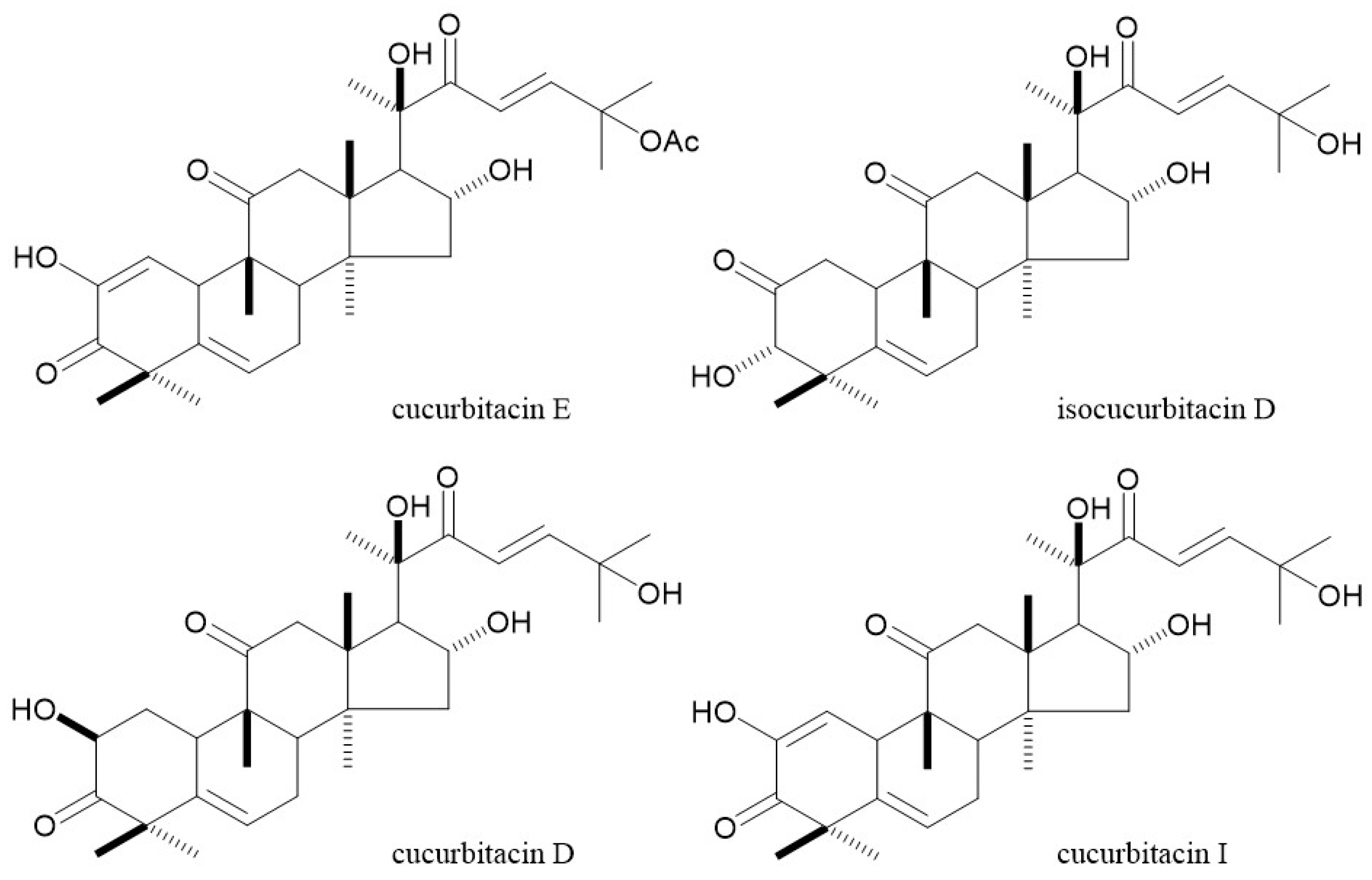
2.2. LC-MS Analysis
2.3. Anti-Proliferative Effects
2.4. Fibrous to Globular Actin Ratio in HT1080
2.5. Effects of Isocucurbitacin D on the Phosphorylation of Cofilin
3. Conclusions
4. Materials and Methods
4.1. Preparation of Anti-Cofilin Antibody Binding Beads
4.2. LC-MS Analysis
4.3. Cell Culture
4.4. Assay for Anti-Proliferative Effects
4.5. Measurement of Fibrous to Globular Actin Ratio in HT1080
4.6. Immunoblotting
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matsuda, H.; Nakashima, S.; Abdel-Halim, O.B.; Morikawa, T.; Yoshikawa, M. Cucurbitane-type triterpenes with anti-proliferative effects on U937 cells from an Egyptian natural medicine, Bryonia cretica: Structures of new triterpene glycosides, bryoniaosides A and B. Chem. Pharm. Bull. 2010, 58, 747–751. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nakashima, S.; Matsuda, H.; Kurume, A.; Oda, Y.; Nakamura, S.; Yamashita, M.; Yoshikawa, M. Cucurbitacin E as a new inhibitor of cofilin phosphorylation in human leukemia U937 cells. Bioorg. Med. Chem. Lett. 2010, 20, 2994–2997. [Google Scholar] [CrossRef]
- Gabrielsen, M.; Schuldt, M.; Munro, J.; Borucka, D.; Cameron, J.; Baugh, M.; Mleczak, A.; Lilla, S.; Morrice, N.; Olson, M.F. Cucurbitacin covalent bonding to cysteine thiols: The filamentous-actin severing protein cofilin1 as an exemplay target. Cell Commun. Signal. 2013, 11, 58. [Google Scholar] [CrossRef]
- He, X.; Gao, Q.; Qiang, Y.; Guo, W.; Ma, Y. Cucurbitacin E induces apoptosis of human prostate cancer cells via cofilin-1 and mTORC1. Oncol. Lett. 2017, 13, 4905–4910. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Tabti, R.; Elderwish, S.; Djehal, A.; Chouha, N.; Pinot, F.; Yu, P.; Nebigil, C.G.; Désaubry, L. SFPH proteins as therapeutic targets for a myriad of diseases. Bioorg. Med. Chem. Lett. 2020, 30, 127600. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Kundu, J.K.; Chae, J.; Shim, J. Targeting ROCK/LIMK/cofilin signaling pathway in cancer. Arch. Pharm. Res. 2019, 42, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Yano, M.; Nakashima, S.; Oda, Y.; Nakamura, S.; Matsuda, H. BBB-permeable aporphine-type alkaloids in Nelumbo nucifera flowers with accelerative effects on neurite outgrowth in PC-12 cells. J. Nat. Med. 2020, 74, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Nakashima, S.; Kondo, E.; Nakamura, S.; Yano, M.; Kubota, C.; Masumoto, Y.; Hirao, M.; Ogawa, Y.; Matsuda, H. Comparison of lawsone contents among Lawsonia inermis plant parts and neurite outgrowth accelerators from branches. J. Nat. Med. 2018, 72, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.Y.; Lee, S.H.; Choi, S.U.; Lee, C.O.; No, Z.; Ahn, J.W. Antitumor activity of Trichosanthes kirilowii. Arch. Pharm. Res. 1994, 17, 348–353. [Google Scholar] [CrossRef]
- Velde, V.V.; Lavie, D. 13C NMR spectroscopy of cucurbitacins. Tetrahedron 1983, 39, 317–321. [Google Scholar] [CrossRef]
- Wang, W.; Nakashima, S.; Nakamura, S.; Oda, Y.; Matsuda, H. Anti-proliferative effect of auriculataoside A on B16 melanoma 4A5 cells by suppression of Cdc42-Rac1-RhoA signaling protein levels. J. Nat. Med. 2019, 73, 450–455. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Peak Area Ratio Relative to Cucurbitacin D | |||
|---|---|---|---|
| cuD/cuD | isocuD/cuD | cuI/cuD | |
| No protein binding fraction | 100.0 ± 0.2 | 31.2 ± 0.1 | 48.5 ± 0.2 |
| Cofilin binding fraction | 100.0 ± 1.4 | 50.8 ± 0.3 | 95.7 ± 0.9 |
| Conc. (μM) | Inhibition (%) | IC50 (μM) | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 0.2 | 0.4 | 0.6 | 0.8 | 1.0 | |||
| 24 h | 100.0 ± 4.7 | 89.8 ± 1.8 | 75.8 ± 2.5 ** | 63.6 ± 2.2 ** | 30.2 ± 2.2 ** | 29.0 ± 1.0 ** | 0.66 | |
| 48 h | 100.0 ± 0.8 | 94.2 ± 1.5 | 75.8 ± 2.9 ** | 63.4 ± 1.8 ** | 26.3 ± 1.2 ** | 20.8 ± 0.6 ** | 0.64 | |
| 72 h | 100.0 ± 2.4 | 106.3 ± 4.4 | 98.2 ± 4.9 | 87.9 ± 7.8 | 25.1 ± 1.7 ** | 15.4 ± 0.1 ** | 0.71 | |
| Conc. (μM) | F/G Ratio (%) | ||||
|---|---|---|---|---|---|
| 0 | 0.01 | 0.1 | 1 | 10 | |
| 100.0 ± 9.3 | 74.1 ± 10.8 | 65.8 ± 6.5 ** | 77.1 ± 7.7 | 60.0 ± 16.1 | |
| Conc. (nM) | Expression Ratio (%) | ||||
|---|---|---|---|---|---|
| 0 | 1 | 10 | 100 | 1000 | |
| p-cofilin/β-actin | 100 | 81.2 * | 82.2 * | 73.3 ** | 61.7 ** |
| p-LIMK1/β-actin | 100 | 161.6 ** | 211.4 ** | 161.5 ** | 131.1 * |
| p-LIMK2/β-actin | 100 | 140.4 * | 164.2 ** | 149.1 ** | 85.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakashima, S.; Oda, Y.; Morita, M.; Ohta, A.; Morikawa, T.; Matsuda, H.; Nakamura, S. Analysis of Active Compounds Using Target Protein Cofilin―Cucurbitacins in Cytotoxic Plant Bryonia cretica. Toxins 2022, 14, 212. https://doi.org/10.3390/toxins14030212
Nakashima S, Oda Y, Morita M, Ohta A, Morikawa T, Matsuda H, Nakamura S. Analysis of Active Compounds Using Target Protein Cofilin―Cucurbitacins in Cytotoxic Plant Bryonia cretica. Toxins. 2022; 14(3):212. https://doi.org/10.3390/toxins14030212
Chicago/Turabian StyleNakashima, Souichi, Yoshimi Oda, Moeko Morita, Ayako Ohta, Toshio Morikawa, Hisashi Matsuda, and Seikou Nakamura. 2022. "Analysis of Active Compounds Using Target Protein Cofilin―Cucurbitacins in Cytotoxic Plant Bryonia cretica" Toxins 14, no. 3: 212. https://doi.org/10.3390/toxins14030212
APA StyleNakashima, S., Oda, Y., Morita, M., Ohta, A., Morikawa, T., Matsuda, H., & Nakamura, S. (2022). Analysis of Active Compounds Using Target Protein Cofilin―Cucurbitacins in Cytotoxic Plant Bryonia cretica. Toxins, 14(3), 212. https://doi.org/10.3390/toxins14030212