Protective Effects of Taraxasterol against Deoxynivalenol-Induced Damage to Bovine Mammary Epithelial Cells


Abstract
:1. Introduction
2. Results
2.1. Taraxasterol Improved the Decrease in Bovine Mammary Epithelial Cell Viability Induced by DON
2.2. Taraxasterol Alleviated the Cellular Damage Caused by DON by Reducing LDH Levels
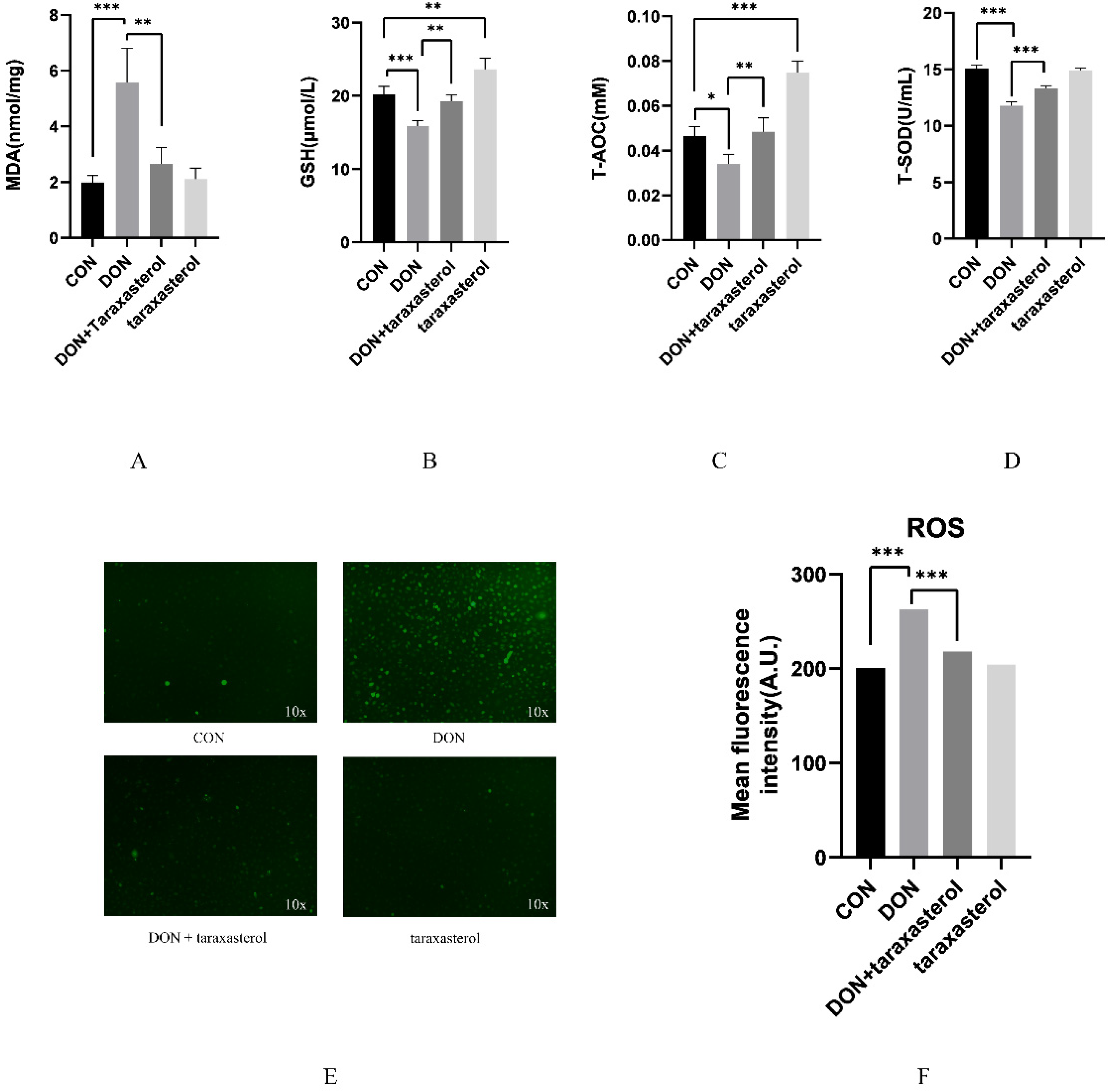
2.3. Taraxasterol Prevented Oxidative Stress Induced by DON
2.4. The Protective Effects of Taraxasterol against the DON-Induced Decrease in Mitochondrial Membrane Potential (ΔΨm, MMP)
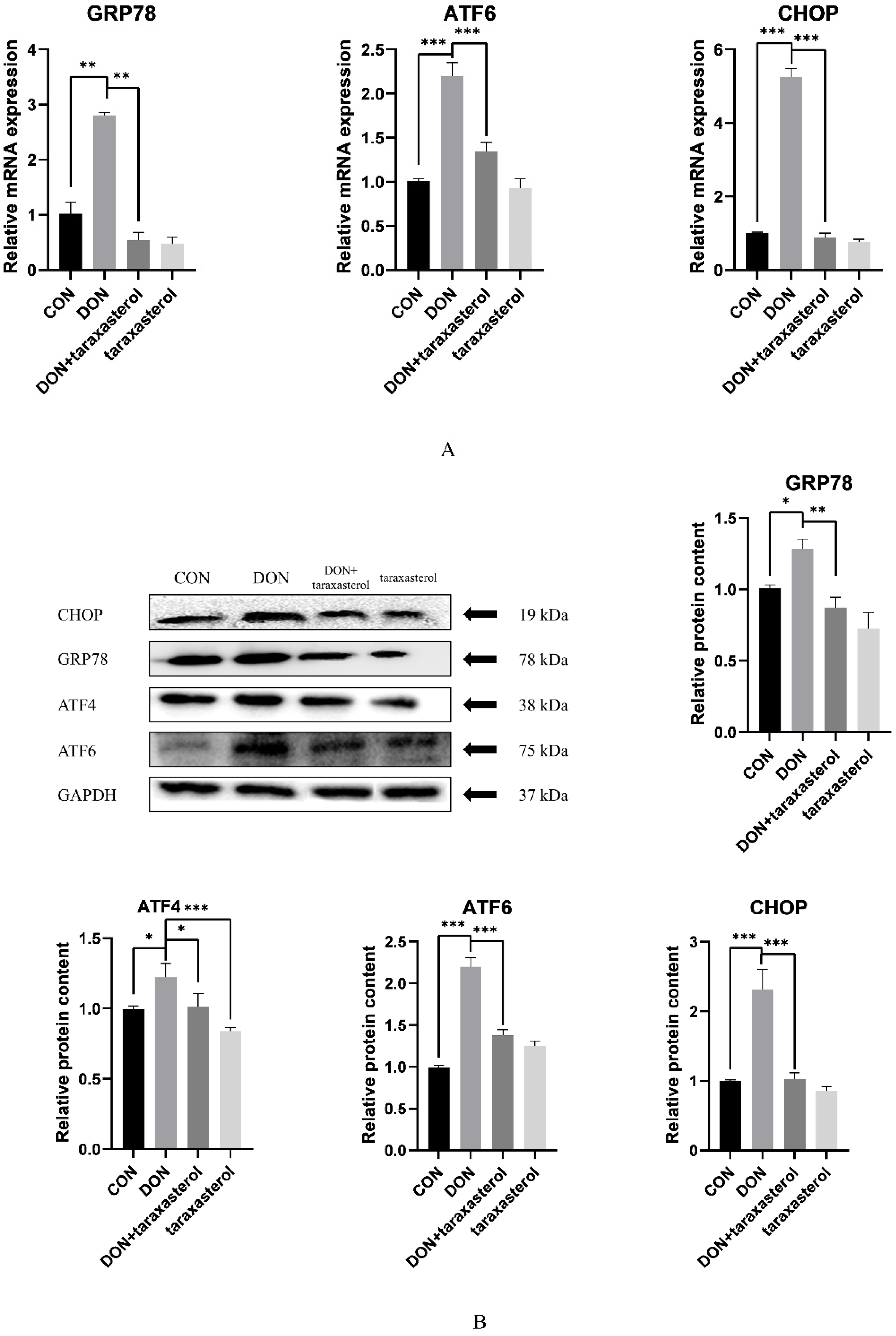
2.5. Taraxasterol Alleviated ER Stress Caused by DON
2.6. Taraxasterol Alleviated the Cell Apoptosis Induced by DON
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Cell Culture
5.3. Cell Treatment
5.4. Cell Viability Assay
5.5. LDH Assay
5.6. Measurement of Malondialdehyde (MDA), Glutathione (GSH), Total Antioxidant Capacity (T-AOC) and Total Superoxide Dismutase (T-SOD) Levels
5.7. Measurement of ROS Production
5.8. Mitochondrial Membrane Potential Assay
5.9. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (PCR)
5.10. Western Blotting
5.11. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ha, E.; Zemel, M.B. Functional properties of whey, whey components, and essential amino acids: Mechanisms underlying health benefits for active people (review). J. Nutr. Biochem. 2003, 14, 251–258. [Google Scholar] [CrossRef]
- Yao, Y.; Long, M. The biological detoxification of deoxynivalenol: A review. Food Chem. Toxicol. 2020, 145, 111649. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Kuča, K.; Humpf, H.-U.; Klímová, B.; Cramer, B. Fate of deoxynivalenol and deoxynivalenol-3-glucoside during cereal-based thermal food processing: A review study. Mycotoxin Res. 2016, 33, 79–91. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; DiNovi, M.; Grasl-Kraupp, B.; et al. Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, e04718. [Google Scholar] [CrossRef] [PubMed]
- Viegas, S.; Assunção, R.; Twarużek, M.; Kosicki, R.; Grajewski, J.; Viegas, C. Mycotoxins feed contamination in a dairy farm—Potential implications for milk contamination and workers’ exposure in a One Health approach. J. Sci. Food Agric. 2019, 100, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, Q.-C.; Zhu, C.-C.; Liu, J.; Zhang, Y.; Cui, X.-S.; Kim, N.-H.; Sun, S.-C. Deoxynivalenol exposure induces autophagy/apoptosis and epigenetic modification changes during porcine oocyte maturation. Toxicol. Appl. Pharmacol. 2016, 300, 70–76. [Google Scholar] [CrossRef]
- Borutova, R.; Faix, S.; Placha, I.; Gresakova, L.; Cobanova, K.; Leng, L. Effects of deoxynivalenol and zearalenone on oxidative stress and blood phagocytic activity in broilers. Arch. Anim. Nutr. 2008, 62, 303–312. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Lim, W.; Park, S.; Kim, J.; You, S.; Song, G. Deoxynivalenol induces apoptosis and disrupts cellular homeostasis through MAPK signaling pathways in bovine mammary epithelial cells. Environ. Pollut. 2019, 252, 879–887. [Google Scholar] [CrossRef]
- Fishbein, A.; Wang, W.; Yang, H.; Yang, J.; Hallisey, V.M.; Deng, J.; Verheul, S.M.L.; Hwang, S.H.; Gartung, A.; Wang, Y.; et al. Resolution of eicosanoid/cytokine storm prevents carcinogen and inflammation-initiated hepatocellular cancer progression. Proc. Natl. Acad. Sci. USA 2020, 117, 21576–21587. [Google Scholar] [CrossRef]
- Fu, Y.; Jin, Y.; Zhao, Y.; Shan, A.; Fang, H.; Shen, J.; Zhou, C.; Yu, H.; Zhou, Y.F.; Wang, X.; et al. Zearalenone induces apoptosis in bovine mammary epithelial cells by activating endoplasmic reticulum stress. J. Dairy Sci. 2019, 102, 10543–10553. [Google Scholar] [CrossRef]
- Zafar, R.; Sharma, K. Occurrence of taraxerol and taraxasterol in medicinal plants. Pharmacogn. Rev. 2015, 9, 19–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durrer, M.; Mevissen, M.; Holinger, M.; Hamburger, M.; Graf-Schiller, S.; Mayer, P.; Potterat, O.; Bruckmaier, R.; Walkenhorst, M. Effects of a Multicomponent Herbal Extract on the Course of Subclinical Ketosis in Dairy Cows—A Blinded Placebo-controlled Field-study. Planta Med. 2020, 86, 1375–1388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xiong, H.; Liu, L. Effects of taraxasterol on inflammatory responses in lipopolysaccharide-induced RAW 264.7 macrophages. J. Ethnopharmacol. 2012, 141, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Piao, T.; Ma, Z.; Li, X.; Liu, J. Taraxasterol inhibits IL-1β-induced inflammatory response in human osteoarthritic chondrocytes. Eur. J. Pharmacol. 2015, 756, 38–42. [Google Scholar] [CrossRef]
- Koo, H.-N.; Hong, S.-H.; Song, B.-K.; Kim, C.-H.; Yoo, Y.-H.; Kim, H.-M. Taraxacum officinale induces cytotoxicity through TNF-α and IL-1α secretion in Hep G2 cells. Life Sci. 2004, 74, 1149–1157. [Google Scholar] [CrossRef]
- Guo, J.-B.; Ye, H.-H.; Chen, J.-F. Anti-proliferation Effect of Taraxacum mongolicum Extract in HepG2 Cells and Its Mechanism. J. Chin. Med. Mater. 2015, 38, 2129–2133. [Google Scholar]
- Yoon, J.-Y.; Cho, H.-S.; Lee, J.-J.; Lee, H.-J.; Jun, S.Y.; Lee, J.-H.; Song, H.-H.; Choi, S.; Saloura, V.; Gil Park, C.; et al. Novel TRAIL sensitizer Taraxacum officinale F.H. Wigg enhances TRAIL-induced apoptosis in Huh7 cells. Mol. Carcinog. 2015, 55, 387–396. [Google Scholar] [CrossRef]
- Xu, L.; Yu, Y.; Sang, R.; Li, J.; Ge, B.; Zhang, X. Protective Effects of Taraxasterol against Ethanol-Induced Liver Injury by Regulating CYP2E1/Nrf2/HO-1 and NF-κB Signaling Pathways in Mice. Oxid. Med. Cell. Longev. 2018, 2018, 8284107. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Xiong, H.; Li, H.; Cheng, Y. Protective effect of taraxasterol against LPS-induced endotoxic shock by modulating inflammatory responses in mice. Immunopharmacol. Immunotoxicol. 2013, 36, 11–16. [Google Scholar] [CrossRef]
- Iqbal, S.Z.; Usman, S.; Razis, A.F.A.; Ali, N.B.; Saif, T.; Asi, M.R. Assessment of Deoxynivalenol in Wheat, Corn and Its Products and Estimation of Dietary Intake. Int. J. Environ. Res. Public Health 2020, 17, 5602. [Google Scholar] [CrossRef]
- Wang, J.; Jin, Y.; Wu, S.; Yu, H.; Zhao, Y.; Fang, H.; Shen, J.; Zhou, C.; Fu, Y.; Li, R.; et al. Deoxynivalenol induces oxidative stress, inflammatory response and apoptosis in bovine mammary epithelial cells. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1663–1674. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Dong, X.; Meng, X. Anti-Inflammatory Effects of Taraxasterol on LPS-Stimulated Human Umbilical Vein Endothelial Cells. Inflammation 2018, 41, 1755–1761. [Google Scholar] [CrossRef] [PubMed]
- Schütz, K.; Carle, R.; Schieber, A. Taraxacum—A review on its phytochemical and pharmacological profile. J. Ethnopharmacol. 2006, 107, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; He, Z.; Wang, J.; Xin, Z.; Wang, J.; Li, F.; Fu, Y. Taraxasterol Inhibits LPS-Induced Inflammatory Response in BV2 Microglia Cells by Activating LXRα. Front. Pharmacol. 2018, 9, 278. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P. Analysis of Cell Viability by the Lactate Dehydrogenase Assay. Cold Spring Harb. Protoc. 2018, 6, 465–468. [Google Scholar] [CrossRef]
- Wallace, D.C. Mitochondrial Diseases in Man and Mouse. Science 1999, 283, 1482–1488. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, J.S.; Steinauer, K.K.; Hornung, B.; Irish, J.; Lecane, P.; Birrell, G.; Peehl, D.M.; Knox, S.J. Role of glutathione depletion and reactive oxygen species generation in apoptotic signaling in a human B lymphoma cell line. Cell Death Differ. 2002, 9, 252–263. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, J.; Fang, H.; Yu, H.; Zhao, Y.; Shen, J.; Zhou, C.; Jin, Y. Pterostilbene inhibits deoxynivalenol-induced oxidative stress and inflammatory response in bovine mammary epithelial cells. Toxicon 2020, 189, 10–18. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial Reactive Oxygen Species (ROS) and ROS-Induced ROS Release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [Green Version]
- Simon, H.-U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Quan, Z.; Gu, J.; Dong, P.; Lu, J.; Wu, X.; Wu, W.; Fei, X.; Li, S.; Wang, Y.; Wang, J.; et al. Reactive oxygen species-mediated endoplasmic reticulum stress and mitochondrial dysfunction contribute to cirsimaritin-induced apoptosis in human gallbladder carcinoma GBC-SD cells. Cancer Lett. 2010, 295, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, T.; Arnold, S.M.; Miller, C.N.; Wu, J.; Li, J.; Gunnison, K.M.; Mori, K.; Akha, A.A.S.; Raden, D.; Kaufman, R.J. Adaptation to ER Stress Is Mediated by Differential Stabilities of Pro-Survival and Pro-Apoptotic mRNAs and Proteins. PLoS Biol. 2006, 4, e374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanvictores, T.; Davis, D.D. Histology, Rough Endoplasmic Reticulum; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Voeltz, G.K.; Prinz, W.A.; Shibata, Y.; Rist, J.M.; Rapoport, T.A. A Class of Membrane Proteins Shaping the Tubular Endoplasmic Reticulum. Cell 2006, 124, 573–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, H.-O.; Yadav, R.K.; Kim, H.-R.; Chae, H.-J. ER stress: Autophagy induction, inhibition and selection. Autophagy 2015, 11, 1956–1977. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Han, Z.; Couvillon, A.D.; Exton, J.H. Critical Role of Endogenous Akt/IAPs and MEK1/ERK Pathways in Counteracting Endoplasmic Reticulum Stress-induced Cell Death. J. Biol. Chem. 2004, 279, 49420–49429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimball, S.R.; Jefferson, L.S. Induction of REDD1 gene expression in the liver in response to endoplasmic reticulum stress is mediated through a PERK, eIF2α phosphorylation, ATF4-dependent cascade. Biochem. Biophys. Res. Commun. 2012, 427, 485–489. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | GenBank Accession No. | Product Size (bp) |
|---|---|---|---|---|
| β-actin | 5′-CCCTGGAGAAGAGCTACGAG-3′ | 5′-GTAGTTTCGTGAATGCCGCAG-3′ | NM_173979.3 | 130 |
| GRP78 | 5′-CGACCCCTGACGAAAGACAA-3′ | 5′-AGGTGTCAGGCGATTTTGGT-3′ | NM_001075148.1 | 198 |
| ATF6 | 5′-ATATTCCTCCGCCTCCCTGT-3′ | 5′-GTCCTTTCCACTTCGTGCCT-3′ | XM_024989876.1 | 103 |
| CHOP | 5′-GAGCTGGAAGCCTGGTATGA-3′ | 5′-CTCCTTGTTTCCAGGGGGTG-3′ | NM_001078163.1 | 90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zheng, K.; Jin, Y.; Fu, Y.; Wang, R.; Zhang, J. Protective Effects of Taraxasterol against Deoxynivalenol-Induced Damage to Bovine Mammary Epithelial Cells. Toxins 2022, 14, 211. https://doi.org/10.3390/toxins14030211
Wang J, Zheng K, Jin Y, Fu Y, Wang R, Zhang J. Protective Effects of Taraxasterol against Deoxynivalenol-Induced Damage to Bovine Mammary Epithelial Cells. Toxins. 2022; 14(3):211. https://doi.org/10.3390/toxins14030211
Chicago/Turabian StyleWang, Junxiong, Kexin Zheng, Yongcheng Jin, Yurong Fu, Rui Wang, and Jing Zhang. 2022. "Protective Effects of Taraxasterol against Deoxynivalenol-Induced Damage to Bovine Mammary Epithelial Cells" Toxins 14, no. 3: 211. https://doi.org/10.3390/toxins14030211
APA StyleWang, J., Zheng, K., Jin, Y., Fu, Y., Wang, R., & Zhang, J. (2022). Protective Effects of Taraxasterol against Deoxynivalenol-Induced Damage to Bovine Mammary Epithelial Cells. Toxins, 14(3), 211. https://doi.org/10.3390/toxins14030211