Recent Progress in Rapid Determination of Mycotoxins Based on Emerging Biorecognition Molecules: A Review

Abstract
:1. Introduction
2. Biorecognition Molecules
2.1. Antibodies
2.2. Aptamers
2.3. Short Peptides
2.4. Molecularly Imprinted Polymers (MIPs)
3. Areas of Application of Recognition Elements for Detection of Mycotoxins
3.1. Sample Preparation
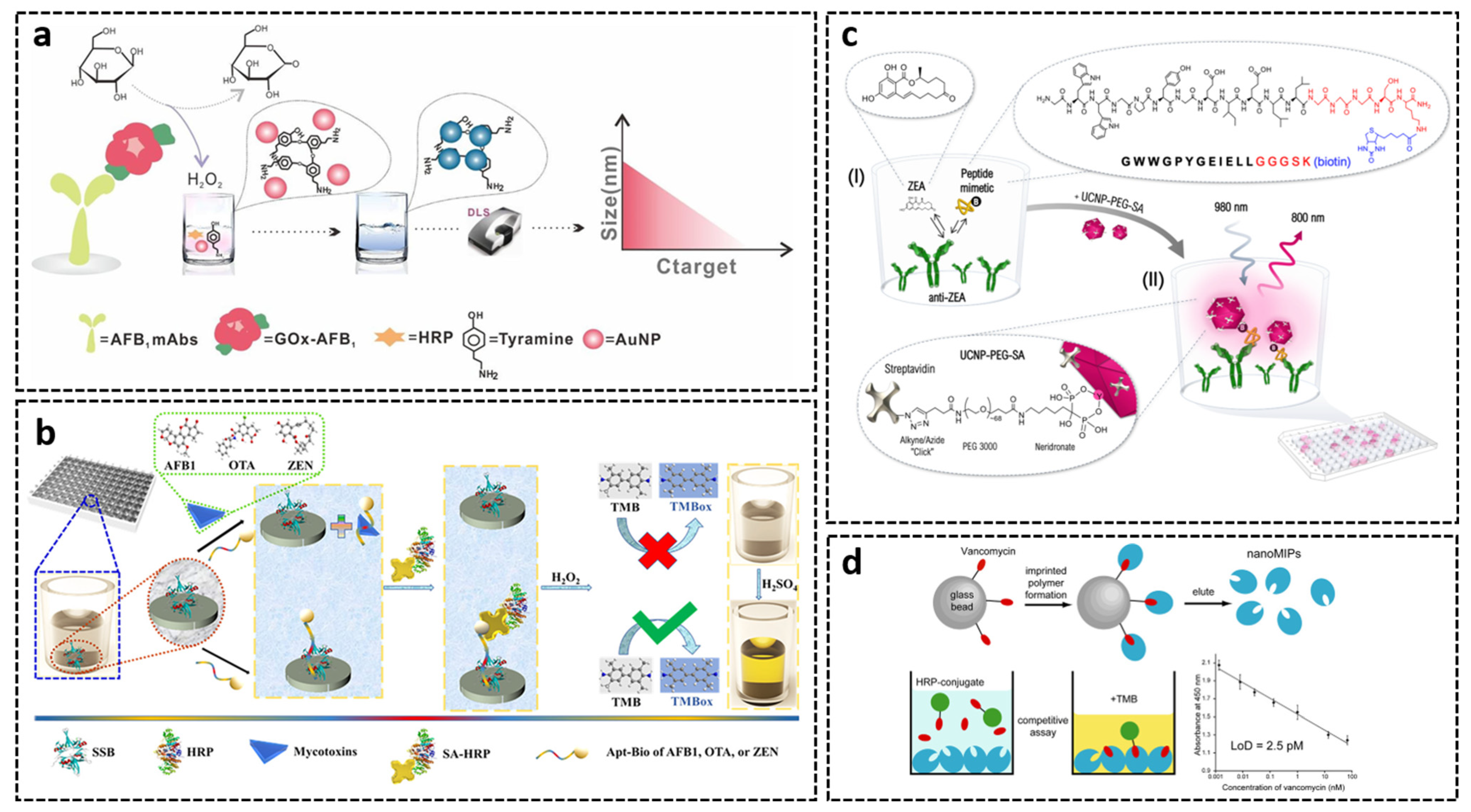
3.2. Microplate- and Tube-Based Assays
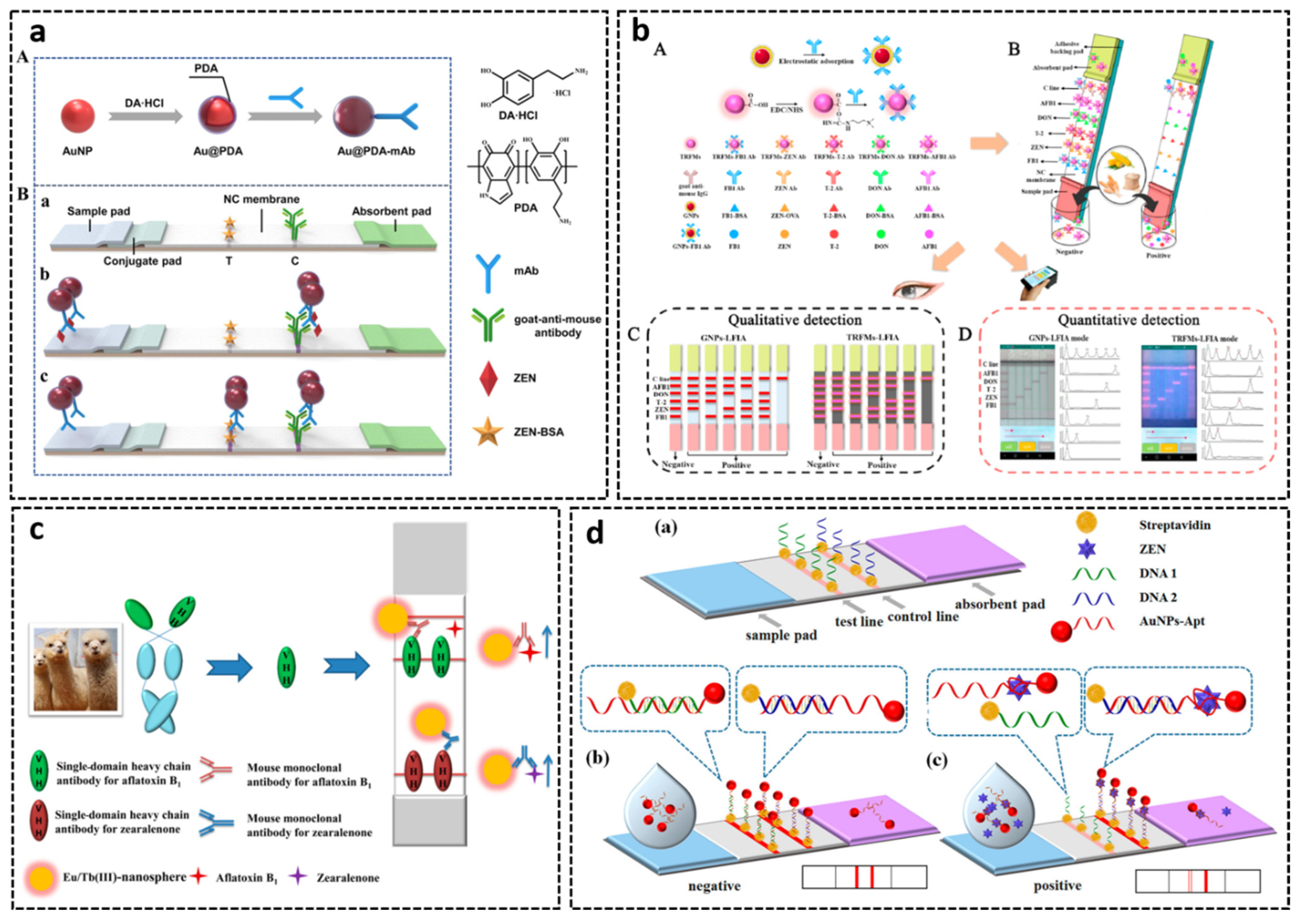
3.3. Lateral Flow Assays
3.4. Biosensors

4. Commercial Biorecognition Elements, Test Kits, and Analysis Systems for Mycotoxins Detection
4.1. Mycotoxin Antibodies
4.2. Microplate or Tube ELISA Test Kits
4.3. Lateral Flow Assays
4.4. Immunoaffinity Columns
4.5. Multiplexed Analysis Platforms
5. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in Occurrence, Importance, and Mycotoxin Control Strategies: Prevention and Detoxification in Foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef]
- Rai, A.; Das, M.; Tripathi, A. Occurrence and toxicity of a fusarium mycotoxin, zearalenone. Crit. Rev. Food Sci. Nutr. 2020, 60, 2710–2729. [Google Scholar] [CrossRef]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef]
- Pleadin, J.; Frece, J.; Markov, K. Mycotoxins in food and feed. In Advances in Food and Nutrition Research; Academic Press Inc.: Cambridge, MA, USA, 2019; Volume 89, pp. 297–345. ISBN 9780128171714. [Google Scholar]
- Al-Jaal, B.; Salama, S.; Al-Qasmi, N.; Jaganjac, M. Mycotoxin contamination of food and feed in the Gulf Cooperation Council countries and its detection. Toxicon 2019, 171, 43–50. [Google Scholar] [CrossRef]
- Misihairabgwi, J.M.; Ezekiel, C.N.; Sulyok, M.; Shephard, G.S.; Krska, R. Mycotoxin contamination of foods in Southern Africa: A 10-year review (2007–2016). Crit. Rev. Food Sci. Nutr. 2019, 59, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global mycotoxin occurrence in feed: A ten-year survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, I.; Naehrer, K. A three-year survey on the worldwide occurrence of mycotoxins in feedstuffs and feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef]
- Williams, J.; Phillips, T.; Jolly, P.E.; Stiles, J.; Jolly, C.; Aggarwal, D. Human aflatoxicosis in developing countries: A review of toxicology, exposure, potential health consequences, and interventions. Am. J. Clin. Nutr. 2004, 80, 22–1106. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Gonçalves, B.L.; de Neeff, D.V.; Ponzilacqua, B.; Coppa, C.F.S.C.; Hintzsche, H.; Sajid, M.; Cruz, A.G.; Corassin, C.H.; Oliveira, C.A.F. Aflatoxin in foodstuffs: Occurrence and recent advances in decontamination. Food Res. Int. 2018, 113, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Mousavi Khaneghah, A.; Moosavi, M.; Omar, S.S.; Oliveira, C.A.F.; Karimi-Dehkordi, M.; Fakhri, Y.; Huseyn, E.; Nematollahi, A.; Farahani, M.; Sant’Ana, A.S. The prevalence and concentration of aflatoxin M1 among different types of cheeses: A global systematic review, meta-analysis, and meta-regression. Food Control 2021, 125, 107960. [Google Scholar] [CrossRef]
- Jørgensen, K. Occurrence of ochratoxin A in commodities and processed food - A review of EU occurrence data. Food Addit. Contam. 2005, 22, 26–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponsone, M.L.; Chiotta, M.L.; Palazzini, J.M.; Combina, M.; Chulze, S. Control of ochratoxin a production in grapes. Toxins 2012, 4, 364–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, P.M. Recent research on fumonisins: A review. Food Addit. Contam. Part A 2012, 29, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Schwake-Anduschus, C.; Langenkämper, G.; Unbehend, G.; Dietrich, R.; Märtlbauer, E.; Münzing, K. Occurrence of Fusarium T-2 and HT-2 toxins in oats from cultivar studies in Germany and degradation of the toxins during grain cleaning treatment and food processing. Food Addit. Contam. Part A 2010, 27, 1253–1260. [Google Scholar] [CrossRef] [Green Version]
- Kiš, M.; Vulić, A.; Kudumija, N.; Šarkanj, B.; Jaki Tkalec, V.; Aladić, K.; Škrivanko, M.; Furmeg, S.; Pleadin, J. A Two-Year Occurrence of Fusarium T-2 and HT-2 Toxin in Croatian Cereals Relative of the Regional Weather. Toxins 2021, 13, 39. [Google Scholar] [CrossRef] [PubMed]
- Karami-Osboo, R.; Mirabolfathy, M.; Aliakbari, F. Natural deoxynivalenol contamination of corn produced in Golestan and Moqan areas in Iran. J. Agric. Sci. Technol. 2010, 12, 233–239. [Google Scholar]
- Wang, L.; Liao, Y.; Peng, Z.; Chen, L.; Zhang, W.; Nüssler, A.K.; Shi, S.; Liu, L.; Yang, W. Food raw materials and food production occurrences of deoxynivalenol in different regions. Trends Food Sci. Technol. 2019, 83, 41–52. [Google Scholar] [CrossRef]
- Mahato, D.K.; Devi, S.; Pandhi, S.; Sharma, B.; Maurya, K.K.; Mishra, S.; Dhawan, K.; Selvakumar, R.; Kamle, M.; Mishra, A.K.; et al. Occurrence, impact on agriculture, human health, and management strategies of zearalenone in food and feed: A review. Toxins 2021, 13, 92. [Google Scholar] [CrossRef]
- Ropejko, K.; Twarużek, M. Zearalenone and Its Metabolites-General Overview, Occurrence, and Toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef]
- Silva, L.J.G.; Pereira, A.M.P.T.; Pena, A.; Lino, C.M. Citrinin in foods and supplements: A review of occurrence and analytical methodologies. Foods 2021, 10, 14. [Google Scholar] [CrossRef]
- Saleh, I.; Goktepe, I. The characteristics, occurrence, and toxicological effects of patulin. Food Chem. Toxicol. 2019, 129, 301–311. [Google Scholar] [CrossRef]
- Mahato, D.K.; Kamle, M.; Sharma, B.; Pandhi, S.; Devi, S.; Dhawan, K.; Selvakumar, R.; Mishra, D.; Kumar, A.; Arora, S.; et al. Patulin in food: A mycotoxin concern for human health and its management strategies. Toxicon 2021, 198, 12–23. [Google Scholar] [CrossRef]
- Agriopoulou, S. Ergot alkaloids mycotoxins in cereals and cereal-derived food products: Characteristics, toxicity, prevalence, and control strategies. Agronomy 2021, 11, 931. [Google Scholar] [CrossRef]
- Krska, R.; Crews, C. Significance, chemistry and determination of ergot alkaloids: A review. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2008, 25, 722–731. [Google Scholar] [CrossRef]
- Shephard, G.S. Impact of mycotoxins on human health in developing countries. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2008, 25, 146–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anfossi, L.; Giovannoli, C.; Baggiani, C. Mycotoxin detection. Curr. Opin. Biotechnol. 2016, 37, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Peng, Z.; Chen, L.; Nüssler, A.K.; Liu, L.; Yang, W. Deoxynivalenol, gut microbiota and immunotoxicity: A potential approach? Food Chem. Toxicol. 2018, 112, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Frizzell, C.; Ndossi, D.; Verhaegen, S.; Dahl, E.; Eriksen, G.; Sørlie, M.; Ropstad, E.; Muller, M.; Elliott, C.T.; Connolly, L. Endocrine disrupting effects of zearalenone, alpha- and beta-zearalenol at the level of nuclear receptor binding and steroidogenesis. Toxicol. Lett. 2011, 206, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Amuzie, C.J.; Pestka, J.J. Suppression of insulin-like growth factor acid-labile subunit expression-A novel mechanism for deoxynivalenol-induced growth retardation. Toxicol. Sci. 2009, 113, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Guo, P.; Liu, A.; Wu, Q.; Xue, X.; Dai, M.; Hao, H.; Qu, W.; Xie, S.; Wang, X.; et al. Nitric oxide (NO)-mediated mitochondrial damage plays a critical role in T-2 toxin-induced apoptosis and growth hormone deficiency in rat anterior pituitary GH3 cells. Food Chem. Toxicol. 2017, 102, 11–23. [Google Scholar] [CrossRef]
- Degen, G.H.; Ali, N.; Gundert-Remy, U. Preliminary data on citrinin kinetics in humans and their use to estimate citrinin exposure based on biomarkers. Toxicol. Lett. 2018, 282, 43–48. [Google Scholar] [CrossRef]
- Song, E.; Xia, X.; Su, C.; Dong, W.; Xian, Y.; Wang, W.; Song, Y. Hepatotoxicity and genotoxicity of patulin in mice, and its modulation by green tea polyphenols administration. Food Chem. Toxicol. 2014, 71, 122–127. [Google Scholar] [CrossRef]
- Zhu, X.; Zeng, Z.; Chen, Y.; Li, R.; Tang, X.; Zhu, X.; Huo, J.; Liu, Y.; Zhang, L.; Chen, J. Genotoxicity of three mycotoxin contaminants of rice: 28-day multi-endpoint assessment in rats. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2021, 867. [Google Scholar] [CrossRef]
- Rašić, D.; Želježić, D.; Kopjar, N.; Kifer, D.; Šegvić Klarić, M.; Peraica, M. DNA damage in rat kidneys and liver upon subchronic exposure to single and combined ochratoxin A and citrinin. World Mycotoxin J. 2019, 12, 163–172. [Google Scholar] [CrossRef]
- Zhou, H.; George, S.; Hay, C.; Lee, J.; Qian, H.; Sun, X. Individual and combined effects of Aflatoxin B1, Deoxynivalenol and Zearalenone on HepG2 and RAW 264.7 cell lines. Food Chem. Toxicol. 2017, 103, 18–27. [Google Scholar] [CrossRef]
- Wan, L.Y.M.; Turner, P.C.; El-Nezami, H. Individual and combined cytotoxic effects of Fusarium toxins (deoxynivalenol, nivalenol, zearalenone and fumonisins B1) on swine jejunal epithelial cells. Food Chem. Toxicol. 2013, 57, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.H.; Lei, M.Y.; Zhang, N.Y.; Gao, X.; Li, C.; Krumm, C.S.; Qi, D.S. Individual and combined cytotoxic effects of aflatoxin B1, zearalenone, deoxynivalenol and fumonisin B1on BRL 3A rat liver cells. Toxicon 2015, 95, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef]
- Liu, D.; Ge, L.; Wang, Q.; Su, J.; Chen, X.; Wang, C.; Huang, K. Low-level contamination of deoxynivalenol: A threat from environmental toxins to porcine epidemic diarrhea virus infection. Environ. Int. 2020, 143, 105949. [Google Scholar] [CrossRef]
- Kuiper-Goodman, T.; Scott, P.M.; Watanabe, H. Risk assessment of the mycotoxin zearalenone. Regul. Toxicol. Pharmacol. 1987, 7, 253–306. [Google Scholar] [CrossRef]
- Tao, Y.; Xie, S.; Xu, F.; Liu, A.; Wang, Y.; Chen, D.; Pan, Y.; Huang, L.; Peng, D.; Wang, X.; et al. Ochratoxin A: Toxicity, oxidative stress and metabolism. Food Chem. Toxicol. 2018, 112, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Ekwomadu, T.I.; Akinola, S.A.; Mwanza, M. Fusarium mycotoxins, their metabolites (Free, emerging, and masked), food safety concerns, and health impacts. Int. J. Environ. Res. Public Health 2021, 18, 11741. [Google Scholar] [CrossRef]
- Li, Q.; Yuan, Q.; Wang, T.; Zhan, Y.; Yang, L.; Fan, Y.; Lei, H.; Su, J. Fumonisin B1 Inhibits Cell Proliferation and Decreases Barrier Function of Swine Umbilical Vein Endothelial Cells. Toxins 2021, 13, 863. [Google Scholar] [CrossRef]
- Nathanail, A.V.; Varga, E.; Meng-Reiterer, J.; Bueschl, C.; Michlmayr, H.; Malachova, A.; Fruhmann, P.; Jestoi, M.; Peltonen, K.; Adam, G.; et al. Metabolism of the Fusarium Mycotoxins T-2 Toxin and HT-2 Toxin in Wheat. J. Agric. Food Chem. 2015, 63, 7862–7872. [Google Scholar] [CrossRef] [PubMed]
- Speijers, G.J.A.; Franken, M.A.M.; van Leeuwen, F.X.R. Subacute toxicity study of patulin in the rat: Effects on the kidney and the gastro-intestinal tract. Food Chem. Toxicol. 1988, 26, 23–30. [Google Scholar] [CrossRef]
- de Oliveira Filho, J.W.G.; Islam, M.T.; Ali, E.S.; Uddin, S.J.; de Oliveira Santos, J.V.; de Alencar, M.V.O.B.; Júnior, A.L.G.; Paz, M.F.C.J.; de Brito, M.d.R.M.; e Sousa, J.M.d.C.; et al. A comprehensive review on biological properties of citrinin. Food Chem. Toxicol. 2017, 110, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Zhang, X.; Shen, H.; Ou-Yang, Z.; Yang, M. Validation of a gas chromatography-electron capture detection of T-2 and HT-2 toxins in Chinese herbal medicines and related products after immunoaffinity column clean-up and pre-column derivatization. Food Chem. 2012, 132, 574–581. [Google Scholar] [CrossRef]
- Schothorst, R.C.; Jekel, A.A. Determination of trichothecenes in wheat by capillary gas chromatography with flame ionisation detection. Food Chem. 2001, 73, 111–117. [Google Scholar] [CrossRef]
- Tanaka, T.; Yoneda, A.; Inoue, S.; Sugiura, Y.; Ueno, Y. Simultaneous determination of trichothecene mycotoxins and zearalenone in cereals by gas chromatography-mass spectrometry. J. Chromatogr. A 2000, 882, 23–28. [Google Scholar] [CrossRef]
- Hussain, S.; Asi, M.R.; Iqbal, M.; Khalid, N.; Wajih-Ul-Hassan, S.; Ariño, A. Patulin mycotoxin in mango and orange fruits, juices, pulps, and jams marketed in Pakistan. Toxins 2020, 12, 52. [Google Scholar] [CrossRef] [Green Version]
- Moez, E.; Noel, D.; Brice, S.; Benjamin, G.; Pascaline, A.; Didier, M. Aptamer assisted ultrafiltration cleanup with high performance liquid chromatography-fluorescence detector for the determination of OTA in green coffee. Food Chem. 2020, 310, 125851. [Google Scholar] [CrossRef]
- Tang, Y.; Mu, L.; Cheng, J.; Du, Z.; Yang, Y. Determination of Multi-Class Mycotoxins in Apples and Tomatoes by Combined Use of QuEChERS Method and Ultra-High-Performance Liquid Chromatography Tandem Mass Spectrometry. Food Anal. Methods 2020, 13, 1381–1390. [Google Scholar] [CrossRef]
- Pantano, L.; La Scala, L.; Olibrio, F.; Galluzzo, F.G.; Bongiorno, C.; Buscemi, M.D.; Macaluso, A.; Vella, A. Quechers LC-MS/MS screening method for mycotoxin detection in cereal products and spices. Int. J. Environ. Res. Public Health 2021, 18, 3774. [Google Scholar] [CrossRef] [PubMed]
- Janik, E.; Niemcewicz, M.; Podogrocki, M.; Ceremuga, M.; Gorniak, L.; Stela, M.; Bijak, M. The existing methods and novel approaches in mycotoxins’ detection. Molecules 2021, 26, 3981. [Google Scholar] [CrossRef]
- Iqbal, S.Z. Mycotoxins in food, recent development in food analysis and future challenges; a review. Curr. Opin. Food Sci. 2021, 42, 237–247. [Google Scholar] [CrossRef]
- Tittlemier, S.A.; Brunkhorst, J.; Cramer, B.; DeRosa, M.C.; Lattanzio, V.M.T.; Malone, R.; Maragos, C.; Stranska, M.; Sumarah, M.W. Developments in mycotoxin analysis: An update for 2019-2020. World Mycotoxin J. 2021, 14, 3–26. [Google Scholar] [CrossRef]
- Chen, X.; Wu, H.; Tang, X.; Zhang, Z.; Li, P. Recent Advances in Electrochemical Sensors for Mycotoxin Detection in Food. Electroanalysis 2021, 33, 1–11. [Google Scholar] [CrossRef]
- Hou, Y.; Jia, B.; Sheng, P.; Liao, X.; Shi, L.; Fang, L.; Zhou, L.; Kong, W. Aptasensors for mycotoxins in foods: Recent advances and future trends. Compr. Rev. Food Sci. Food Saf. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Liao, X.; Fang, L.; Jia, B.; Liu, M.; Li, D.; Zhou, L.; Kong, W. Recent advances on immunosensors for mycotoxins in foods and other commodities. TrAC - Trends Anal. Chem. 2021, 136, 116193. [Google Scholar] [CrossRef]
- Nolan, P.; Auer, S.; Spehar, A.; Elliott, C.T.; Campbell, K. Current trends in rapid tests for mycotoxins. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2019, 36, 800–814. [Google Scholar] [CrossRef] [Green Version]
- Peltomaa, R.; Benito-Peña, E.; Moreno-Bondi, M.C. Bioinspired recognition elements for mycotoxin sensors. Anal. Bioanal. Chem. 2018, 410, 747–771. [Google Scholar] [CrossRef]
- Li, R.; Wen, Y.; Wang, F.; He, P. Recent advances in immunoassays and biosensors for mycotoxins detection in feedstuffs and foods. J. Anim. Sci. Biotechnol. 2021, 12, 108. [Google Scholar] [CrossRef]
- Singh, J.; Mehta, A. Rapid and sensitive detection of mycotoxins by advanced and emerging analytical methods: A review. Food Sci. Nutr. 2020, 8, 2183–2204. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Xu, Y. Antibody Developments and Immunoassays for Mycotoxins. Curr. Org. Chem. 2018, 21, 2622–2631. [Google Scholar] [CrossRef]
- Tschofen, M.; Knopp, D.; Hood, E.; Stöger, E. Plant Molecular Farming: Much More than Medicines. Annu. Rev. Anal. Chem. 2016, 9, 271–294. [Google Scholar] [CrossRef]
- Melnik, S.; Neumann, A.C.; Karongo, R.; Dirndorfer, S.; Stübler, M.; Ibl, V.; Niessner, R.; Knopp, D.; Stöger, E. Cloning and plant-based production of antibody MC10E7 for a lateral flow immunoassay to detect [4-arginine]microcystin in freshwater. Plant Biotechnol. J. 2018, 16, 27–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, A.C.; Melnik, S.; Niessner, R.; Stöger, E.; Knopp, D. Microcystin-LR enrichment from freshwater by a recombinant plant-derived antibody using Sol-Gel-Glass immunoextraction. Anal. Sci. 2019, 35, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Ning, Y.; Hu, J.; Lu, F. Aptamers used for biosensors and targeted therapy. Biomed. Pharmacother. 2020, 132, 110902. [Google Scholar] [CrossRef]
- Thyparambil, A.; Bazin, I.; Guiseppi-Elie, A. Molecular Modeling and Simulation Tools in the Development of Peptide-Based Biosensors for Mycotoxin Detection: Example of Ochratoxin. Toxins 2017, 9, 395. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.I.; Shaikh, H.; Mustafa, G.; Bhanger, M.I. Recent Applications of Molecularly Imprinted Polymers in Analytical Chemistry. Sep. Purif. Rev. 2019, 48, 179–219. [Google Scholar] [CrossRef]
- De Middeleer, G.; Dubruel, P.; De Saeger, S. Molecularly imprinted polymers immobilized on 3D printed scaffolds as novel solid phase extraction sorbent for metergoline. Anal. Chim. Acta 2017, 986, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Hatamabadi, D.; Mostafiz, B.; Beirami, A.D.; Banan, K.; Moghaddam, N.S.T.; Afsharara, H.; Keçili, R.; Ghorbani-Bidkorbeh, F. Are molecularly imprinted polymers (Mips) beneficial in detection and determination of mycotoxins in cereal samples? Iran. J. Pharm. Res. 2020, 19, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Langone, J.J.; Vunakis, H. Van Aflatoxin B: Specific Antibodies and Their Use in Radioimmunoassay. J. Natl. Cancer Inst. 1976, 56, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Aalund, O.; Brunfeldt, K.; Hald, B.; Krogh, P.; Poulsen, K. A Radioimmunoassay for Ochratoxin A: A Preliminary Investigation. Acta Pathol. Microbiol. Scand. Sect. C Immunol. 1975, 83 C, 390–392. [Google Scholar] [CrossRef]
- Bonel, L.; Vidal, J.C.; Duato, P.; Castillo, J.R. Ochratoxin A nanostructured electrochemical immunosensors based on polyclonal antibodies and gold nanoparticles coupled to the antigen. Anal. Methods 2010, 2, 335. [Google Scholar] [CrossRef]
- Thirumala-Devi, K.; Mayo, M.A.; Reddy, G.; Reddy, S.V.; Delfosse, P.; Reddy, D.V.R. Production of polyclonal antibodies against Ochratoxin A and its detection in chilies by ELISA. J. Agric. Food Chem. 2000, 48, 5079–5082. [Google Scholar] [CrossRef]
- Wang, J.J.; Liu, B.H.; Hsu, Y.T.; Yu, F.Y. Sensitive competitive direct enzyme-linked immunosorbent assay and gold nanoparticle immunochromatographic strip for detecting aflatoxin M1 in milk. Food Control 2011, 22, 964–969. [Google Scholar] [CrossRef]
- Liu, B.H.; Hsu, Y.T.; Lu, C.C.; Yu, F.Y. Detecting aflatoxin B1 in foods and feeds by using sensitive rapid enzyme-linked immunosorbent assay and gold nanoparticle immunochromatographic strip. Food Control 2013, 30, 184–189. [Google Scholar] [CrossRef]
- Köhler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Groopman, J.D.; Trudel, L.J.; Donahue, P.R.; Marshak-Rothstein, A.; Wogan, G.N. High-affinity monoclonal antibodies for aflatoxins and their application to solid-phase immunoassays. Proc. Natl. Acad. Sci. USA 1984, 81, 7728–7731. [Google Scholar] [CrossRef] [Green Version]
- Woychik, N.A.; Hinsdill, R.D.; Chu, F.S. Production and characterization of monoclonal antibodies against aflatoxin M1. Appl. Environ. Microbiol. 1984, 48, 1096–1099. [Google Scholar] [CrossRef] [Green Version]
- Rousseau, D.M.; Candlish, A.A.G.; Slegers, G.A.; Van Peteghem, C.H.; Stimson, W.H.; Smith, J.E. Detection of ochratoxin A in porcine kidneys by a monoclonal antibody-based radioimmunoassay. Appl. Environ. Microbiol. 1987, 53, 514–518. [Google Scholar] [CrossRef] [Green Version]
- Casale, W.L.; Pestka, J.J.; Patrick Hart, L. Enzyme-Linked Immunosorbent Assay Employing Monoclonal Antibody Specific for Deoxynivalenol (Vomitoxin) and Several Analogues. J. Agric. Food Chem. 1988, 36, 663–668. [Google Scholar] [CrossRef]
- Dixon, D.E.; Warner, R.L.; Hart, L.P.; Pestka, J.J.; Ram, B.P. Hybridoma cell line production of a specific monoclonal antibody to the mycotoxins zearalenone and.alpha.-zearalenol. J. Agric. Food Chem. 1987, 35, 122–126. [Google Scholar] [CrossRef]
- Hunter, K.W.; Brimfield, A.A.; Miller, M.; Finkelman, F.D.; Chu, S.F. Preparation and characterization of monoclonal antibodies to the trichothecene mycotoxin T-2. Appl. Environ. Microbiol. 1985, 49, 168–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azcona-Olivera, J.I.; Abouzied, M.M.; Plattner, R.D.; Pestka, J.J. Production of monoclonal antibodies to the mycotoxins fumonisins B1, B2, and B3. J. Agric. Food Chem. 1992, 40, 531–534. [Google Scholar] [CrossRef]
- Peng, D.; Yang, B.; Pan, Y.; Wang, Y.; Chen, D.; Liu, Z.; Yang, W.; Tao, Y.; Yuan, Z. Development and validation of a sensitive monoclonal antibody-based indirect competitive enzyme-linked immunosorbent assay for the determination of the aflatoxin M1 levels in milk. Toxicon 2016, 113, 18–24. [Google Scholar] [CrossRef]
- Zhang, X.; Eremin, S.A.; Wen, K.; Yu, X.; Li, C.; Ke, Y.; Jiang, H.; Shen, J.; Wang, Z. Fluorescence Polarization Immunoassay Based on a New Monoclonal Antibody for the Detection of the Zearalenone Class of Mycotoxins in Maize. J. Agric. Food Chem. 2017, 65, 2240–2247. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.W.; Lu, C.C.; Liu, B.H.; Yu, F.Y. Development of novel monoclonal antibodies-based ultrasensitive enzyme-linked immunosorbent assay and rapid immunochromatographic strip for aflatoxin B1 detection. Food Control 2016, 59, 700–707. [Google Scholar] [CrossRef]
- Zhang, X.; Wen, K.; Wang, Z.; Jiang, H.; Beier, R.C.; Shen, J. An ultra-sensitive monoclonal antibody-based fluorescent microsphere immunochromatographic test strip assay for detecting aflatoxin M1 in milk. Food Control 2016, 60, 588–595. [Google Scholar] [CrossRef]
- Li, W.; Powers, S.; Dai, S.Y. Using commercial immunoassay kits for mycotoxins: “Joys and sorrows”? World Mycotoxin J. 2014, 7, 417–430. [Google Scholar] [CrossRef]
- Peltomaa, R.; Barderas, R.; Benito-Peña, E.; Moreno-Bondi, M.C. Recombinant antibodies and their use for food immunoanalysis. Anal. Bioanal. Chem. 2021, 414, 193–217. [Google Scholar] [CrossRef] [PubMed]
- Arola, H.O.; Tullila, A.; Kiljunen, H.; Campbell, K.; Siitari, H.; Nevanen, T.K. Specific Noncompetitive Immunoassay for HT-2 Mycotoxin Detection. Anal. Chem. 2016, 88, 2446–2452. [Google Scholar] [CrossRef]
- Sompunga, P.; Pruksametanan, N.; Rangnoi, K.; Choowongkomon, K.; Yamabhai, M. Generation of human and rabbit recombinant antibodies for the detection of Zearalenone by phage display antibody technology. Talanta 2019, 201, 397–405. [Google Scholar] [CrossRef]
- Hu, Z.Q.; Li, H.P.; Liu, J.L.; Xue, S.; Gong, A.D.; Zhang, J.B.; Liao, Y.C. Production of a phage-displayed mouse ScFv antibody against fumonisin B1 and molecular docking analysis of their interactions. Biotechnol. Bioprocess Eng. 2016, 21, 134–143. [Google Scholar] [CrossRef]
- Ren, W.; Xu, Y.; Huang, Z.; Li, Y.; Tu, Z.; Zou, L.; He, Q.; Fu, J.; Liu, S.; Hammock, B.D. Single-chain variable fragment antibody-based immunochromatographic strip for rapid detection of fumonisin B1 in maize samples. Food Chem. 2020, 319, 126546. [Google Scholar] [CrossRef]
- Cai, C.; Zhang, Q.; Nidiaye, S.; Yan, H.; Zhang, W.; Tang, X.; Li, P. Development of a specific anti-idiotypic nanobody for monitoring aflatoxin M1 in milk and dairy products. Microchem. J. 2021, 167, 106326. [Google Scholar] [CrossRef]
- Wang, F.; Li, Z.F.; Bin Wan, D.; Vasylieva, N.; Shen, Y.D.; Xu, Z.L.; Yang, J.Y.; Gettemans, J.; Wang, H.; Hammock, B.D.; et al. Enhanced Non-Toxic Immunodetection of Alternaria Mycotoxin Tenuazonic Acid Based on Ferritin-Displayed Anti-Idiotypic Nanobody-Nanoluciferase Multimers. J. Agric. Food Chem. 2021, 69, 4911–4917. [Google Scholar] [CrossRef] [PubMed]
- Unkauf, T.; Miethe, S.; Fühner, V.; Schirrmann, T.; Frenzel, A.; Hust, M. Generation of recombinant antibodies against toxins and viruses by phage display for diagnostics and therapy. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2016; Volume 917, pp. 55–76. [Google Scholar]
- Brichta, J.; Hnilova, M.; Viskovic, T. Generation of hapten-specificrecombinantantibodies: Antibody phage display technology: A review. Vet. Med. 2005, 50, 231–252. [Google Scholar] [CrossRef] [Green Version]
- Ståhl, S.; Uhlén, M. Bacterial surface display: Trends and progress. Trends Biotechnol. 1997, 15, 185–192. [Google Scholar] [CrossRef]
- Löfblom, J. Bacterial display in combinatorial protein engineering. Biotechnol. J. 2011, 6, 1115–1129. [Google Scholar] [CrossRef] [PubMed]
- Min, W.-K.; Kim, S.-G.; Seo, J.-H. Affinity maturation of single-chain variable fragment specific for aflatoxin B1 using yeast surface display. Food Chem. 2015, 188, 604–611. [Google Scholar] [CrossRef]
- Davies, S.L.; James, D.C. Engineering Mammalian Cells for Recombinant Monoclonal Antibody Production. In D. Production of recombinant monoclonal antibodies in mammalian cells; Al-Rubeai, M., Ed.; Springer: New York, NY, USA, 2009; pp. 153–173. [Google Scholar]
- Frenzel, A.; Hust, M.; Schirrmann, T. Expression of recombinant antibodies. Front. Immunol. 2013, 4, 217. [Google Scholar] [CrossRef] [Green Version]
- Schirrmann, T.; Al-Halabi, L.; Dübel, S.; Hust, M. Production systems for recombinant antibodies. Front. Biosci. 2008, 13, 4576–4594. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, P.; Lei, J.; Zhang, Q.; Zhang, W.; Li, C. A simple strategy to obtain ultra-sensitive single-chain fragment variable antibodies for aflatoxin detection. RSC Adv. 2013, 3, 22367–22372. [Google Scholar] [CrossRef]
- Hu, Z.Q.; Li, H.P.; Wu, P.; Li, Y.B.; Zhou, Z.Q.; Zhang, J.B.; Liu, J.L.; Liao, Y.C. An affinity improved single-chain antibody from phage display of a library derived from monoclonal antibodies detects fumonisins by immunoassay. Anal. Chim. Acta 2015, 867, 74–82. [Google Scholar] [CrossRef]
- Chang, H.-J.; Choi, S.-W.; Chun, H.S. Expression of functional single-chain variable domain fragment antibody (scFv) against mycotoxin zearalenone in Pichia pastoris. Biotechnol. Lett. 2008, 30, 1801–1806. [Google Scholar] [CrossRef]
- Chalyan, T.; Potrich, C.; Schreuder, E.; Falke, F.; Pasquardini, L.; Pederzolli, C.; Heideman, R.; Pavesi, L. AFM1 Detection in Milk by Fab’ Functionalized Si3N4 Asymmetric Mach-Zehnder Interferometric Biosensors. Toxins 2019, 11, 409. [Google Scholar] [CrossRef] [Green Version]
- Edupuganti, S.R.; Edupuganti, O.P.; Hearty, S.; O’Kennedy, R. A highly stable, sensitive, regenerable and rapid immunoassay for detecting aflatoxin B1 in corn incorporating covalent AFB1 immobilization and a recombinant Fab antibody. Talanta 2013, 115, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Romanazzo, D.; Ricci, F.; Volpe, G.; Elliott, C.T.; Vesco, S.; Kroeger, K.; Moscone, D.; Stroka, J.; Van Egmond, H.; Vehniäinen, M.; et al. Development of a recombinant Fab-fragment based electrochemical immunosensor for deoxynivalenol detection in food samples. Biosens. Bioelectron. 2010, 25, 2615–2621. [Google Scholar] [CrossRef] [PubMed]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hammers, C.; Songa, E.B.; Bendahman, N.; Hammers, R. Naturally occurring antibodies devoid of light chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef]
- Greenberg, A.S.; Avila, D.; Hughes, M.; Hughes, A.; McKinney, E.C.; Flajnik, M.F. A new antigen receptor gene family that undergoes rearrangement and extensive somatic diversification in sharks. Nature 1995, 374, 168–173. [Google Scholar] [CrossRef]
- Muyldermans, S.; Baral, T.N.; Retamozzo, V.C.; De Baetselier, P.; De Genst, E.; Kinne, J.; Leonhardt, H.; Magez, S.; Nguyen, V.K.; Revets, H.; et al. Camelid immunoglobulins and nanobody technology. Vet. Immunol. Immunop. 2009, 128, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, K.B.; Zabetakis, D.; Goldman, E.R.; Anderson, G.P. Enhanced stabilization of a stable single domain antibody for SEB toxin by random mutagenesis and stringent selection. Protein Eng. Des. Sel. 2014, 27, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Govaert, J.; Pellis, M.; Deschacht, N.; Vincke, C.; Conrath, K.; Muyldermans, S.; Saerens, D. Dual beneficial effect of interloop disulfide bond for single domain antibody fragments. J. Biol. Chem. 2012, 287, 1970–1979. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhang, Q.; Tang, X.; Zhang, W.; Li, P. Development of an anti-idiotypic VHH antibody and toxin-free enzyme immunoassay for ochratoxin A in cereals. Toxins 2019, 11, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Tang, Z.; Duan, Z.; He, Z.; Shu, M.; Wang, X.; Gee, S.J.; Hammock, B.D.; Xu, Y. Nanobody-based enzyme immunoassay for ochratoxin A in cereal with high resistance to matrix interference. Talanta 2017, 164, 154–158. [Google Scholar] [CrossRef] [Green Version]
- He, T.; Wang, Y.; Li, P.; Zhang, Q.; Lei, J.; Zhang, Z.; Ding, X.; Zhou, H.; Zhang, W. Nanobody-based enzyme immunoassay for aflatoxin in agro-products with high tolerance to cosolvent methanol. Anal. Chem. 2014, 86, 8873–8880. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Z.; Wang, F.; Cai, J.; Dong, J.; Zhang, J.; Si, R.; Wang, C.; Wang, Y.; Shen, Y.; et al. Isolation of Bactrian Camel Single Domain Antibody for Parathion and Development of One-Step dc-FEIA Method Using VHH-Alkaline Phosphatase Fusion Protein. Anal. Chem. 2018, 90, 12886–12892. [Google Scholar] [CrossRef]
- Muyldermans, S. Applications of Nanobodies. Annu. Rev. Anim. Biosci. 2021, 9, 401–421. [Google Scholar] [CrossRef] [PubMed]
- Doyle, P.J.; Arbabi-Ghahroudi, M.; Gaudette, N.; Furzer, G.; Savard, M.E.; Gleddie, S.; McLean, M.D.; Mackenzie, C.R.; Hall, J.C. Cloning, expression, and characterization of a single-domain antibody fragment with affinity for 15-acetyl-deoxynivalenol. Mol. Immunol. 2008, 45, 3703–3713. [Google Scholar] [CrossRef]
- Wang, F.; Li, Z.F.; Yang, Y.Y.; Bin Wan, D.; Vasylieva, N.; Zhang, Y.Q.; Cai, J.; Wang, H.; Shen, Y.D.; Xu, Z.L.; et al. Chemiluminescent Enzyme Immunoassay and Bioluminescent Enzyme Immunoassay for Tenuazonic Acid Mycotoxin by Exploitation of Nanobody and Nanobody-Nanoluciferase Fusion. Anal. Chem. 2020, 92, 11935–11942. [Google Scholar] [CrossRef] [PubMed]
- Shriver-Lake, L.C.; Goldman, E.R.; Dean, S.N.; Liu, J.L.; Davis, T.M.; Anderson, G.P. Lipid-tagged single domain antibodies for improved enzyme-linked immunosorbent assays. J. Immunol. Methods 2020, 481–482, 112790. [Google Scholar] [CrossRef]
- Anderson, G.P.; Liu, J.L.; Shriver-Lake, L.C.; Zabetakis, D.; Sugiharto, V.A.; Chen, H.W.; Lee, C.R.; Defang, G.N.; Wu, S.J.L.; Venkateswaran, N.; et al. Oriented Immobilization of Single-Domain Antibodies Using SpyTag/SpyCatcher Yields Improved Limits of Detection. Anal. Chem. 2019, 91, 9424–9429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, P.; Zhang, Q.; Zhang, W.; Huang, Y.; Ding, X.; Jiang, J. Production of ultrasensitive generic monoclonal antibodies against major aflatoxins using a modified two-step screening procedure. Anal. Chim. Acta. 2009, 636, 63–69. [Google Scholar] [CrossRef]
- Soukhtanloo, M.; Talebian, E.; Golchin, M.; Mohammadi, M.; Amirheidari, B. Production and Characterization of Monoclonal Antibodies against Aflatoxin B 1. J. Immunoass. Immunochem. 2014, 35, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Ertekin, Ö.; Pirinçci, Ş.Ş.; Öztürk, S. Monoclonal IgA Antibodies for Aflatoxin Immunoassays. Toxins 2016, 8, 148. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.A.; Wang, S.; Allan, R.D.; Kennedy, I.R. A rapid aflatoxin B1 ELISA: Development and validation with reduced matrix effects for peanuts, corn, pistachio, and Soybeans. J. Agric. Food Chem. 2004, 52, 2746–2755. [Google Scholar] [CrossRef]
- Kolosova, A.Y.; Shim, W.-B.; Yang, Z.-Y.; Eremin, S.A.; Chung, D.-H. Direct competitive ELISA based on a monoclonal antibody for detection of aflatoxin B1. Stabilization of ELISA kit components and application to grain samples. Anal. Bioanal. Chem. 2006, 384, 286–294. [Google Scholar] [CrossRef]
- Wu, Y.; Yu, B.; Cui, P.; Yu, T.; Shi, G.; Shen, Z. Development of a quantum dot–based lateral flow immunoassay with high reaction consistency to total aflatoxins in botanical materials. Anal. Bioanal. Chem. 2021, 413, 1629–1637. [Google Scholar] [CrossRef]
- Devi, K.T.; Mayo, M.A.; Reddy, K.L.N.; Delfosse, P.; Reddy, G.; Reddy, S.V.; Reddy, D.V.R. Production and characterization of monoclonal antibodies for aflatoxin B1. Lett. Appl. Microbiol. 1999, 29, 284–288. [Google Scholar] [CrossRef] [Green Version]
- Cervino, C.; Knopp, D.; Weller, M.; Niessner, R. Novel Aflatoxin Derivatives and Protein Conjugates. Molecules 2007, 12, 641–653. [Google Scholar] [CrossRef]
- Cervino, C.; Weber, E.; Knopp, D.; Niessner, R. Comparison of hybridoma screening methods for the efficient detection of high-affinity hapten-specific monoclonal antibodies. J. Immunol. Methods 2008, 329, 184–193. [Google Scholar] [CrossRef]
- LAU, H.P.; GAUR, P.K.; CHU, F.S. Preparation and characterization of aflatoxin B2a-hemiglutarate and its use for the production of antibody against aflatoxin B1. J. Food Saf. 1980, 3, 1–13. [Google Scholar] [CrossRef]
- Zhang, D.; Li, P.; Yang, Y.; Zhang, Q.; Zhang, W.; Xiao, Z.; Ding, X. A high selective immunochromatographic assay for rapid detection of aflatoxin B1. Talanta 2011, 85, 736–742. [Google Scholar] [CrossRef]
- Gaur, P.K.; Lau, H.P.; Pestka, J.J.; Chu, F.S. Production and characterization of aflatoxin B(2a) antiserum. Appl. Environ. Microbiol. 1981, 41, 478–482. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Li, Y.T.; Jiang, J.Q.; Li, R.F.; Fan, G.Y.; Lv, J.M.; Zhou, Y.; Zhang, W.J.; Wang, Z.L. Development of a direct competitive ELISA kit for detecting deoxynivalenol contamination in wheat. Molecules 2020, 25, 50. [Google Scholar] [CrossRef] [Green Version]
- Kong, D.; Wu, X.; Li, Y.; Liu, L.; Song, S.; Zheng, Q.; Kuang, H.; Xu, C. Ultrasensitive and eco-friendly immunoassays based monoclonal antibody for detection of deoxynivalenol in cereal and feed samples. Food Chem. 2019, 270, 130–137. [Google Scholar] [CrossRef]
- Lee, H.M.; Song, S.O.; Cha, S.H.; Wee, S.B.; Bischoff, K.; Park, S.W.; Son, S.W.; Kang, H.G.; Cho, M.H. Development of a monoclonal antibody against deoxynivalenol for magnetic nanoparticle-based extraction and an enzyme-linked immunosorbent assay. J. Vet. Sci. 2013, 14, 143–150. [Google Scholar] [CrossRef]
- Peng, D.; Chang, F.; Wang, Y.; Chen, D.; Liu, Z.; Zhou, X.; Feng, L.; Yuan, Z. Development of a sensitive monoclonal-based enzyme-linked immunosorbent assay for monitoring T-2 toxin in food and feed. Food Addit. Contam. Part A 2016, 33, 683–692. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Peng, D.; Xie, S.; Chen, D.; Pan, Y.; Tao, Y.; Yuan, Z. Construction of electrochemical immunosensor based on gold-nanoparticles/carbon nanotubes/chitosan for sensitive determination of T-2 toxin in feed and swine meat. Int. J. Mol. Sci. 2018, 19, 3895. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Dong, Y.; Guo, J.; Jiang, X.; Liu, J.; Xu, S.; Wang, H. Monoclonal antibody production and the development of an indirect competitive enzyme-linked immunosorbent assay for screening T-2 toxin in milk. Toxicon 2018, 156, 1–6. [Google Scholar] [CrossRef]
- Chu, F.S.; Grossman, S.; Wei, R.D.; Mirocha, C.J. Production of antibody against T-2 toxin. Appl. Environ. Microbiol. 1979, 37, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Luo, X.; Yang, S.; Cao, X.; Wang, Z.; Shi, W.; Zhang, S. High specific monoclonal antibody production and development of an ELISA method for monitoring T-2 toxin in rice. J. Agric. Food Chem. 2014, 62, 1492–1497. [Google Scholar] [CrossRef]
- Cha, S.H.; Kim, S.H.; Bischoff, K.; Kim, H.J.; Son, S.W.; Kang, H.G. Production of a highly group-specific monoclonal antibody against zearalenone and its application in an enzyme-linked immunosorbent assay. J. Vet. Sci. 2012, 13, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.T.; Ram, B.P.; Hart, L.P.; Pestka, J.J. Indirect enzyme-linked immunosorbent assay for the mycotoxin zearalenone. Appl. Environ. Microbiol. 1985, 50, 332–336. [Google Scholar] [CrossRef] [Green Version]
- Pestka, J.J.; Liu, M.-T.; Knudson, B.K.; Hogberg, M.G. Immunization of Swine for Production of Antibody Against Zearalenone. J. Food Prot. 1985, 48, 953–957. [Google Scholar] [CrossRef]
- Thongrussamee, T.; Kuzmina, N.S.; Shim, W.B.; Jiratpong, T.; Eremin, S.A.; Intrasook, J.; Chung, D.H. Monoclonal-based enzyme-linked immunosorbent assay for the detection of zearalenone in cereals. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2008, 25, 997–1006. [Google Scholar] [CrossRef]
- Thouvenot, D.; Morfin, R.F. Radioimmunoassay for zearalenone and zearalanol in human serum: Production, properties, and use of porcine antibodies. Appl. Environ. Microbiol. 1983, 45, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Teshima, R.; Kawase, M.; Tanaka, T.; Sawada, J.I.; Ikebuchi, H.; Terao, T.; Hirai, K.; Sato, M.; Ichinoe, M. Production and Characterization of a Specific Monoclonal Antibody against Mycotoxin Zearalenone. J. Agric. Food Chem. 1990, 38, 1618–1622. [Google Scholar] [CrossRef]
- Gao, Y.; Yang, M.; Peng, C.; Li, X.; Cai, R.; Qi, Y. Preparation of highly specific anti-zearalenone antibodies by using the cationic protein conjugate and development of an indirect competitive enzyme-linked immunosorbent assay. Analyst 2012, 137, 229–236. [Google Scholar] [CrossRef]
- Sun, Y.; Hu, X.; Zhang, Y.; Yang, J.; Wang, F.; Wang, Y.; Deng, R.; Zhang, G. Development of an immunochromatographic strip test for the rapid detection of zearalenone in corn. J. Agric. Food Chem. 2014, 62, 11116–11121. [Google Scholar] [CrossRef]
- Köppen, R.; Riedel, J.; Proske, M.; Drzymala, S.; Rasenko, T.; Durmaz, V.; Weber, M.; Koch, M. Photochemical trans -/ cis -Isomerization and quantitation of zearalenone in edible oils. J. Agric. Food Chem. 2012, 60, 11733–11740. [Google Scholar] [CrossRef]
- Brezina, U.; Kersten, S.; Valenta, H.; Sperfeld, P.; Riedel, J.; Dänicke, S. UV-induced cis-trans isomerization of zearalenone in contaminated maize. Mycotoxin Res. 2013, 29, 221–227. [Google Scholar] [CrossRef]
- Gadzała-Kopciuch, R.; Kuźniewska, A.; Buszewski, B. Analytical approaches and preparation of biological, food and environmental samples for analyses of zearalenone and its metabolites. Rev. Anal. Chem. 2020, 39, 157–167. [Google Scholar] [CrossRef]
- Oswald, S.; Niessner, R.; Knopp, D. First experimental evidence for an enzyme-generated chemiluminescence-induced trans-cis isomerization of chip-immobilized zearalenone in a microfluidic cell of a biosensor. Luminesence 2014, 29, 23–24. [Google Scholar]
- Chu, F.S.; Ueno, I. Production of antibody against aflatoxin B1. Appl. Environ. Microbiol. 1977, 33, 1125. [Google Scholar] [CrossRef] [Green Version]
- Chu, F.S.; Chang, F.C.C.; Hinsdill, R.D. Production of antibody against ochratoxin A. Appl. Environ. Microbiol. 1976, 31, 831–835. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Li, Y.; Jiang, J.; Li, R.; Fan, G.; Lei, Z.; Wang, H.; Wang, Z.; Zhang, W. Preparation and characterisation of monoclonal antibodies against deoxynivalenol. Ital. J. Anim. Sci. 2020, 19, 560–568. [Google Scholar] [CrossRef]
- Yu, F.-Y.; Chu, F.S. Production and Characterization of Antibodies against Fumonisin B1. J. Food Prot. 1996, 59, 992–997. [Google Scholar] [CrossRef]
- Kalayu Yirga, S.; Ling, S.; Yang, Y.; Yuan, J.; Wang, S. The Preparation and Identification of a Monoclonal Antibody against Citrinin and the Development of Detection via Indirect Competitive ELISA. Toxins 2017, 9, 110. [Google Scholar] [CrossRef] [Green Version]
- Mhadhbi, H.; Benrejeb, S.; Martel, A. Studies on the affinity chromatography purification of anti-patulin polyclonal antibodies by enzyme linked immunosorbent assay and electrophoresis. Food Addit. Contam. 2005, 22, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- De Champdoré, M.; Bazzicalupo, P.; De Napoli, L.; Montesarchio, D.; Di Fabio, G.; Cocozza, I.; Parracino, A.; Rossi, M.; D’Auria, S. A new competitive fluorescence assay for the detection of patulin toxin. Anal. Chem. 2007, 79, 751–757. [Google Scholar] [CrossRef]
- McElroy, L.J.; Weiss, C.M. The production of polyclonal antibodies against the mycotoxin derivative patulin hemiglutarate. Can. J. Microbiol. 1993, 39, 861–863. [Google Scholar] [CrossRef]
- Pennacchio, A.; Varriale, A.; Esposito, M.G.; Staiano, M.; D’Auria, S. A near-infrared fluorescence assay method to detect patulin in food. Anal. Biochem. 2015, 481, 55–59. [Google Scholar] [CrossRef]
- Liu, B.H.; Tsao, Z.J.; Wang, J.J.; Yu, F.Y. Development of a monoclonal antibody against ochratoxin A and its application in enzyme-linked immunosorbent assay and gold nanoparticle immunochromatographic strip. Anal. Chem. 2008, 80, 7029–7035. [Google Scholar] [CrossRef]
- Cho, Y.J.; Lee, D.H.; Kim, D.O.; Min, W.K.; Bong, K.T.; Lee, G.G.; Seo, J.H. Production of a monoclonal antibody against ochratoxin A and its application to immunochromatographic assay. J. Agric. Food Chem. 2005, 53, 8447–8451. [Google Scholar] [CrossRef]
- Maragos, C.M.; Li, L.; Chen, D. Production and characterization of a single chain variable fragment (scFv) against the mycotoxin deoxynivalenol. Food Agric. Immunol. 2012, 23, 51–67. [Google Scholar] [CrossRef] [Green Version]
- Min, W.-K.; Cho, Y.-J.; Park, J.-B.; Bae, Y.-H.; Kim, E.-J.; Park, K.; Park, Y.-C.; Seo, J.-H. Production and characterization of monoclonal antibody and its recombinant single chain variable fragment specific for a food-born mycotoxin, fumonisin B1. Bioprocess Biosyst. Eng. 2010, 33, 109–115. [Google Scholar] [CrossRef]
- Min, W.K.; Kweon, D.H.; Park, K.; Park, Y.C.; Seo, J.H. Characterisation of monoclonal antibody against aflatoxin B1 produced in hybridoma 2C12 and its single-chain variable fragment expressed in recombinant Escherichia coli. Food Chem. 2011, 126, 1316–1323. [Google Scholar] [CrossRef]
- Wang, R.; Gu, X.; Zhuang, Z.; Zhong, Y.; Yang, H.; Wang, S. Screening and Molecular Evolution of a Single Chain Variable Fragment Antibody (scFv) against Citreoviridin Toxin. J. Agric. Food Chem. 2016, 64, 7640–7648. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Q.; Sun, Z.; Wang, Y.; Su, B.; Zhang, C.; Cao, H.; Liu, X. Nanobody affinity improvement: Directed evolution of the anti-ochratoxin A single domain antibody. Int. J. Biol. Macromol. 2020, 151, 312–321. [Google Scholar] [CrossRef]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef]
- Ruscito, A.; Smith, M.; Goudreau, D.N.; Derosa, M.C. Current status and future prospects for aptamer-based mycotoxin detection. J. AOAC Int. 2016, 99, 865–877. [Google Scholar] [CrossRef]
- Hasegawa, H.; Savory, N.; Abe, K.; Ikebukuro, K. Methods for Improving Aptamer Binding Affinity. Molecules 2016, 21, 421. [Google Scholar] [CrossRef]
- Cruz-Aguado, J.A.; Penner, G. Determination of Ochratoxin A with a DNA Aptamer. J. Agric. Food Chem. 2008, 56, 10456–10461. [Google Scholar] [CrossRef] [PubMed]
- Shim, W.B.; Kim, M.J.; Mun, H.; Kim, M.G. An aptamer-based dipstick assay for the rapid and simple detection of aflatoxin B1. Biosens. Bioelectron. 2014, 62, 288–294. [Google Scholar] [CrossRef]
- Malhotra, S.; Pandey, A.K.; Rajput, Y.S.; Sharma, R. Selection of aptamers for aflatoxin M1 and their characterization. J. Mol. Recognit. 2014, 27, 493–500. [Google Scholar] [CrossRef]
- Yue, S.; Jie, X.; Wei, L.; Bin, C.; Wang, D.; Yi, Y.; Lin, Q.; Li, J.; Zheng, T. Simultaneous detection of ochratoxin a and fumonisin b1 in cereal samples using an aptamer-photonic crystal encoded suspension array. Anal. Chem. 2014, 86, 11797–11802. [Google Scholar] [CrossRef]
- Chryseis Le, L.; Cruz-Aguado, J.; Penner, G.A. DNA Ligands for Aflatoxins and Zearalenone. World Patent Application Application PCT/CA2010/001292, 20 August 2010. [Google Scholar]
- Ong, C.C.; Siva Sangu, S.; Illias, N.M.; Chandra Bose Gopinath, S.; Saheed, M.S.M. Iron nanoflorets on 3D-graphene-nickel: A ‘Dandelion’ nanostructure for selective deoxynivalenol detection. Biosens. Bioelectron. 2020, 154, 112088. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Huang, Q.; Nie, D.; Zhao, X.; Cao, H.; Wu, W.; Han, Z. A multifunctional n-doped cu–mofs (N–cu–mof) nanomaterial-driven electrochemical aptasensor for sensitive detection of deoxynivalenol. Molecules 2021, 26, 2243. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Duan, N.; Zhang, W.; Zhao, S.; Wang, Z. Screening and development of DNA aptamers as capture probes for colorimetric detection of patulin. Anal. Biochem. 2016, 508, 58–64. [Google Scholar] [CrossRef]
- Chen, X.; Huang, Y.; Duan, N.; Wu, S.; Xia, Y.; Ma, X.; Zhu, C.; Jiang, Y.; Wang, Z. Screening and identification of DNA aptamers against T-2 toxin assisted by graphene oxide. J. Agric. Food Chem. 2014, 62, 10368–10374. [Google Scholar] [CrossRef] [PubMed]
- Rouah-Martin, E.; Mehta, J.; van Dorst, B.; de Saeger, S.; Dubruel, P.; Maes, B.U.W.; Lemiere, F.; Goormaghtigh, E.; Daems, D.; Herrebout, W.; et al. Aptamer-based molecular recognition of lysergamine, metergoline and small ergot alkaloids. Int. J. Mol. Sci. 2012, 13, 17138–17159. [Google Scholar] [CrossRef]
- Nguyen, B.H.; Tran, L.D.; Do, Q.P.; Le Nguyen, H.; Tran, N.H.; Nguyen, P.X. Label-free detection of aflatoxin M1 with electrochemical Fe 3O4/polyaniline-based aptasensor. Mater. Sci. Eng. C 2013, 33, 2229–2234. [Google Scholar] [CrossRef]
- McKeague, M.; Bradley, C.R.; De Girolamo, A.; Visconti, A.; Miller, J.D.; Derosa, M.C. Screening and initial binding assessment of fumonisin b(1) aptamers. Int. J. Mol. Sci. 2010, 11, 4864–4881. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Huang, Y.; Duan, N.; Wu, S.; Ma, X.; Xia, Y.; Zhu, C.; Jiang, Y.; Wang, Z. Selection and identification of ssDNA aptamers recognizing zearalenone. Anal. Bioanal. Chem. 2013, 405, 6573–6581. [Google Scholar] [CrossRef]
- Penner, G. Commercialization of an aptamer-based diagnostic test. IVD Technol. 2012, 18, 31–37. Available online: https://www.researchgate.net/publication/291178318 (accessed on 12 October 2021).
- Wang, Y.; Wang, H.; Li, P.; Zhang, Q.; Kim, H.J.; Gee, S.J.; Hammock, B.D. Phage-displayed peptide that mimics aflatoxins and its application in immunoassay. J. Agric. Food Chem. 2013, 61, 2426–2433. [Google Scholar] [CrossRef]
- He, Q.; Xu, Y.; Zhang, C.; Li, Y.; Huang, Z. Phage-borne peptidomimetics as immunochemical reagent in dot-immunoassay for mycotoxin zearalenone. Food Control 2014, 39, 56–61. [Google Scholar] [CrossRef]
- Lai, W.; Fung, D.Y.C.; Yang, X.; Renrong, L.; Xiong, Y. Development of a colloidal gold strip for rapid detection of ochratoxin A with mimotope peptide. Food Control 2009, 20, 791–795. [Google Scholar] [CrossRef]
- He, Q.-H.; Xu, Y.; Huang, Y.-H.; Liu, R.-R.; Huang, Z.-B.; Li, Y.-P. Phage-displayed peptides that mimic zearalenone and its application in immunoassay. Food Chem. 2011, 126, 1312–1315. [Google Scholar] [CrossRef]
- Yuan, Q.; Pestka, J.J.; Hespenheide, B.M.; Kuhn, L.A.; Linz, J.E.; Hart, L.P. Identification of mimotope peptides which bind to the mycotoxin deoxynivalenol-specific monoclonal antibody. Appl. Environ. Microbiol. 1999, 65, 3279–3286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Yu, Z.; He, Q.; Xu, Y. An immunoassay for ochratoxin A without the mycotoxin. Food Control 2007, 18, 872–877. [Google Scholar] [CrossRef]
- Thirumala-Devi, K.; Miller, J.S.; Reddy, G.; Reddy, D.V.R.; Mayo, M.A. Phage-displayed peptides that mimic aflatoxin B1 in serological reactivity. J. Appl. Microbiol. 2001, 90, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Tozzi, C.; Anfossi, L.; Baggiani, C.; Giovannoli, C.; Giraudi, G. A combinatorial approach to obtain affinity media with binding properties towards the aflatoxins. Anal. Bioanal. Chem. 2003, 375, 994–999. [Google Scholar] [CrossRef]
- Heurich, M.; Altintas, Z.; Tothill, I.E. Computational design of peptide ligands for ochratoxin A. Toxins 2013, 5, 1202–1212. [Google Scholar] [CrossRef]
- Giraudi, G.; Anfossi, L.; Baggiani, C.; Giovannoli, C.; Tozzi, C. Solid-phase extraction of ochratoxin A from wine based on a binding hexapeptide prepared by combinatorial synthesis. J. Chromatogr. A 2007, 1175, 174–180. [Google Scholar] [CrossRef]
- Bazin, I.; Andreotti, N.; Hassine, A.I.H.; De Waard, M.; Sabatier, J.M.; Gonzalez, C. Peptide binding to ochratoxin A mycotoxin: A new approach in conception of biosensors. Biosens. Bioelectron. 2013, 40, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Peng, J.; Wu, Q.; Zhao, Y.; Shang, H.; Wang, S. A novel screening on the specific peptide by molecular simulation and development of the electrochemical immunosensor for aflatoxin B1 in grains. Food Chem. 2022, 372, 131322. [Google Scholar] [CrossRef] [PubMed]
- Appell, M.; Maragos, C.M.; Kendra, D.F. Molecularly imprinted polymers for mycotoxins. In Proceedings of the ACS Symposium Series, 32nd ACS National Meeting, San Francisco, CA, USA, 10–14, September 2006; American Chemical Society: Washington, DC, USA, 2008; Volume 1001, pp. 152–169. [Google Scholar]
- Leibl, N.; Haupt, K.; Gonzato, C.; Duma, L. Molecularly Imprinted Polymers for Chemical Sensing: A Tutorial Review. Chemosensors 2021, 9, 123. [Google Scholar] [CrossRef]
- Urraca, J.L.; Marazuela, M.D.; Moreno-Bondi, M.C. Molecularly imprinted polymers applied to the clean-up of zearalenone and α-zearalenol from cereal and swine feed sample extracts. Anal. Bioanal. Chem. 2006, 385, 1155–1161. [Google Scholar] [CrossRef]
- Urraca, J.L.; Marazuela, M.D.; Merino, E.R.; Orellana, G.; Moreno-Bondi, M.C. Molecularly imprinted polymers with a streamlined mimic for zearalenone analysis. J. Chromatogr. A 2006, 1116, 127–134. [Google Scholar] [CrossRef]
- Baggiani, C.; Giraudi, G.; Vanni, A. A molecular imprinted polymer with recognition properties towards the carcinogenic mycotoxin ochratoxin a. Bioseparation 2001, 10, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.C.C.; Lai, E.P.C. Molecularly Imprinted Polymers for Ochratoxin A Extraction and Analysis. Toxins 2010, 2, 1536–1553. [Google Scholar] [CrossRef] [PubMed]
- Sergeyeva, T.A.; Piletska, O.V.; Brovko, O.O.; Goncharova, L.A.; Piletsky, S.A.; El’ska, G.V. Aflatoxin-selective molecularly-imprinted polymer membranes based on acrylate-polyurethane semi-interpenetrating polymer networks. Ukr. Biokhimichnyi Zhurnal 2007, 79, 109–115. [Google Scholar]
- Sergeyeva, T.; Yarynka, D.; Piletska, E.; Lynnik, R.; Zaporozhets, O.; Brovko, O.; Piletsky, S.; El’skaya, A. Fluorescent sensor systems based on nanostructured polymeric membranes for selective recognition of Aflatoxin B1. Talanta 2017, 175, 101–107. [Google Scholar] [CrossRef]
- Munawar, H.; Smolinska-Kempisty, K.; Cruz, A.G.; Canfarotta, F.; Piletska, E.; Karim, K.; Piletsky, S.A. Molecularly imprinted polymer nanoparticle-based assay (MINA): Application for fumonisin B1 determination. Analyst 2018, 143, 3481–3488. [Google Scholar] [CrossRef]
- Maier, N.M.; Buttinger, G.; Welhartizki, S.; Gavioli, E.; Lindner, W. Molecularly imprinted polymer-assisted sample clean-up of ochratoxin a from red wine: Merits and limitations. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2004, 804, 103–111. [Google Scholar] [CrossRef]
- Baggiani, C.; Giovannoli, C.; Anfossi, L.; Passini, C.; Baravalle, P.; Giraudi, G. A Connection between the binding properties of imprinted and nonimprinted polymers: A change of perspective in molecular imprinting. J. Am. Chem. Soc. 2012, 134, 1513–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascale, M.; De Girolamo, A.; Visconti, A.; Magan, N.; Chianella, I.; Piletska, E.V.; Piletsky, S.A. Use of itaconic acid-based polymers for solid-phase extraction of deoxynivalenol and application to pasta analysis. Anal. Chim. Acta 2008, 609, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Piletska, E.; Karim, K.; Coker, R.; Piletsky, S. Development of the custom polymeric materials specific for aflatoxin B1 and ochratoxin A for application with the ToxiQuant T1 sensor tool. J. Chromatogr. A 2010, 1217, 2543–2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piletsky, S.A.; Piletska, E.V.; Karim, K.; Freebairn, K.W.; Legge, C.H.; Turner, A.P.F. Polymer cookery: Influence of polymerization conditions on the performance of molecularly imprinted polymers. Macromolecules 2002, 35, 7499–7504. [Google Scholar] [CrossRef]
- Turner, N.W.; Piletska, E.V.; Karim, K.; Whitcombe, M.; Malecha, M.; Magan, N.; Baggiani, C.; Piletsky, S.A. Effect of the solvent on recognition properties of molecularly imprinted polymer specific for ochratoxin A. Biosens. Bioelectron. 2004, 20, 1060–1067. [Google Scholar] [CrossRef]
- Navarro-Villoslada, F.; Vicente, B.S.; Moreno-Bondi, M.C. Application of multivariate analysis to the screening of molecularly imprinted polymers for bisphenol A. Anal. Chim. Acta 2004, 504, 149–162. [Google Scholar] [CrossRef]
- Weiss, R.; Freudenschuss, M.; Krska, R.; Mizaikoff, B. Improving methods of analysis for mycotoxins: Molecularly imprinted polymers for deoxynivalenol and zearalenone. Food Addit. Contam. 2003, 20, 386–395. [Google Scholar] [CrossRef]
- Díaz-Bao, M.; Regal, P.; Barreiro, R.; Fente, C.A.; Cepeda, A. A facile method for the fabrication of magnetic molecularly imprinted stir-bars: A practical example with aflatoxins in baby foods. J. Chromatogr. A 2016, 1471, 51–59. [Google Scholar] [CrossRef]
- Liang, Y.; He, J.; Huang, Z.; Li, H.; Zhang, Y.; Wang, H.; Rui, C.; Li, Y.; You, L.; Li, K.; et al. An amino-functionalized zirconium-based metal-organic framework of type UiO-66-NH2 covered with a molecularly imprinted polymer as a sorbent for the extraction of aflatoxins AFB1, AFB2, AFG1 and AFG2 from grain. Microchim. Acta 2020, 187, 32. [Google Scholar] [CrossRef]
- Giovannoli, C.; Passini, C.; Di Nardo, F.; Anfossi, L.; Baggiani, C. Determination of ochratoxin A in Italian red wines by molecularly imprinted solid phase extraction and HPLC analysis. J. Agric. Food Chem. 2014, 62, 5220–5225. [Google Scholar] [CrossRef]
- De Smet, D.; Dubruel, P.; Van Peteghem, C.; Schacht, E.; De Saeger, S. Molecularly imprinted solid-phase extraction of fumonisin B analogues in bell pepper, rice and corn flakes. Food Addit. Contam. Part A 2009, 26, 874–884. [Google Scholar] [CrossRef]
- Urraca, J.L.; Huertas-Pérez, J.F.; Cazorla, G.A.; Gracia-Mora, J.; García-Campaña, A.M.; Moreno-Bondi, M.C. Development of magnetic molecularly imprinted polymers for selective extraction: Determination of citrinin in rice samples by liquid chromatography with UV diode array detection. Anal. Bioanal. Chem. 2016, 408, 3033–3042. [Google Scholar] [CrossRef]
- De Smet, D.; Monbaliu, S.; Dubruel, P.; Van Peteghem, C.; Schacht, E.; De Saeger, S. Synthesis and application of a T-2 toxin imprinted polymer. J. Chromatogr. A 2010, 1217, 2879–2886. [Google Scholar] [CrossRef]
- Regal, P.; Díaz-Bao, M.; Barreiro, R.; Fente, C.; Cepeda, A. Design of a molecularly imprinted stir-bar for isolation of patulin in apple and LC-MS/MS detection. Separations 2017, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Lenain, P.; Diana Di Mavungu, J.; Dubruel, P.; Robbens, J.; De Saeger, S. Development of suspension polymerized molecularly imprinted beads with metergoline as template and application in a solid-phase extraction procedure toward ergot alkaloids. Anal. Chem. 2012, 84, 10411–10418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou-Hany, R.A.G.; Urraca, J.L.; Descalzo, A.B.; Gómez-Arribas, L.N.; Moreno-Bondi, M.C.; Orellana, G. Tailoring molecularly imprinted polymer beads for alternariol recognition and analysis by a screening with mycotoxin surrogates. J. Chromatogr. A 2015, 1425, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Ali, W.H.; Derrien, D.; Alix, F.; Pérollier, C.; Lépine, O.; Bayoudh, S.; Chapuis-Hugon, F.; Pichon, V. Solid-phase extraction using molecularly imprinted polymers for selective extraction of a mycotoxin in cereals. J. Chromatogr. A 2010, 1217, 6668–6673. [Google Scholar] [CrossRef] [PubMed]
- Lucci, P.; Derrien, D.; Alix, F.; Pérollier, C.; Bayoudh, S. Molecularly imprinted polymer solid-phase extraction for detection of zearalenone in cereal sample extracts. Anal. Chim. Acta 2010, 672, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Pichon, V.; Combès, A. Selective tools for the solid-phase extraction of Ochratoxin A from various complex samples: Immunosorbents, oligosorbents, and molecularly imprinted polymers. Anal. Bioanal. Chem. 2016, 408, 6983–6999. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Kong, W.; Zhou, S.; Yin, L.; Wan, L.; Yang, M. Molecularly imprinted polymer-based solid phase clean-up for analysis of ochratoxin A in beer, red wine, and grape juice. J. Sep. Sci. 2013, 36, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Bodbodak, S.; Hesari, J.; Peighambardoust, S.H.; Mahkam, M. Selective decontamination of aflatoxin M1 in milk by molecularly imprinted polymer coated on the surface of stainless steel plate. Int. J. Dairy Technol. 2018, 71, 868–878. [Google Scholar] [CrossRef]
- Sun, J.; Guo, W.; Ji, J.; Li, Z.; Yuan, X.; Pi, F.; Zhang, Y.; Sun, X. Removal of patulin in apple juice based on novel magnetic molecularly imprinted adsorbent Fe3O4@SiO2@CS-GO@MIP. LWT-Food Sci. Technol. 2020, 118, 108854. [Google Scholar] [CrossRef]
- Jedziniak, P.; Panasiuk, Ł.; Pietruszka, K.; Posyniak, A. Multiple mycotoxins analysis in animal feed with LC-MS/MS: Comparison of extract dilution and immunoaffinity clean-up. J. Sep. Sci. 2019, 42, 1240–1247. [Google Scholar] [CrossRef]
- Jinap, S.; De Rijk, T.C.; Arzandeh, S.; Kleijnen, H.C.H.; Zomer, P.; Van der Weg, G.; Mol, J.G.J. Aflatoxin determination using in-line immunoaffinity chromatography in foods. Food Control 2012, 26, 42–48. [Google Scholar] [CrossRef]
- Uchigashima, M.; Saigusa, M.; Yamashita, H.; Miyake, S.; Fujita, K.; Nakajima, M.; Nishijima, M. Development of a Novel Immunoaffinity Column for Aflatoxin Analysis Using an Organic Solvent-Tolerant Monoclonal Antibody. J. Agric. Food Chem. 2009, 57, 8728–8734. [Google Scholar] [CrossRef]
- Wilcox, J.; Pazdanska, M.; Milligan, C.; Chan, D.; Macdonald, S.J.; Donnelly, C. Analysis of aflatoxins and ochratoxin a in cannabis and cannabis products by LC–fluorescence detection using cleanup with either multiantibody immunoaffinity columns or an automated system with in-line reusable immunoaffinity cartridges. J. AOAC Int. 2021, 103, 494–503. [Google Scholar] [CrossRef] [PubMed]
- De Girolamo, A.; McKeague, M.; Miller, J.D.; DeRosa, M.C.; Visconti, A. Determination of ochratoxin A in wheat after clean-up through a DNA aptamer-based solid phase extraction column. Food Chem. 2011, 127, 1378–1384. [Google Scholar] [CrossRef] [PubMed]
- Pichon, V.; Brothier, F.; Combès, A. Aptamer-based-sorbents for sample treatment - A review. Anal. Bioanal. Chem. 2015, 407, 681–698. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ma, Y.; Zhang, X.; Yang, X.; Hu, X. A dispersive solid phase extraction adsorbent based on aptamer modified chitosan nanofibers for zearalenone separation in corn, wheat, and beer samples. Anal. Methods 2020, 12, 5852–5860. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Niu, R.; Yang, Y.; Zhou, X.; Luo, S.; Zhang, C.; Wang, Y. Aptamer-functionalized chitosan magnetic nanoparticles as a novel adsorbent for selective extraction of ochratoxin A. Int. J. Biol. Macromol. 2020, 153, 583–590. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, Y.; Zhi, Y.; Wang, X.; Wu, Y.; Zheng, Y. Aptamer-modified magnetic metal-organic framework MIL-101 for highly efficient and selective enrichment of ochratoxin A. J. Sep. Sci. 2018, 42, 716–724. [Google Scholar] [CrossRef]
- Jodlbauer, J.; Maier, N.M.; Lindner, W. Towards ochratoxin A selective molecularly imprinted polymers for solid-phase extraction. J. Chromatogr. A 2002, 945, 45–63. [Google Scholar] [CrossRef]
- Jayasinghe, G.D.T.M.; Domínguez-González, R.; Bermejo-Barrera, P.; Moreda-Piñeiro, A. Ultrasound assisted combined molecularly imprinted polymer for the selective micro-solid phase extraction and determination of aflatoxins in fish feed using liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2020, 1609, 460431. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Shao, H.; Ma, J.; Li, H.; He, Y.; Wang, M.; Jin, F.; Wang, J.; Abd El-Aty, A.M.; Hacımüftüoğlu, A.; et al. Preparation of core-shell magnetic molecularly imprinted polymers for extraction of patulin from juice samples. J. Chromatogr. A 2020, 1615, 460751. [Google Scholar] [CrossRef]
- Turner, N.W.; Bramhmbhatt, H.; Szabo-Vezse, M.; Poma, A.; Coker, R.; Piletsky, S.A. Analytical methods for determination of mycotoxins: An update (2009-2014). Anal. Chim. Acta 2015, 901, 12–33. [Google Scholar] [CrossRef]
- Zhang, K.; Banerjee, K. A Review: Sample Preparation and Chromatographic Technologies for Detection of Aflatoxins in Foods. Toxins 2020, 12, 539. [Google Scholar] [CrossRef] [PubMed]
- Delaunay, N.; Combès, A.; Pichon, V. Immunoaffinity Extraction and Alternative Approaches for the Analysis of Toxins in Environmental, Food or Biological Matrices. Toxins 2020, 12, 795. [Google Scholar] [CrossRef]
- Li, X.; Li, P.; Zhang, Q.; Zhang, Z.; Li, R.; Zhang, W.; Ding, X.; Chen, X.; Tang, X. A Sensitive Immunoaffinity Column-Linked Indirect Competitive ELISA for Ochratoxin A in Cereal and Oil Products Based on a New Monoclonal Antibody. Food Anal. Methods 2013, 6, 1433–1440. [Google Scholar] [CrossRef]
- Hojo, E.; Matsuura, N.; Kamiya, K.; Yonekita, T.; Morishita, N.; Murakami, H.; Kawamura, O. Development of a rapid and versatile method of enzyme-linked immunoassay combined with immunoaffinity column for aflatoxin analysis. J. Food Prot. 2019, 82, 1472–1478. [Google Scholar] [CrossRef]
- Li, D.; Ying, Y.; Wu, J.; Niessner, R.; Knopp, D. Comparison of monomeric and polymeric horseradish peroxidase as labels in competitive ELISA for small molecule detection. Microchim. Acta 2013, 180, 711–717. [Google Scholar] [CrossRef]
- Lin, Y.; Zhou, Q.; Tang, D.; Niessner, R.; Yang, H.; Knopp, D. Silver Nanolabels-Assisted Ion-Exchange Reaction with CdTe Quantum Dots Mediated Exciton Trapping for Signal-On Photoelectrochemical Immunoassay of Mycotoxins. Anal. Chem. 2016, 88, 7858–7866. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, N.; Wang, H.; Zhao, Q. An immunoassay for ochratoxin A using tetramethylrhodamine-labeled ochratoxin A as a probe based on a binding-induced change in fluorescence intensity. Analyst 2020, 145, 651–655. [Google Scholar] [CrossRef]
- Tang, D.; Zhong, Z.; Niessner, R.; Knopp, D. Multifunctional magnetic bead-based electrochemical immunoassay for the detection of aflatoxin B1 in food. Analyst 2009, 134, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Yu, Y.; Niessner, R.; Miró, M.; Knopp, D. Magnetic bead-based fluorescence immunoassay for aflatoxin B1 in food using biofunctionalized rhodamine B-doped silica nanoparticles. Analyst 2010, 135, 2661–2667. [Google Scholar] [CrossRef]
- Tang, D.; Liu, B.; Niessner, R.; Li, P.; Knopp, D. Target-induced displacement reaction accompanying cargo release from magnetic mesoporous silica nanocontainers for fluorescence immunoassay. Anal. Chem. 2013, 85, 10589–10596. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Niessner, R.; Knopp, D. Magnetic Bead-Based Colorimetric Immunoassay for Aflatoxin B1 Using Gold Nanoparticles. Sensors 2014, 14, 21535–21548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Niessner, R.; Tang, D.; Knopp, D. Nanoparticle-based immunosensors and immunoassays for aflatoxins. Anal. Chim. Acta 2016, 912, 10–23. [Google Scholar] [CrossRef]
- Pastucha, M.; Farka, Z.; Lacina, K.; Mikušová, Z.; Skládal, P. Magnetic nanoparticles for smart electrochemical immunoassays: A review on recent developments. Microchim. Acta 2019, 186, 312. [Google Scholar] [CrossRef]
- Zhu, H.; Quan, Z.; Hou, H.; Cai, Y.; Liu, W.; Liu, Y. A colorimetric immunoassay based on cobalt hydroxide nanocages as oxidase mimics for detection of ochratoxin A. Anal. Chim. Acta 2020, 1132, 101–109. [Google Scholar] [CrossRef]
- Zhan, S.; Zheng, L.; Zhou, Y.; Wu, K.; Duan, H.; Huang, X.; Xiong, Y. A gold growth-based plasmonic ELISA for the sensitive detection of fumonisin B1 in maize. Toxins 2019, 11, 323. [Google Scholar] [CrossRef] [Green Version]
- Zhan, S.; Hu, J.; Li, Y.; Huang, X.; Xiong, Y. Direct competitive ELISA enhanced by dynamic light scattering for the ultrasensitive detection of aflatoxin B1 in corn samples. Food Chem. 2021, 342, 128327. [Google Scholar] [CrossRef] [PubMed]
- Beloglazova, N.V.; Speranskaya, E.S.; Wu, A.; Wang, Z.; Sanders, M.; Goftmana, V.V.; Zhang, D.; Goryacheva, I.Y.; Saeger, S.D. Novel multiplex fluorescent immunoassays based on quantum dot nanolabels for mycotoxins determination. Biosens. Bioelectron. 2014, 62, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Na, K.I.; Kim, S.J.; Choi, D.S.; Min, W.K.; Kim, S.G.; Seo, J.H. Extracellular production of functional single-chain variable fragment against aflatoxin B 1 using Escherichia coli. Lett. Appl. Microbiol. 2019, 68, 241–247. [Google Scholar] [CrossRef]
- Rangnoi, K.; Rüker, F.; Wozniak-Knopp, G.; Cvak, B.; O’Kennedy, R.; Yamabhai, M. Binding Characteristic of Various Antibody Formats against Aflatoxins. ACS Omega 2021, 6, 25258–25268. [Google Scholar] [CrossRef] [PubMed]
- Van Houwelingen, A.; De Saeger, S.; Rusanova, T.; Waalwijk, C.; Beekwilder, J. Generation of recombinant alpaca VHH antibody fragments for the detection of the mycotoxin ochratoxin A. World Mycotoxin J. 2008, 1, 407–417. [Google Scholar] [CrossRef]
- Tullila, A.; Nevanen, T.K. Utilization of Multi-Immunization and Multiple Selection Strategies for Isolation of Hapten-Specific Antibodies from Recombinant Antibody Phage Display Libraries. Int. J. Mol. Sci. 2017, 18, 1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, G.-H.; Lee, D.-H.; Min, W.-K.; Cho, Y.-J.; Kweon, D.-H.; Son, D.-H.; Park, K.; Seo, J.-H. Cloning, expression, and characterization of single-chain variable fragment antibody against mycotoxin deoxynivalenol in recombinant Escherichia coli. Protein Expr. Purif. 2004, 35, 84–92. [Google Scholar] [CrossRef]
- Lauer, B.; Ottleben, I.; Jacobsen, H.J.; Reinard, T. Production of a single-chain variable fragment antibody against fumonisin B1. J. Agric. Food Chem. 2005, 53, 899–904. [Google Scholar] [CrossRef]
- Zhou, H.R.; Pestka, J.J.; Hart, L.P. Molecular cloning and expression of recombinant phage antibody against fumonisin B1. J. Food Prot. 1996, 59, 1208–1212. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Chen, Y.; Yang, Y.; Chen, X.; Guo, X.; Du, A. Characterization of anti-citrinin specific ScFvs selected from non-immunized mouse splenocytes by eukaryotic ribosome display. PLoS ONE 2015, 10, e0131482. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Li, X.; Zhao, J.; Zhu, J.; Luo, Y.; Duan, H.; Ji, P.; Wang, K.; Liu, B.; Wang, X.; et al. Nanobody horseradish peroxidase and -EGFP fusions as reagents to detect porcine parvovirus in the immunoassays. J. Nanobiotechnology 2020, 18, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, Y.; Wang, K.; Lu, Q.; Ji, P.; Liu, B.; Zhu, J.; Liu, Q.; Sun, Y.; Zhang, J.; Zhou, E.M.; et al. Nanobody-horseradish peroxidase fusion protein as an ultrasensitive probe to detect antibodies against Newcastle disease virus in the immunoassay. J. Nanobiotechnology 2019, 17, 35. [Google Scholar] [CrossRef] [Green Version]
- Alsulami, T.; Nath, N.; Flemming, R.; Wang, H.; Zhou, W.; Yu, J.H. Development of a novel homogeneous immunoassay using the engineered luminescent enzyme NanoLuc for the quantification of the mycotoxin fumonisin B1. Biosens. Bioelectron. 2021, 177, 112939. [Google Scholar] [CrossRef] [PubMed]
- Barthelmebs, L.; Jonca, J.; Hayat, A.; Prieto-Simon, B.; Marty, J.L. Enzyme-Linked Aptamer Assays (ELAAs), based on a competition format for a rapid and sensitive detection of Ochratoxin A in wine. Food Control 2011, 22, 737–743. [Google Scholar] [CrossRef]
- Li, Y.; Sun, L.; Zhao, Q. Aptamer-Structure Switch Coupled with Horseradish Peroxidase Labeling on a Microplate for the Sensitive Detection of Small Molecules. Anal. Chem. 2019, 91, 2615–2619. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhou, M.; Wang, Y.; Liu, J. Nanozyme and aptamer- based immunosorbent assay for aflatoxin B1. J. Hazard. Mater. 2020, 399, 123154. [Google Scholar] [CrossRef]
- Sun, L.; Li, Y.; Wang, H.; Zhao, Q. An aptamer assay for aflatoxin B1 detection using Mg2+ mediated free zone capillary electrophoresis coupled with laser induced fluorescence. Talanta 2019, 204, 182–188. [Google Scholar] [CrossRef]
- He, D.; Wu, Z.; Cui, B.; Xu, E. Aptamer and gold nanorod–based fumonisin B1 assay using both fluorometry and SERS. Microchim. Acta 2020, 187, 215. [Google Scholar] [CrossRef]
- Xing, K.Y.; Peng, J.; Shan, S.; Liu, D.F.; Huang, Y.N.; Lai, W.H. Green Enzyme-Linked Immunosorbent Assay Based on the Single-Stranded Binding Protein-Assisted Aptamer for the Detection of Mycotoxin. Anal. Chem. 2020, 92, 8422–8426. [Google Scholar] [CrossRef]
- Li, P.; Deng, S.; Zech Xu, Z. Toxicant substitutes in immunological assays for mycotoxins detection: A mini review. Food Chem. 2021, 344, 128589. [Google Scholar] [CrossRef]
- Huang, D.T.; Fu, H.J.; Huang, J.J.; Luo, L.; Lei, H.T.; Shen, Y.D.; Chen, Z.J.; Wang, H.; Xu, Z.L. Mimotope-Based Immunoassays for the Rapid Analysis of Mycotoxin: A Review. J. Agric. Food Chem. 2021, 69, 11743–11752. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Leng, Y.; Li, X.; Huang, X.; Xiong, Y. Emerging strategies to enhance the sensitivity of competitive ELISA for detection of chemical contaminants in food samples. TrAC-Trends Anal. Chem. 2020, 126, 115861. [Google Scholar] [CrossRef]
- Mukhtar, H.; Ma, L.; Pang, Q.; Zhou, Y.; Wang, X.; Xu, T.; Hammock, B.D.; Wang, J. Cyclic peptide: A safe and effective alternative to synthetic aflatoxin B1-competitive antigens. Anal. Bioanal. Chem. 2019, 411, 3881–3890. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, S.; Hong, Z.; Lin, Y.; Dai, H. A mimotope peptide-based dual-signal readout competitive enzyme-linked immunoassay for non-toxic detection of zearalenone. J. Mater. Chem. B 2019, 7, 6972–6980. [Google Scholar] [CrossRef]
- Peltomaa, R.; Fikacek, S.; Benito-Peña, E.; Barderas, R.; Head, T.; Deo, S.; Daunert, S.; Moreno-Bondi, M.C. Bioluminescent detection of zearalenone using recombinant peptidomimetic Gaussia luciferase fusion protein. Microchim. Acta 2020, 187, 547. [Google Scholar] [CrossRef]
- Lu Hou, S.; Ma, Z.E.; Meng, H.; Xu, Y.; He, Q. hua Ultrasensitive and green electrochemical immunosensor for mycotoxin ochratoxin A based on phage displayed mimotope peptide. Talanta 2019, 194, 919–924. [Google Scholar] [CrossRef] [PubMed]
- Peltomaa, R.; Agudo-Maestro, I.; Más, V.; Barderas, R.; Benito-Peña, E.; Moreno-Bondi, M.C. Development and comparison of mimotope-based immunoassays for the analysis of fumonisin B1. Anal. Bioanal. Chem. 2019, 411, 6801–6811. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Shi, Q.; You, K.; Li, Y.; He, Q. Phage displayed mimotope peptide-based immunosensor for green and ultrasensitive detection of mycotoxin deoxynivalenol. J. Pharm. Biomed. Anal. 2019, 168, 94–101. [Google Scholar] [CrossRef]
- Peltomaa, R.; Farka, Z.; Mickert, M.J.; Brandmeier, J.C.; Pastucha, M.; Hlaváček, A.; Martínez-Orts, M.; Canales, Á.; Skládal, P.; Benito-Peña, E.; et al. Competitive upconversion-linked immunoassay using peptide mimetics for the detection of the mycotoxin zearalenone. Biosens. Bioelectron. 2020, 170, 112683. [Google Scholar] [CrossRef] [PubMed]
- Piletsky, S.A.; Piletska, E.V.; Chen, B.; Karim, K.; Weston, D.; Barrett, G.; Lowe, P.; Turner, A.P.F. Chemical grafting of molecularly imprinted homopolymers to the surface of microplates. Application of artificial adrenergic receptor in enzyme-linked assay for β-agonists determination. Anal. Chem. 2000, 72, 4381–4385. [Google Scholar] [CrossRef]
- Wang, S.; Xu, Z.; Fang, G.; Zhang, Y.; Liu, B.; Zhu, H. Development of a Biomimetic Enzyme-Linked Immunosorbent Assay Method for the Determination of Estrone in Environmental Water using Novel Molecularly Imprinted Films of Controlled Thickness as Artificial Antibodies. J. Agric. Food Chem. 2009, 57, 4528–4534. [Google Scholar] [CrossRef] [PubMed]
- Chianella, I.; Guerreiro, A.; Moczko, E.; Caygill, J.S.; Piletska, E.V.; De Vargas Sansalvador, I.M.P.; Whitcombe, M.J.; Piletsky, S.A. Direct replacement of antibodies with molecularly imprinted polymer nanoparticles in ELISA - Development of a novel assay for vancomycin. Anal. Chem. 2013, 85, 8462–8468. [Google Scholar] [CrossRef] [Green Version]
- Cenci, L.; Piotto, C.; Bettotti, P.; Bossi, A.M. Study on molecularly imprinted nanoparticle modified microplates for pseudo-ELISA assays. Talanta 2018, 178, 772–779. [Google Scholar] [CrossRef]
- Munawar, H.; Safaryan, A.H.M.; De Girolamo, A.; Garcia-Cruz, A.; Marote, P.; Karim, K.; Lippolis, V.; Pascale, M.; Piletsky, S.A. Determination of Fumonisin B1 in maize using molecularly imprinted polymer nanoparticles-based assay. Food Chem. 2019, 298, 125044. [Google Scholar] [CrossRef]
- Jia, B.; Liao, X.; Sun, C.; Fang, L.; Zhou, L.; Kong, W. Development of a quantum dot nanobead-based fluorescent strip immunosensor for on-site detection of aflatoxin B1 in lotus seeds. Food Chem. 2021, 356, 129614. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, Q.; Jin, Y.; Zhang, X.; He, L.; Zhang, W.; Chen, Y. Surface enhanced Raman scattering-based lateral flow immunosensor for sensitive detection of aflatoxin M1 in urine. Anal. Chim. Acta 2020, 1128, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Geballa-Koukoula, A.; Gerssen, A.; Nielen, M.W.F. Direct analysis of lateral flow immunoassays for deoxynivalenol using electrospray ionization mass spectrometry. Anal. Bioanal. Chem. 2020, 412, 7547–7558. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Chen, J.; Chen, X.; Ma, T.; Cai, X.; Duan, H.; Leng, Y.; Huang, X.; Xiong, Y. A novel magneto-gold nanohybrid-enhanced lateral flow immunoassay for ultrasensitive and rapid detection of ochratoxin A in grape juice. Food Chem. 2021, 336, 127710. [Google Scholar] [CrossRef]
- Chen, Y.; Fu, Q.; Xie, J.; Wang, H.; Tang, Y. Development of a high sensitivity quantum dot-based fluorescent quenching lateral flow assay for the detection of zearalenone. Anal. Bioanal. Chem. 2019, 411, 2169–2175. [Google Scholar] [CrossRef]
- Chen, X.; Miao, X.; Ma, T.; Leng, Y.; Hao, L.; Duan, H.; Yuan, J.; Li, Y.; Huang, X.; Xiong, Y. Gold Nanobeads with Enhanced Absorbance for Improved Sensitivity in Competitive Lateral Flow Immunoassays. Foods 2021, 10, 1488. [Google Scholar] [CrossRef]
- Qie, Z.; Yan, W.; Gao, Z.; Meng, W.; Xiao, R.; Wang, S. Ovalbumin antibody-based fluorometric immunochromatographic lateral flow assay using CdSe/ZnS quantum dot beads as label for determination of T-2 toxin. Microchim. Acta 2019, 186, 816. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, L.; Kuang, H.; Xu, C. Visible and eco-friendly immunoassays for the detection of cyclopiazonic acid in maize and rice. J. Food Sci. 2020, 85, 105–113. [Google Scholar] [CrossRef]
- Cai, P.; Wang, R.; Ling, S.; Wang, S. A high sensitive platinum-modified colloidal gold immunoassay for tenuazonic acid detection based on monoclonal IgG. Food Chem. 2021, 360, 130021. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Hu, X.; Sun, Y.; Xing, Y.; Xing, G.; Wang, Y.; Li, Q.; Wang, Y.; Deng, R.; Zhang, G. Broad-spectrum detection of zeranol and its analogues by a colloidal gold-based lateral flow immunochromatographic assay in milk. Food Chem. 2020, 321, 126697. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhang, G.; Fang, B.; Xiong, Q.; Duan, H.; Lai, W. Lateral Flow Immunoassay Based on Polydopamine-Coated Gold Nanoparticles for the Sensitive Detection of Zearalenone in Maize. ACS Appl. Mater. Interfaces 2019, 11, 31283–31290. [Google Scholar] [CrossRef]
- Calabria, D.; Calabretta, M.M.; Zangheri, M.; Marchegiani, E.; Trozzi, I.; Guardigli, M.; Michelini, E.; Di Nardo, F.; Anfossi, L.; Baggiani, C.; et al. Recent Advancements in Enzyme-Based Lateral Flow Immunoassays. Sensors 2021, 21, 3358. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Upadhyay, N.; Nara, S. Recent advancements in lateral flow immunoassays: A journey for toxin detection in food. Crit. Rev. Food Sci. Nutr. 2018, 58, 1715–1734. [Google Scholar] [CrossRef]
- Guo, X.; Yuan, Y.; Liu, J.; Fu, S.; Zhang, J.; Mei, Q.; Zhang, Y. Single-Line Flow Assay Platform Based on Orthogonal Emissive Upconversion Nanoparticles. Anal. Chem. 2021, 93, 3010–3017. [Google Scholar] [CrossRef]
- Tang, D.; Sauceda, J.C.; Lin, Z.; Ott, S.; Basova, E.; Goryacheva, I.; Biselli, S.; Lin, J.; Niessner, R.; Knopp, D. Magnetic nanogold microspheres-based lateral-flow immunodipstick for rapid detection of aflatoxin B2 in food. Biosens. Bioelectron. 2009, 25, 514–518. [Google Scholar] [CrossRef]
- Jin, Y.; Chen, Q.; Luo, S.; He, L.; Fan, R.; Zhang, S.; Yang, C.; Chen, Y. Dual near-infrared fluorescence-based lateral flow immunosensor for the detection of zearalenone and deoxynivalenol in maize. Food Chem. 2021, 336, 127718. [Google Scholar] [CrossRef]
- Charlermroj, R.; Phuengwas, S.; Makornwattana, M.; Sooksimuang, T.; Sahasithiwat, S.; Panchan, W.; Sukbangnop, W.; Elliott, C.T.; Karoonuthaisiri, N. Development of a microarray lateral flow strip test using a luminescent organic compound for multiplex detection of five mycotoxins. Talanta 2021, 233, 122540. [Google Scholar] [CrossRef]
- Li, R.; Meng, C.; Wen, Y.; Fu, W.; He, P. Fluorometric lateral flow immunoassay for simultaneous determination of three mycotoxins (aflatoxin B1, zearalenone and deoxynivalenol) using quantum dot microbeads. Microchim. Acta 2019, 186, 748. [Google Scholar] [CrossRef]
- Di Nardo, F.; Alladio, E.; Baggiani, C.; Cavalera, S.; Giovannoli, C.; Spano, G.; Anfossi, L. Colour-encoded lateral flow immunoassay for the simultaneous detection of aflatoxin B1 and type-B fumonisins in a single Test line. Talanta 2019, 192, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhu, J.; Zhang, Z.; Zhang, Q.; Zhang, W.; Yu, L.; Jiang, J.; Chen, X.; Wang, X.; Li, P. Simultaneous lateral flow immunoassay for multi-class chemical contaminants in maize and peanut with one-stop sample preparation. Toxins 2019, 11, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Tang, S.; Jin, Y.; Yang, C.; He, L.; Wang, J.; Chen, Y. Multiplex SERS-based lateral flow immunosensor for the detection of major mycotoxins in maize utilizing dual Raman labels and triple test lines. J. Hazard. Mater. 2020, 393, 122348. [Google Scholar] [CrossRef]
- Goryacheva, O.A.; Guhrenz, C.; Schneider, K.; Beloglazova, N.V.; Goryacheva, I.Y.; De Saeger, S.; Gaponik, N. Silanized Luminescent Quantum Dots for the Simultaneous Multicolor Lateral Flow Immunoassay of Two Mycotoxins. ACS Appl. Mater. Interfaces 2020, 12, 24575–24584. [Google Scholar] [CrossRef]
- Liu, Z.; Hua, Q.; Wang, J.; Liang, Z.; Li, J.; Wu, J.; Shen, X.; Lei, H.; Li, X. A smartphone-based dual detection mode device integrated with two lateral flow immunoassays for multiplex mycotoxins in cereals. Biosens. Bioelectron. 2020, 158, 112178. [Google Scholar] [CrossRef]
- Tang, X.; Li, P.; Zhang, Q.; Zhang, Z.; Zhang, W.; Jiang, J. Time-Resolved Fluorescence Immunochromatographic Assay Developed Using Two Idiotypic Nanobodies for Rapid, Quantitative, and Simultaneous Detection of Aflatoxin and Zearalenone in Maize and Its Products. Anal. Chem. 2017, 89, 11520–11528. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yang, H.; Huang, Z.; Li, Y.; He, Q.; Tu, Z.; Ji, Y.; Ren, W. A peptide/maltose-binding protein fusion protein used to replace the traditional antigen for immunological detection of deoxynivalenol in food and feed. Food Chem. 2018, 268, 242–248. [Google Scholar] [CrossRef]
- Yan, J.X.; Hu, W.J.; You, K.H.; Ma, Z.E.; Xu, Y.; Li, Y.P.; He, Q.H. Biosynthetic Mycotoxin Conjugate Mimetics-Mediated Green Strategy for Multiplex Mycotoxin Immunochromatographic Assay. J. Agric. Food Chem. 2020, 68, 2193–2200. [Google Scholar] [CrossRef]
- Wang, L.; Chen, W.; Ma, W.; Liu, L.; Ma, W.; Zhao, Y.; Zhu, Y.; Xu, L.; Kuang, H.; Xu, C. Fluorescent strip sensor for rapid determination of toxins. Chem. Commun. 2011, 47, 1574–1576. [Google Scholar] [CrossRef]
- Wang, L.; Ma, W.; Chen, W.; Liu, L.; Ma, W.; Zhu, Y.; Xu, L.; Kuang, H.; Xu, C. An aptamer-based chromatographic strip assay for sensitive toxin semi-quantitative detection. Biosens. Bioelectron. 2011, 26, 3059–3062. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, H.; Zhai, W.; Feng, X.; Fan, X.; Chen, A.; Wang, M. A lateral flow strip based on a truncated aptamer-complementary strand for detection of type-B aflatoxins in nuts and dried figs. Toxins 2020, 12, 136. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Liu, L.; Duan, N.; Li, Q.; Zhou, Y.; Wang, Z. Aptamer-Based Lateral Flow Test Strip for Rapid Detection of Zearalenone in Corn Samples. J. Agric. Food Chem. 2018, 66, 1949–1954. [Google Scholar] [CrossRef]
- Wu, S.; Liu, L.; Duan, N.; Wang, W.; Yu, Q.; Wang, Z. A test strip for ochratoxin A based on the use of aptamer-modified fluorescence upconversion nanoparticles. Microchim. Acta 2018, 185, 497. [Google Scholar] [CrossRef]
- Wei, C.W.; Cheng, J.Y.; Huang, C.T.; Yen, M.H.; Young, T.H. Using a microfluidic device for 1 μl DNA microarray hybridization in 500 s. Nucleic Acids Res. 2005, 33, e78. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Zhang, G.; Huang, Y.; Yang, S.; Ren, S.; Gao, Z.; Chen, A. Dual-competitive lateral flow aptasensor for detection of aflatoxin B1 in food and feedstuffs. J. Hazard. Mater. 2018, 344, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Thévenot, D.R.; Toth, K.; Durst, R.A.; Wilson, G.S. Electrochemical biosensors: Recommended definitions and classification. Biosens. Bioelectron. 2001, 16, 121–131. [Google Scholar] [CrossRef]
- Székács, I.; Adányi, N.; Szendrő, I.; Székács, A. Direct and Competitive Optical Grating Immunosensors for Determination of Fusarium Mycotoxin Zearalenone. Toxins 2021, 13, 43. [Google Scholar] [CrossRef] [PubMed]
- Evtugyn, G.; Porfireva, A.; Kulikova, T.; Hianik, T. Recent Achievements in Electrochemical and Surface Plasmon Resonance Aptasensors for Mycotoxins Detection. Chemosensors 2021, 9, 180. [Google Scholar] [CrossRef]
- Zhang, N.; Liu, B.; Cui, X.; Li, Y.; Tang, J.; Wang, H.; Zhang, D.; Li, Z. Recent advances in aptasensors for mycotoxin detection: On the surface and in the colloid. Talanta 2021, 223, 121729. [Google Scholar] [CrossRef] [PubMed]
- Rico-Yuste, A.; Abouhany, R.; Urraca, J.L.; Descalzo, A.B.; Orellana, G.; Moreno-Bondi, M.C. Eu(III)-Templated molecularly imprinted polymer used as a luminescent sensor for the determination of tenuazonic acid mycotoxin in food samples. Sensors Actuators, B Chem. 2021, 329, 129256. [Google Scholar] [CrossRef]
- Elfadil, D.; Lamaoui, A.; Della Pelle, F.; Amine, A.; Compagnone, D. Molecularly imprinted polymers combined with electrochemical sensors for food contaminants analysis. Molecules 2021, 26, 4607. [Google Scholar] [CrossRef] [PubMed]
- Santana Oliveira, I.; da Silva Junior, A.G.; de Andrade, C.A.S.; Lima Oliveira, M.D. Biosensors for early detection of fungi spoilage and toxigenic and mycotoxins in food. Curr. Opin. Food Sci. 2019, 29, 64–79. [Google Scholar] [CrossRef]
- Mujahid, A.; Mustafa, G.; Dickert, F. Label-Free Bioanalyte Detection from Nanometer to Micrometer Dimensions—Molecular Imprinting and QCMs †. Biosensors 2018, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Tothill, I. Biosensors and nanomaterials and their application for mycotoxin determination. World Mycotoxin J. 2011, 4, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Li, W.; Zhu, C.; Li, Y.; Shen, X.; Li, L.; Yan, X.; You, T. Recent progress on electrochemical biosensing of aflatoxins: A review. TrAC-Trends Anal. Chem. 2020, 133, 115966. [Google Scholar] [CrossRef]
- Zhao, W.W.; Xu, J.J.; Chen, H.Y. Photoelectrochemical Immunoassays. Anal. Chem. 2018, 90, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Evtugyn, G.; Porfireva, A.; Shamagsumova, R.; Hianik, T. Advances in Electrochemical Aptasensors Based on Carbon Nanomaterials. Chemosensors 2020, 8, 96. [Google Scholar] [CrossRef]
- Tang, J.; Huang, Y.; Zhang, C.; Liu, H.; Tang, D. Amplified impedimetric immunosensor based on instant catalyst for sensitive determination of ochratoxin A. Biosens. Bioelectron. 2016, 86, 386–392. [Google Scholar] [CrossRef]
- Zong, C.; Jiang, F.; Wang, X.; Li, P.; Xu, L.; Yang, H. Imaging sensor array coupled with dual-signal amplification strategy for ultrasensitive chemiluminescence immunoassay of multiple mycotoxins. Biosens. Bioelectron. 2021, 177, 112998. [Google Scholar] [CrossRef]
- Lin, Y.; Zhou, Q.; Tang, D.; Niessner, R.; Knopp, D. Signal-On Photoelectrochemical Immunoassay for Aflatoxin B1 Based on Enzymatic Product-Etching MnO2 Nanosheets for Dissociation of Carbon Dots. Anal. Chem. 2017, 89, 5637–5645. [Google Scholar] [CrossRef]
- Tang, X.; Wu, J.; Wu, W.; Zhang, Z.; Zhang, W.; Zhang, Q.; Zhang, W.; Chen, X.; Li, P. Competitive-Type Pressure-Dependent Immunosensor for Highly Sensitive Detection of Diacetoxyscirpenol in Wheat via Monoclonal Antibody. Anal. Chem. 2020, 92, 3563–3571. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Nie, D.; Huang, Q.; Fan, K.; Tang, Z.; Wu, Y.; Han, Z. Thin-layer MoS 2 and thionin composite-based electrochemical sensing platform for rapid and sensitive detection of zearalenone in human biofluids. Biosens. Bioelectron. 2019, 130, 322–329. [Google Scholar] [CrossRef]
- Pei, F.; Feng, S.; Wu, Y.; Lv, X.; Wang, H.; Chen, S.M.; Hao, Q.; Cao, Y.; Lei, W.; Tong, Z. Label-free photoelectrochemical immunosensor for aflatoxin B1 detection based on the Z-scheme heterojunction of g-C3N4/Au/WO3. Biosens. Bioelectron. 2021, 189, 113373. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Liu, X.; Su, B.; Chen, Q.; Cao, H.; Yun, Y.; Xu, Y.; Hammock, B.D. Ultrasensitive and rapid detection of ochratoxin A in agro-products by a nanobody-mediated FRET-based immunosensor. J. Hazard. Mater. 2020, 387, 121678. [Google Scholar] [CrossRef]
- Liu, X.; Wen, Y.; Wang, W.; Zhao, Z.; Han, Y.; Tang, K.; Wang, D. Nanobody-based electrochemical competitive immunosensor for the detection of AFB1 through AFB1-HCR as signal amplifier. Microchim. Acta 2020, 187, 352. [Google Scholar] [CrossRef]
- Badie Bostan, H.; Danesh, N.M.; Karimi, G.; Ramezani, M.; Mousavi Shaegh, S.A.; Youssefi, K.; Charbgoo, F.; Abnous, K.; Taghdisi, S.M. Ultrasensitive detection of ochratoxin A using aptasensors. Biosens. Bioelectron. 2017, 98, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Han, Y.; Wang, S.; Ye, Z.; Wei, L.; Xiao, L. Single-Particle LRET Aptasensor for the Sensitive Detection of Aflatoxin B1 with Upconversion Nanoparticles. Anal. Chem. 2019, 91, 11856–11863. [Google Scholar] [CrossRef] [PubMed]
- Jahangiri–Dehaghani, F.; Zare, H.R.; Shekari, Z. Measurement of aflatoxin M1 in powder and pasteurized milk samples by using a label–free electrochemical aptasensor based on platinum nanoparticles loaded on Fe–based metal–organic frameworks. Food Chem. 2020, 310, 125820. [Google Scholar] [CrossRef]
- Ahmadi, A.; Danesh, N.M.; Ramezani, M.; Alibolandi, M.; Lavaee, P.; Emrani, A.S.; Abnous, K.; Taghdisi, S.M. A rapid and simple ratiometric fluorescent sensor for patulin detection based on a stabilized DNA duplex probe containing less amount of aptamer-involved base pairs. Talanta 2019, 204, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Huang, C.; Shao, L.; Wang, X.; Jiao, Y.; Li, W.; Chen, J.; Xu, X. Magneto-controlled aptasensor for simultaneous detection of ochratoxin A and fumonisin B1 using inductively coupled plasma mass spectrometry with multiple metal nanoparticles as element labels. Anal. Chim. Acta 2020, 1127, 182–189. [Google Scholar] [CrossRef]
- Zhong, H.; Yu, C.; Gao, R.; Chen, J.; Yu, Y.; Geng, Y.; Wen, Y.; He, J. A novel sandwich aptasensor for detecting T-2 toxin based on rGO-TEPA-Au@Pt nanorods with a dual signal amplification strategy. Biosens. Bioelectron. 2019, 144, 111635. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Tian, F.; Zhou, J.; Zhao, Q.; Fu, R.; Jiao, B. Colorimetric aptasensor for ochratoxin A detection based on enzyme-induced gold nanoparticle aggregation. J. Hazard. Mater. 2020, 388, 121758. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, H.; Wu, J.; Han, G.; Liu, Y.; Zou, P. A novel fluorescent aptasensor based on exonuclease-assisted triple recycling amplification for sensitive and label-free detection of aflatoxin B1. J. Hazard. Mater. 2021, 415, 125584. [Google Scholar] [CrossRef]
- Yao, Y.; Wang, H.; Wang, X.; Wang, X.; Li, F. Development of a chemiluminescent aptasensor for ultrasensitive and selective detection of aflatoxin B1 in peanut and milk. Talanta 2019, 201, 52–57. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Y.; Li, J.; Huo, B.; Huang, H.; Bai, J.; Peng, Y.; Li, S.; Han, D.; Ren, S.; et al. A fluorescence aptasensor for the sensitive detection of T-2 toxin based on FRET by adjusting the surface electric potentials of UCNPs and MIL-101. Anal. Chim. Acta 2021, 1160, 338450. [Google Scholar] [CrossRef]
- Wu, Z.; He, D.; Cui, B.; Jin, Z.; Xu, E.; Yuan, C.; Liu, P.; Fang, Y.; Chai, Q. Trimer-based aptasensor for simultaneous determination of multiple mycotoxins using SERS and fluorimetry. Microchim. Acta 2020, 187, 495. [Google Scholar] [CrossRef]
- Hao, L.; Wang, W.; Shen, X.; Wang, S.; Li, Q.; An, F.; Wu, S. A Fluorescent DNA Hydrogel Aptasensor Based on the Self-Assembly of Rolling Circle Amplification Products for Sensitive Detection of Ochratoxin A. J. Agric. Food Chem. 2020, 68, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Abnous, K.; Danesh, N.M.; Ramezani, M.; Alibolandi, M.; Nameghi, M.A.; Zavvar, T.S.; Taghdisi, S.M. A novel colorimetric aptasensor for ultrasensitive detection of aflatoxin M1 based on the combination of CRISPR-Cas12a, rolling circle amplification and catalytic activity of gold nanoparticles. Anal. Chim. Acta 2021, 1165, 338549. [Google Scholar] [CrossRef] [PubMed]
- Ong, J.Y.; Pike, A.; Tan, L.L. Recent Advances in Conventional Methods and Electrochemical Aptasensors for Mycotoxin Detection. Foods 2021, 10, 1437. [Google Scholar] [CrossRef]
- Evtugyn, G.; Hianik, T. Electrochemical immuno- and aptasensors for mycotoxin determination. Chemosensors 2019, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Beitollahi, H.; Tajik, S.; Dourandish, Z.; Zhang, K.; Van Le, Q.; Jang, H.W.; Kim, S.Y.; Shokouhimehr, M. Recent Advances in the Aptamer-Based Electrochemical Biosensors for Detecting Aflatoxin B1 and Its Pertinent Metabolite Aflatoxin M1. Sensors 2020, 20, 3256. [Google Scholar] [CrossRef] [PubMed]
- Goud, K.Y.; Reddy, K.K.; Satyanarayana, M.; Kummari, S.; Gobi, K.V. A review on recent developments in optical and electrochemical aptamer-based assays for mycotoxins using advanced nanomaterials. Microchim. Acta 2020, 187, 29. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, F.; Roshanfekr, H.; Peyman, H. Ultra-sensitive electrochemical aptasensor for label-free detection of Aflatoxin B1 in wheat flour sample using factorial design experiments. Food Chem. 2021, 343, 128436. [Google Scholar] [CrossRef]
- Mu, Z.; Ma, L.; Wang, J.; Zhou, J.; Yuan, Y.; Bai, L. A target-induced amperometic aptasensor for sensitive zearalenone detection by CS@AB-MWCNTs nanocomposite as enhancers. Food Chem. 2021, 340, 128128. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yao, X.; Chen, Y.; Gao, Z.; Zhang, J. Near-Infrared Light-Induced Self-Powered Aptasensing Platform for Aflatoxin B1 Based on Upconversion Nanoparticles-Doped Bi2S3Nanorods. Anal. Chem. 2021, 93, 677–682. [Google Scholar] [CrossRef]
- Guo, X.; Wen, F.; Zheng, N.; Li, S.; Fauconnier, M.L.; Wang, J. A qPCR aptasensor for sensitive detection of aflatoxin M1. Anal. Bioanal. Chem. 2016, 408, 5577–5584. [Google Scholar] [CrossRef] [Green Version]
- Sanzani, S.M.; Reverberi, M.; Fanelli, C.; Ippolito, A. Detection of ochratoxin a using molecular beacons and real-time PCR thermal cycler. Toxins 2015, 7, 812–820. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Cheng, Y.; Zheng, J.; Li, J.; Sun, Y.; Peng, S.; Zhu, Z. Target-engineered photo-responsive DNA strands: A novel signal-on photoelectrochemical biosensing platform for ochratoxin A. Anal. Methods 2019, 11, 5638–5644. [Google Scholar] [CrossRef]
- Jia, F.; Liu, D.; Dong, N.; Li, Y.; Meng, S.; You, T. Interaction between the functionalized probes: The depressed efficiency of dual-amplification strategy on ratiometric electrochemical aptasensor for aflatoxin B1. Biosens. Bioelectron. 2021, 182, 113169. [Google Scholar] [CrossRef]
- He, L.; Shen, Z.; Wang, J.; Zeng, J.; Wang, W.; Wu, H.; Wang, Q.; Gan, N. Simultaneously responsive microfluidic chip aptasensor for determination of kanamycin, aflatoxin M1, and 17β-estradiol based on magnetic tripartite DNA assembly nanostructure probes. Microchim. Acta 2020, 187, 176. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Liu, S.; Wang, H.; Zhang, X.; Song, X.; Yu, J.; Huang, J. Primer remodeling amplification-activated multisite-catalytic hairpin assembly enabling the concurrent formation of Y-shaped DNA nanotorches for the fluorescence assay of ochratoxin A. Analyst 2019, 144, 3389–3397. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, Y.; Sun, X.; Bai, J.; Peng, Y.; Ning, B.; Gao, Z.; Liu, B. Ultrasensitive competitive detection of patulin toxin by using strand displacement amplification and DNA G-quadruplex with aggregation-induced emission. Anal. Chim. Acta 2020, 1106, 161–167. [Google Scholar] [CrossRef]
- Yin, J.; Liu, Y.; Wang, S.; Deng, J.; Lin, X.; Gao, J. Engineering a universal and label-free evaluation method for mycotoxins detection based on strand displacement amplification and G-quadruplex signal amplification. Sensors Actuators, B Chem. 2018, 256, 573–579. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Liu, S.; Sun, W.; Zhang, M.; Jiang, L.; Li, M.; Yu, J.; Huang, J. Toehold-mediated DNA strand displacement-driven super-fast tripedal DNA walker for ultrasensitive and label-free electrochemical detection of ochratoxin A. Anal. Chim. Acta 2021, 1143, 21–30. [Google Scholar] [CrossRef]
- Yin, N.; Yuan, S.; Zhang, M.; Wang, J.; Li, Y.; Peng, Y.; Bai, J.; Ning, B.; Liang, J.; Gao, Z. An aptamer-based fluorometric zearalenone assay using a lighting-up silver nanocluster probe and catalyzed by a hairpin assembly. Microchim. Acta 2019, 186, 765. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Li, J.; Liu, B.; Zhang, D.; Zhang, C.; Guo, Y.; Chu, X.; Wang, W.; Wang, H.; Yan, X.; et al. Signal enhancing strategies in aptasensors for the detection of small molecular contaminants by nanomaterials and nucleic acid amplification. Talanta 2022, 236, 122866. [Google Scholar] [CrossRef]
- Wang, Y.; Song, W.; Zhao, H.; Ma, X.; Yang, S.; Qiao, X.; Sheng, Q.; Yue, T. DNA walker-assisted aptasensor for highly sensitive determination of Ochratoxin A. Biosens. Bioelectron. 2021, 182, 113171. [Google Scholar] [CrossRef] [PubMed]
- Lowdon, J.W.; Diliën, H.; Singla, P.; Peeters, M.; Cleij, T.J.; van Grinsven, B.; Eersels, K. MIPs for commercial application in low-cost sensors and assays – An overview of the current status quo. Sensors Actuators B Chem. 2020, 325, 128973. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Xue, X.; Xu, X.; Wen, W.; Chen, M.M.; Zhang, X.; Wang, S. Heterostructured CuO-g-C3N4 nanocomposites as a highly efficient photocathode for photoelectrochemical aflatoxin B1 sensing. Sensors Actuators B Chem. 2021, 329, 129146. [Google Scholar] [CrossRef]
- Akgönüllü, S.; Armutcu, C.; Denizli, A. Molecularly imprinted polymer film based plasmonic sensors for detection of ochratoxin A in dried fig. Polym. Bull. 2021, 78, 1–9. [Google Scholar] [CrossRef]
- Pacheco, J.G.; Castro, M.; Machado, S.; Barroso, M.F.; Nouws, H.P.A.; Delerue-Matos, C. Molecularly imprinted electrochemical sensor for ochratoxin A detection in food samples. Sensors Actuators B Chem. 2015, 215, 107–112. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, M.; Zhang, H.; Wen, W.; Zhang, X.; Wang, S. Solid-state electrochemiluminescence sensor based on RuSi nanoparticles combined with molecularly imprinted polymer for the determination of ochratoxin A. Sensors Actuators B Chem. 2016, 222, 264–269. [Google Scholar] [CrossRef]
- Li, W.; Diao, K.; Qiu, D.; Zeng, Y.; Tang, K.; Zhu, Y. A highly-sensitive and selective antibody-like sensor based on molecularly imprinted poly (L-arginine) on COOH-MWCNTs for electrochemical recognition and detection of deoxynivalenol. Food Chem. 2021, 350, 129229. [Google Scholar] [CrossRef]
- Radi, A.E.; Eissa, A.; Wahdan, T. Molecularly Imprinted Impedimetric Sensor for Determination of Mycotoxin Zearalenone. Electroanalysis 2020, 32, 1788–1794. [Google Scholar] [CrossRef]
- Mao, L.; Ji, K.; Yao, L.; Xue, X.; Wen, W.; Zhang, X.; Wang, S. Molecularly imprinted photoelectrochemical sensor for fumonisin B 1 based on GO-CdS heterojunction. Biosens. Bioelectron. 2019, 127, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xiong, H.; Chen, M.; Zhang, X.; Wang, S. Surface-enhanced molecularly imprinted electrochemiluminescence sensor based on Ru@SiO2 for ultrasensitive detection of fumonisin B1. Biosens. Bioelectron. 2017, 96, 55–61. [Google Scholar] [CrossRef]
- Hu, X.; Liu, Y.; Xia, Y.; Zhao, F.; Zeng, B. A novel ratiometric electrochemical sensor for the selective detection of citrinin based on molecularly imprinted poly(thionine) on ionic liquid decorated boron and nitrogen co-doped hierarchical porous carbon. Food Chem. 2021, 363, 130385. [Google Scholar] [CrossRef]
- Huang, Q.; Zhao, Z.; Nie, D.; Jiang, K.; Guo, W.; Fan, K.; Zhang, Z.; Meng, J.; Wu, Y.; Han, Z. Molecularly imprinted poly(thionine)-based electrochemical sensing platform for fast and selective ultratrace determination of patulin. Anal. Chem. 2019, 91, 4116–4123. [Google Scholar] [CrossRef] [PubMed]
- Hatamluyi, B.; Rezayi, M.; Beheshti, H.R.; Boroushaki, M.T. Ultra-sensitive molecularly imprinted electrochemical sensor for patulin detection based on a novel assembling strategy using Au@Cu-MOF/N-GQDs. Sensors Actuators B. Chem. 2020, 318, 128219. [Google Scholar] [CrossRef]
- Hu, X.; Xia, Y.; Liu, Y.; Zhao, F.; Zeng, B. Determination of patulin using dual-dummy templates imprinted electrochemical sensor with PtPd decorated N-doped porous carbon for amplification. Microchim. Acta 2021, 188, 148. [Google Scholar] [CrossRef]
- Gupta, G.; Bhaskar, A.S.B.; Tripathi, B.K.; Pandey, P.; Boopathi, M.; Rao, L.V.L.; Singh, B.; Vijayaraghavan, R. Supersensitive detection of T-2 toxin by the in situ synthesized π-conjugated molecularly imprinted nanopatterns. An in situ investigation by surface plasmon resonance combined with electrochemistry. Biosens. Bioelectron. 2011, 26, 2534–2540. [Google Scholar] [CrossRef]
- Selvam, S.P.; Kadam, A.N.; Maiyelvaganan, K.R.; Prakash, M.; Cho, S. Novel SeS2-loaded Co MOF with Au@PANI comprised electroanalytical molecularly imprinted polymer-based disposable sensor for patulin mycotoxin. Biosens. Bioelectron. 2021, 187, 113302. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.N.; Niessner, R.; Knopp, D. Enzyme and Immunoassays. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley: Hoboken, NJ, USA, 2021; pp. 1–36. [Google Scholar]
- Zhang, L.; Zhang, Z.; Tian, Y.; Cui, M.; Huang, B.; Luo, T.; Zhang, S.; Wang, H. Rapid, simultaneous detection of mycotoxins with smartphone recognition-based immune microspheres. Anal. Bioanal. Chem. 2021, 413, 3683–3693. [Google Scholar] [CrossRef]
- Oswald, S.; Karsunke, X.Y.Z.; Dietrich, R.; Märtlbauer, E.; Niessner, R.; Knopp, D. Automated regenerable microarray-based immunoassay for rapid parallel quantification of mycotoxins in cereals. Anal. Bioanal. Chem. 2013, 405, 6405–6415. [Google Scholar] [CrossRef]
- Mahmoudpour, M.; Ezzati Nazhad Dolatabadi, J.; Torbati, M.; Homayouni-Rad, A. Nanomaterials based surface plasmon resonance signal enhancement for detection of environmental pollutions. Biosens. Bioelectron. 2019, 127, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Ren, P.; Huang, L.; Ouyang, Z.; Wang, Z.; Kong, X.; Li, T.; Yin, Y.; Wu, Y.; He, Q. Simultaneous detection of aflatoxin B1, ochratoxin A, zearalenone and deoxynivalenol in corn and wheat using surface plasmon resonance. Food Chem. 2019, 300, 125176. [Google Scholar] [CrossRef]
- Liu, S.; Liu, X.; Pan, Q.; Dai, Z.; Pan, M.; Wang, S. A Portable, Label-Free, Reproducible Quartz Crystal Microbalance Immunochip for the Detection of Zearalenone in Food Samples. Biosensors 2021, 11, 53. [Google Scholar] [CrossRef]
- Susilo, M.D.D.; Jayadi, T.; Kusumaatmaja, A.; Nugraheni, A.D. Development of molecular imprinting polymer nanofiber for aflatoxin B1 detection based on quartz crystal microbalance. In Proceedings of the Materials Science Forum, 9th International Conference on Materials Science and Engineering Technology (ICMSET 2020) held virtually in Kyoto, 9–11 October 2020; Trans Tech Publications Ltd.: Bäch, Switzerland, 2021; Volume 1023, pp. 103–111. [Google Scholar] [CrossRef]
- Ricciardi, C.; Castagna, R.; Ferrante, I.; Frascella, F.; Luigi Marasso, S.; Ricci, A.; Canavese, G.; Lorè, A.; Prelle, A.; Lodovica Gullino, M.; et al. Development of a microcantilever-based immunosensing method for mycotoxin detection. Biosens. Bioelectron. 2013, 40, 233–239. [Google Scholar] [CrossRef]
- Maragos, C.M. Signal amplification using colloidal gold in a biolayer interferometry-based immunosensor for the mycotoxin deoxynivalenol. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2012, 29, 1108–1117. [Google Scholar] [CrossRef] [PubMed]
- Lux, G.; Langer, A.; Pschenitza, M.; Karsunke, X.; Strasser, R.; Niessner, R.; Knopp, D.; Rant, U. Detection of the carcinogenic water pollutant benzo [a]pyrene with an electro-switchable biosurface. Anal. Chem. 2015, 87, 4538–4545. [Google Scholar] [CrossRef] [PubMed]
- Rant, U. Sensing with electro-switchable biosurfaces. Bioanal. Rev. 2012, 4, 97–114. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, J.; Fotina, H.; Zhang, H.; Chen, J. Advances in Antibody Preparation Techniques for Immunoassays of Total Aflatoxin in Food. Molecules 2020, 25, 4113. [Google Scholar] [CrossRef]
- Weller, M.G. Quality Issues of Research Antibodies. Anal. Chem. Insights 2016, 2016, 21–27. [Google Scholar] [CrossRef] [Green Version]
- O’Kennedy, R.; Fitzgerald, S.; Murphy, C. Don’t blame it all on antibodies – The need for exhaustive characterisation, appropriate handling, and addressing the issues that affect specificity. TrAC-Trends Anal. Chem. 2017, 89, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Directive, E. Directive 2010/63/EU of the European Parliament and of the council of 22 September 2010 on the protection of animals used for scientific purposes. Off. J. Eur. Union 2010, 276, 33–79. [Google Scholar]
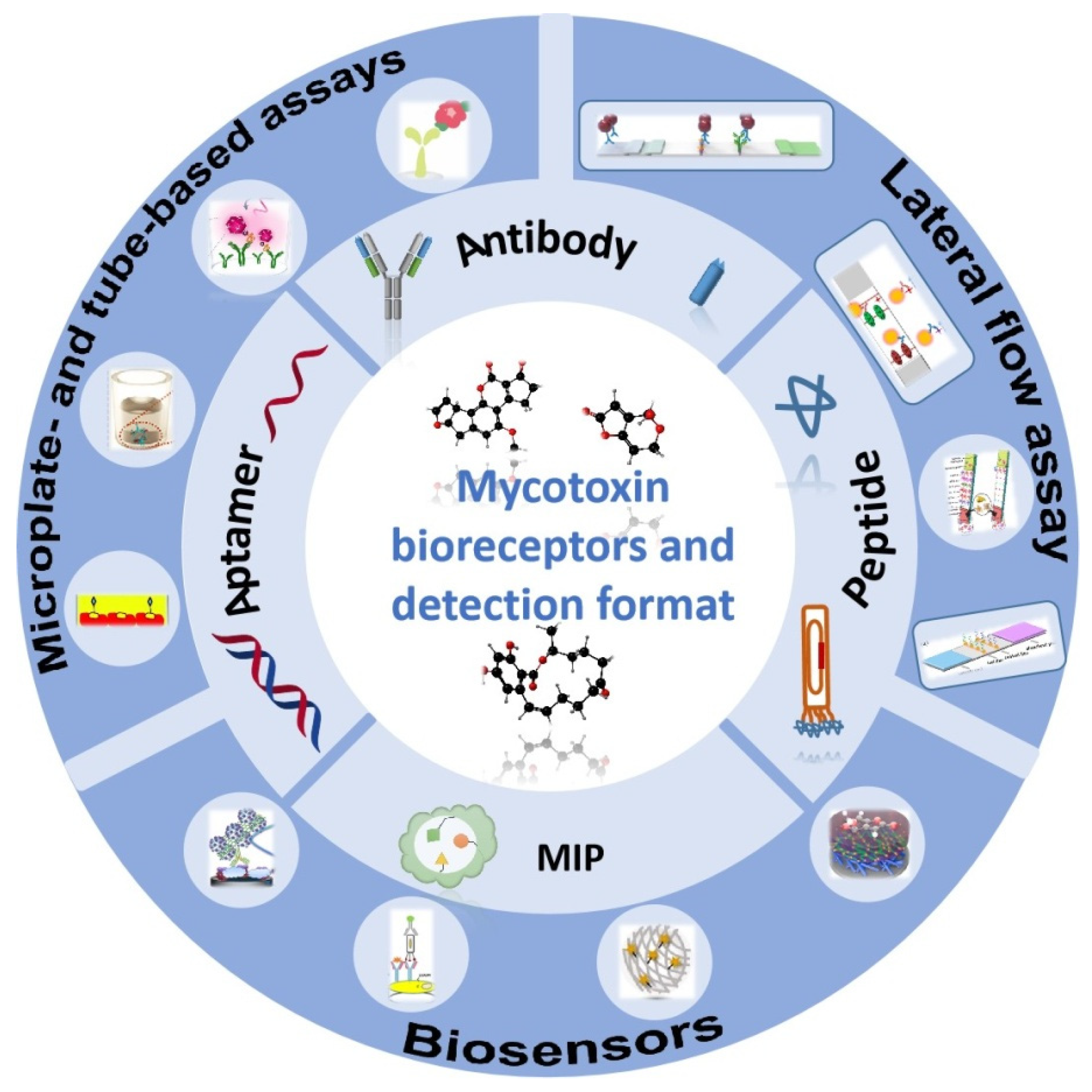
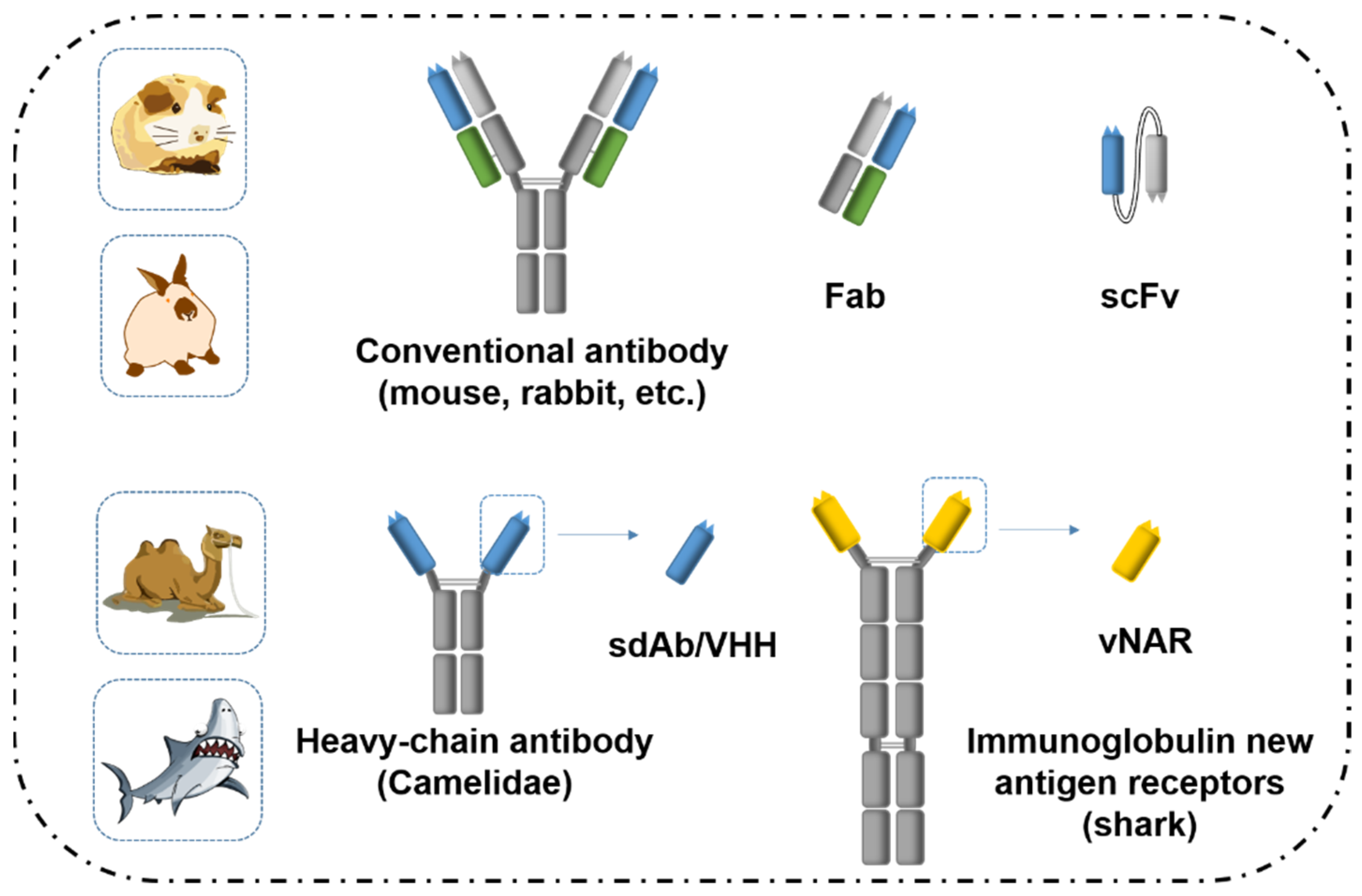

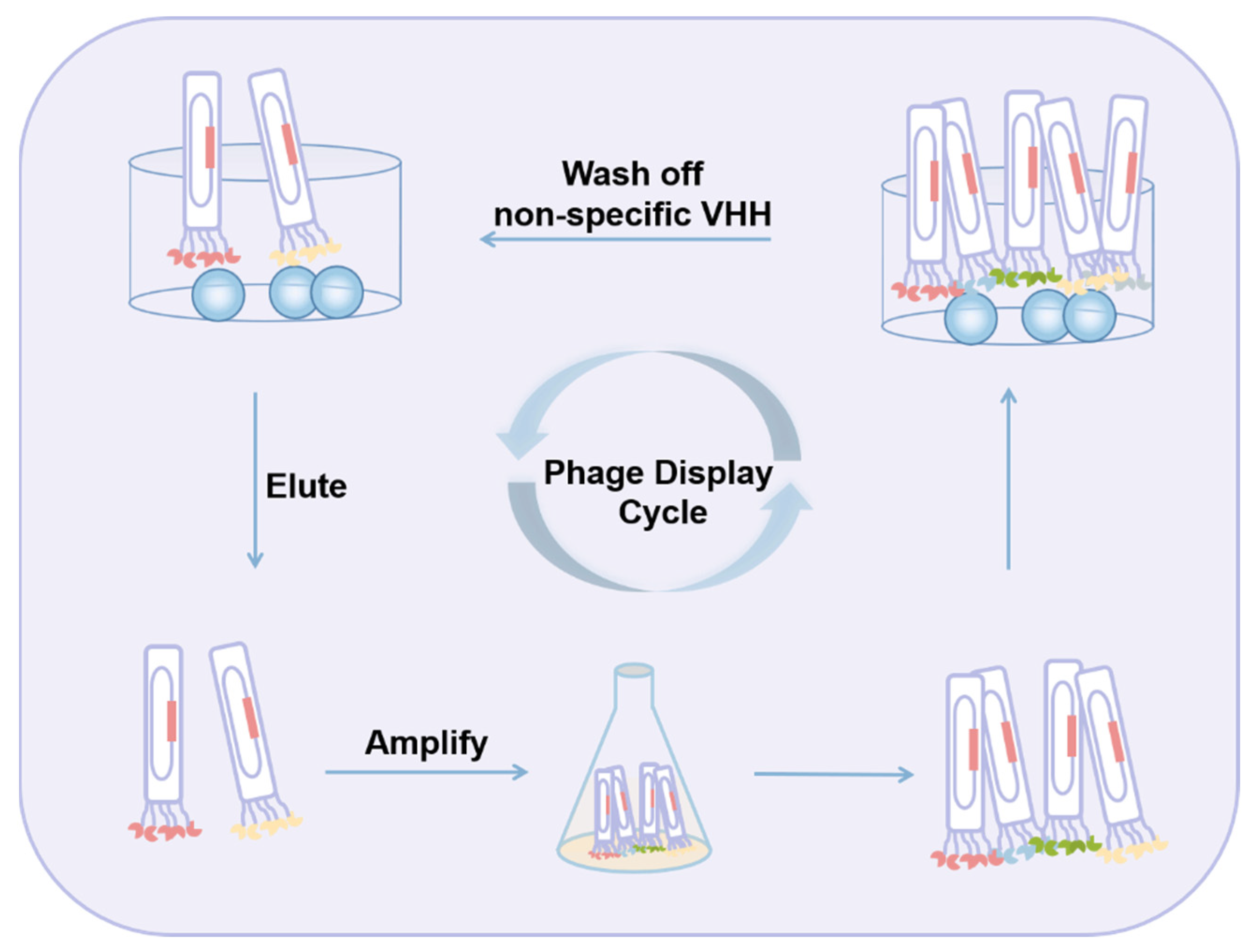
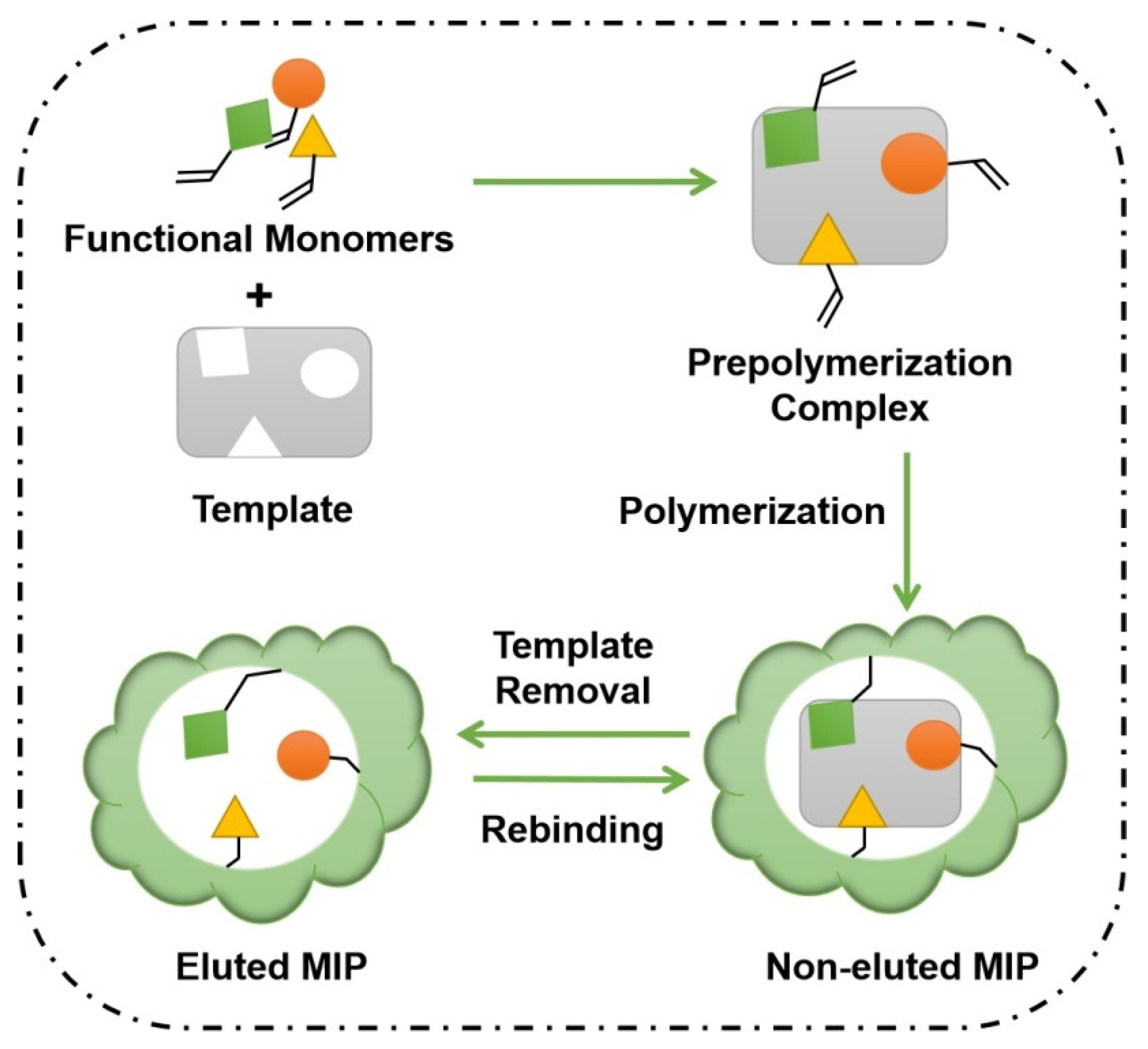
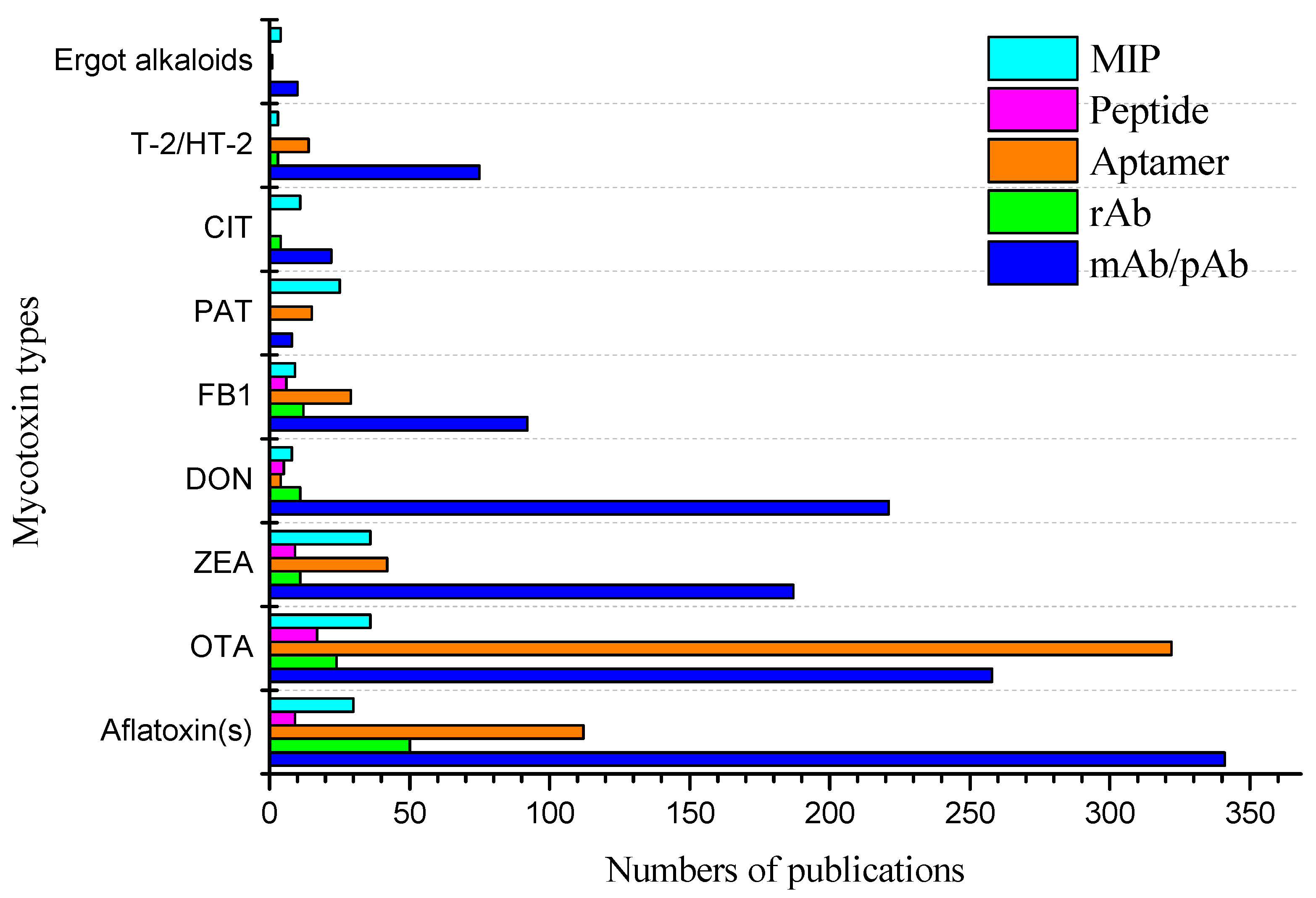

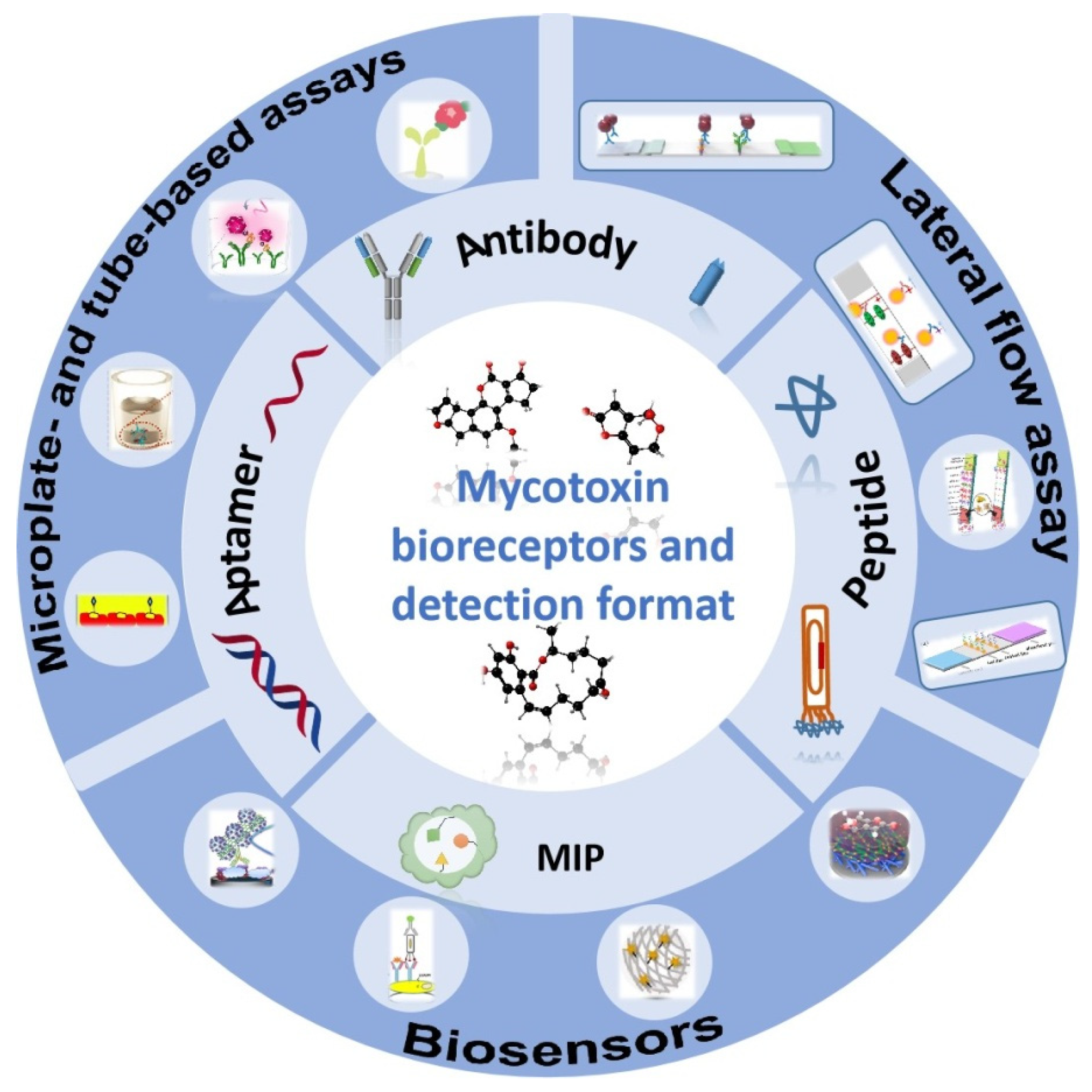{kind=link}
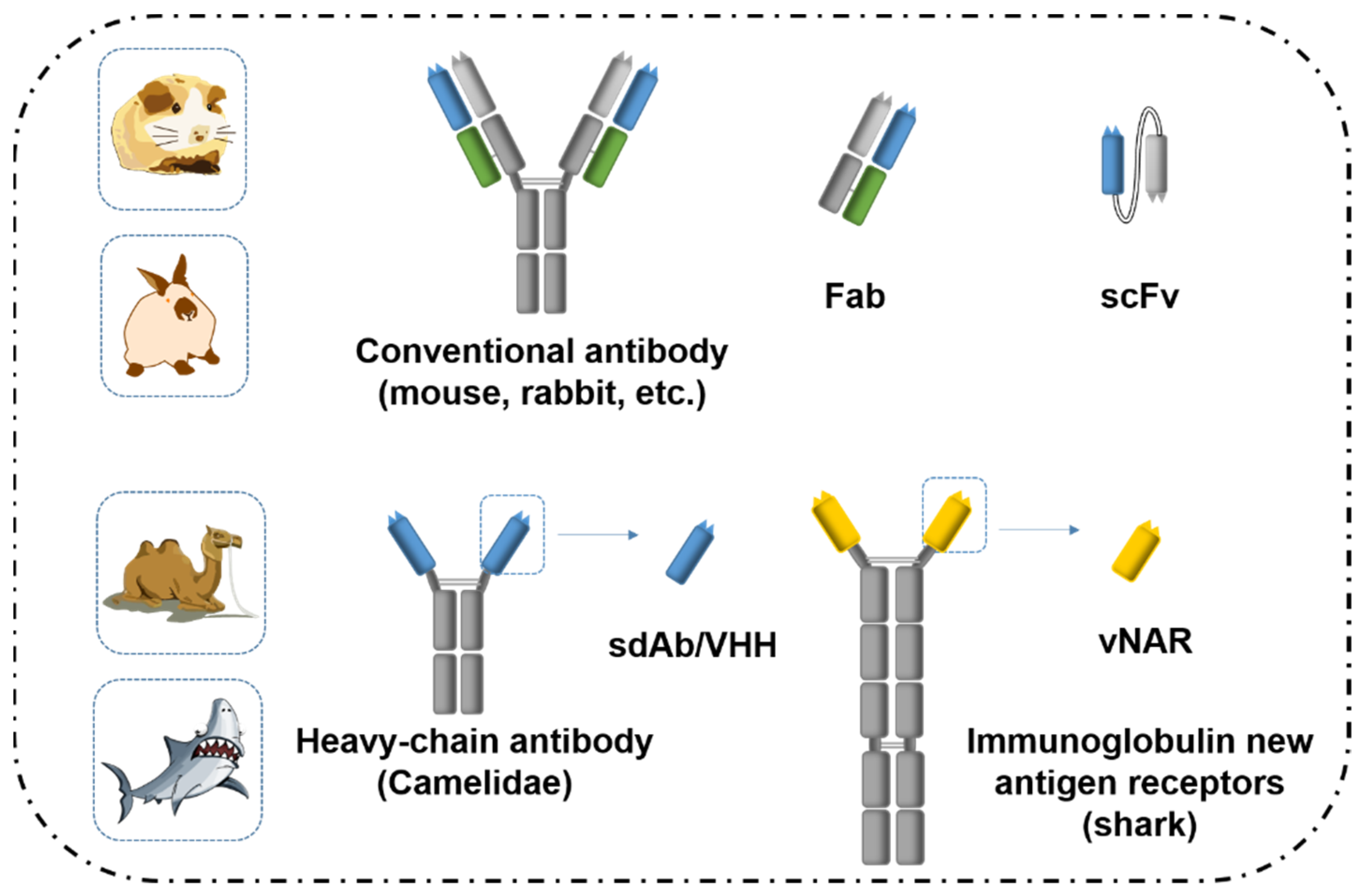{kind=link}
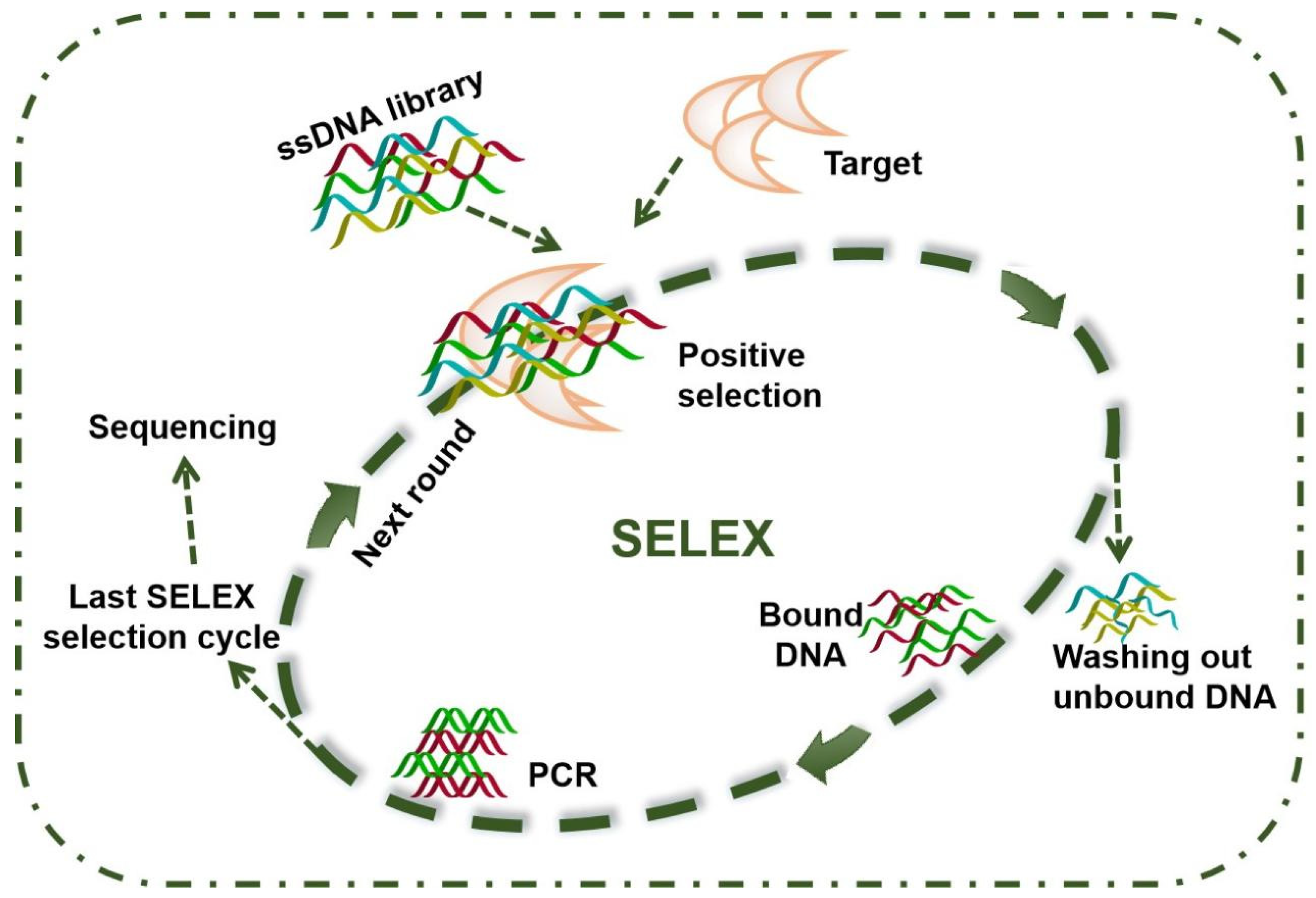{kind=link}
{kind=link}
{kind=link}
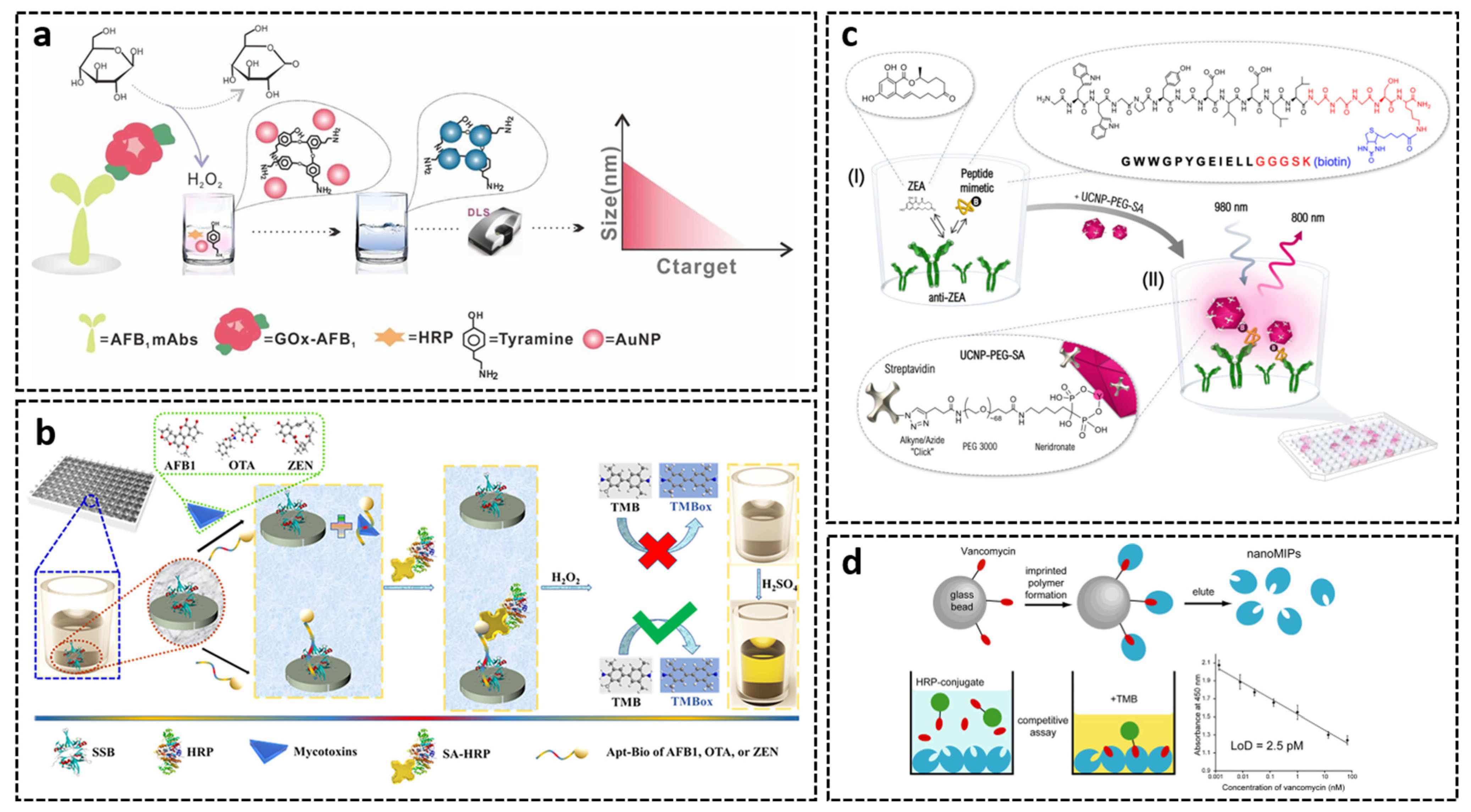{kind=link}
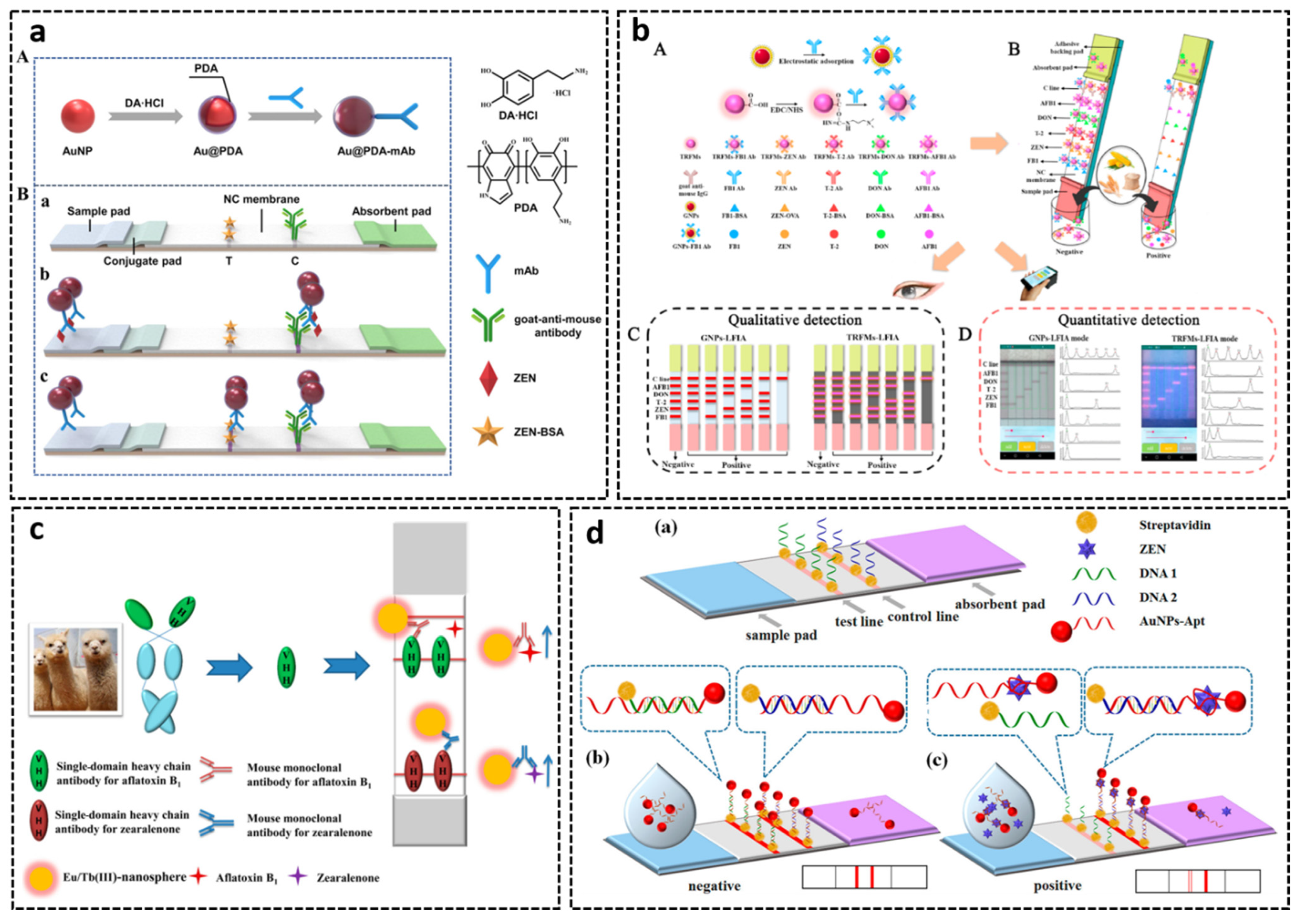{kind=link}
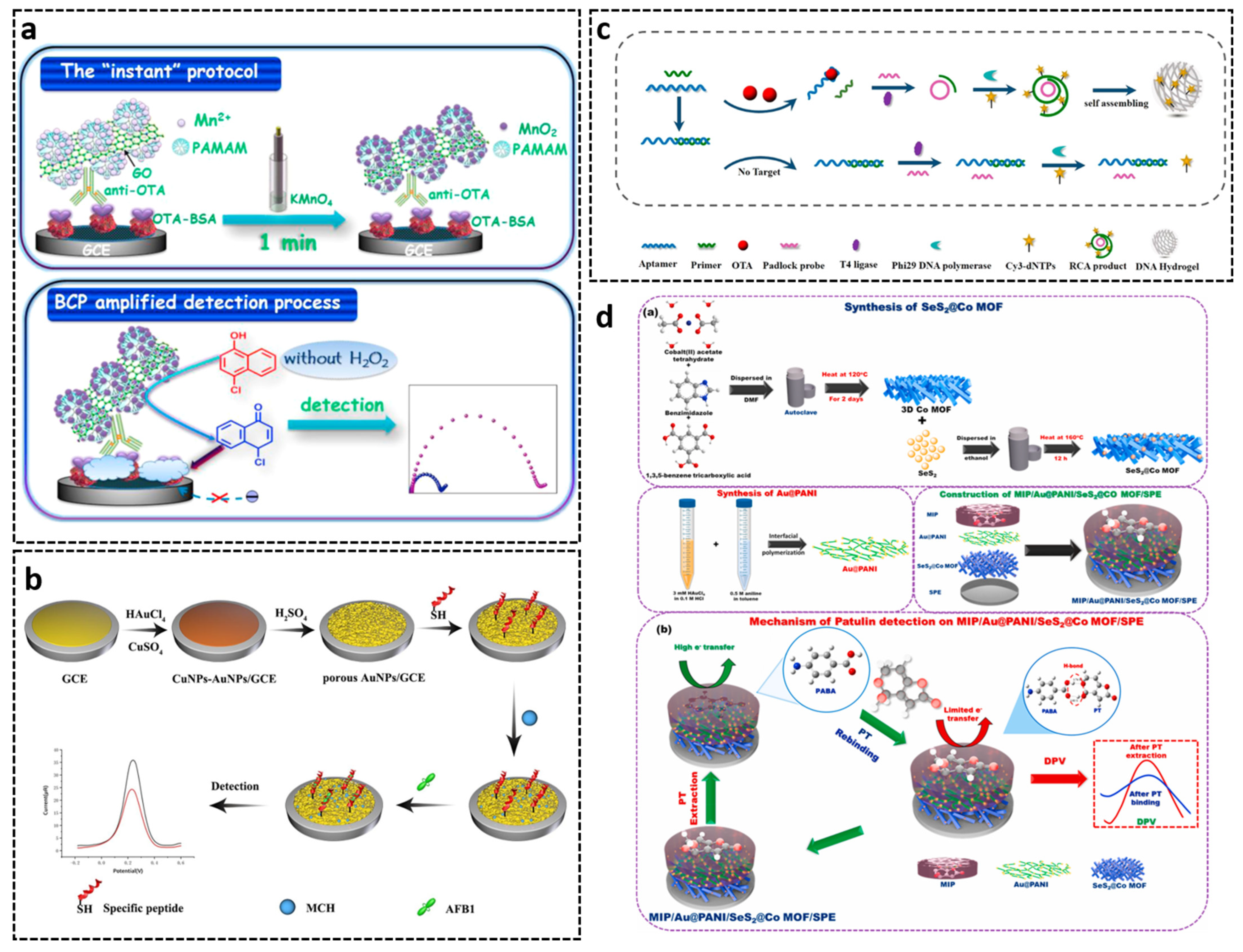{kind=link}
{kind=link}
| Mycotoxins | Structure | Main Fungi Species | Commodities Affected | Toxic Effects |
|---|---|---|---|---|
| Aflatoxins: B1, B2, G1, G2, M1* | AFB1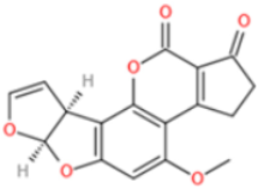 | Asp. flavus, Asp. parasiticus | Nuts, spices, grains such as maize, rice, wheat, * milk and milk products, etc. | Carcinogenic, teratogenic, mutagenic, immunosuppressive [39] |
| DON | DON | F. graminearum, F. culmorum, F. cerealis | Cereals, cereal products | Diarrhea, vomiting, anorexia, immune dysregulation [40] |
| ZEN | trans-ZEN cis-ZEN  | F. graminearum (Gibberella zeae), F. culmorum, F. cerealis, F. equiseti, F. crookwellense, F. semitectum | Cereals, cereal products, maize, rice, beer, etc. | Hyperoestrogenic, hepatotoxic, haematotoxic, immunotoxic, genotoxic [41] |
| OTA | OTA | Asp. ochraceus, Asp. niger, Asp. carbonarius, Asp. terreus, P. verrucosum, P. nordicum | Cereals, wine, coffee, cocoa, beans, dried fruits, nuts, spices, cheese, etc. | Nephrotoxic, hepatotoxic, neurotoxic, teratogenic, immunotoxic [42] |
| Fumonisins: FB1, FB2, FB3 | FB1 | F. verticillioides, F. proliferatum | Mainly maize and maize-based products, sorghum, asparagus | Carcinogenic, cytotoxic, nephrotoxic, hepatotoxic [43,44] |
| T-2/HT-2 toxin |  T-2 toxin  HT-2 toxin | F. langsethiae, F. poae, F. sporotrichioides | Wheat, rye, maize, soybeans | Growth retardation, myelotoxic, hemotoxic, necrotic lesions on contact sites [45] |
| PAT | 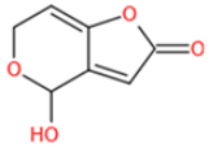 | P. expansum | Fruits and vegetables | Nausea, vomiting and other gastro- intestinal symptoms, kidney damage [46] |
| CIT |  | P. citrinum, P. camemberti, Asp. terreus, Asp. niveus | Fermented maize, cheese, corn, wheat, barley, red yeast rice, apples, brewed beer, cereal products | Nephrotoxic, may cause liver and kidney diseases, nervous system damage [47] |
| Mycotoxins | Country | Maximum Permitted Level (μg/kg) |
|---|---|---|
| AFs | China | 5–20 (0.5) *, (AFB1) |
| EU | 2–12 (0.1) *, (AFB1), 4–15, (sum of B1, B2, G1, G2), | |
| U.S. | 20, (sum of B1, B2, G1, G2), | |
| AFM1 | China | 0.5 |
| EU | 0.05 (0.025) * | |
| U.S. | 0.5 | |
| ZEN | China | 60 |
| EU | 50–400 (20) * | |
| U.S. | not set | |
| OTA | China | 2–10 |
| EU | 2–80 (0.5) * | |
| U.S. | not set | |
| DON | China | 1000 |
| EU | 500–1750, (200) * | |
| U.S. | 1000 | |
| PAT | China | 50 |
| EU | 25–50, (10) * | |
| U.S. | 50 | |
| FMs | China | in preparation |
| EU | 800–4000, (200) *, (FB1, FB2) | |
| T-2/HT-2 | U.S. | 2000–4000, (FB1, FB2, FB3) |
| China | not set | |
| EU | in preparation 2 | |
| CIT | EU | 2000 |
| EAs | EU | 100–500, (20) *, (sum of 12 compounds) |
| Mycotoxin(s) | Immunogen Structure | Coupling Method | Antibody Type | Titer | IC50 | LOD | Reference |
|---|---|---|---|---|---|---|---|
| Total AFs | 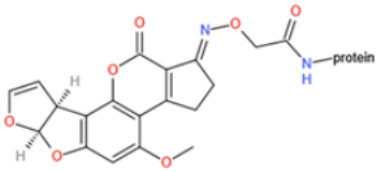 AFB1-oxime-BSA | Carbodiimide method | pAb | Higher than 1000 | AFB1 1.8 ng/mL AFB2 16 ng/mL AFG1 20 ng/mL AFG2 320 ng/mL | AFB1 0.4 ng/mL | [160] |
| AFB1 | 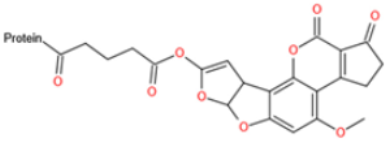 AFB2a-HG-BSA 1 | Mixed anhydride method | pAb | 710–800 | 0.15 ng/assay | 0.02 ng/assay | [137] |
| AFM1 | 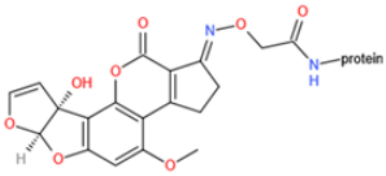 AFM1-BSA | Carbodiimide method | pAb and mAb | n.a. | 25 ng/mL (mAb); 0.5 ng/mL (pAb) | n.a. | [82] |
| OTA |  OTA-BSA | Carbodiimide method | pAb | n.a. | 3 ng/mL | 1 ng/mL | [161] |
| DON | 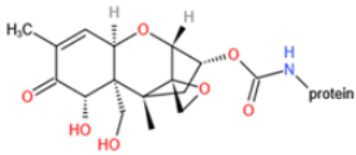 DON-BSA | N,N′-carbonyldiimidazole method | mAb | n.a. | 9.84 ng/mL | n.a. | [162] |
| T-2 toxin |  T-2-HS-BSA 2 | Carbodiimide method | pAb | 303 | 3.5 ng/assay | 1 ng/assay | [146] |
| ZEN | 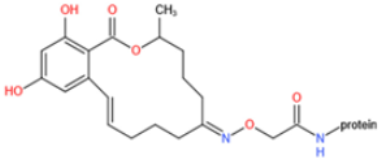 ZEN-oxime-BSA | Mixed anhydride procedure | pAb | 5120 | n.a. | 0.5 ng/mL | [149] |
 5-NH2-ZEN-BSA | Glutaraldehyde method | mAb | 520 | 11.2 ng/mL | 0.3 ng/mL | [153] | |
| ZEN | 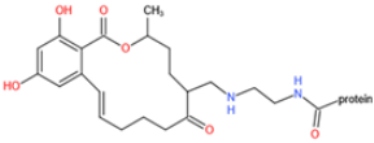 | Mannich reaction | pAb and mAb | 30,000 | 233.35 ng/mL (pAb); 55.72 ng/mL (mAb) | n.a. | [154] |
 | 1,4-Butanediol diglycidyl ether method | mAb | 1.024 × 106 | 1.115 ng/mL | n.a. | [155] | |
| FB1 | 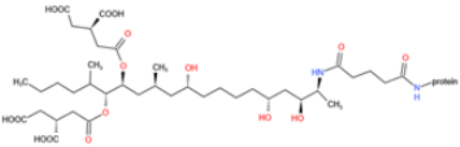 FB1-KLH | Glutaraldehyde method | pAb | 10,000 | 0.45 ng/mL | 0.1 ng/mL | [163] |
| CIT | 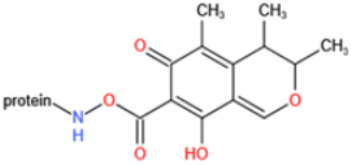 CIT-BSA | Activated ester method | mAb | 32,000 | 0.28 ng/mL | 0.01 ng/mL | [164] |
| PAT | 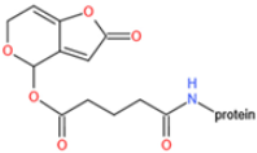 PAT-HG-BSA | Carbodiimide method | pAb | 1100 | n.a. | n.a. | [165] |
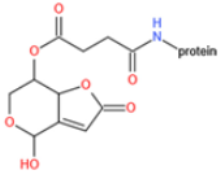 PAT-Ins-HS-BSA 3 | Carbodiimide method | pAb | n.a. | n.a. | 10 ng/mL | [166] | |
 PAT-Sat-HS-BSA 4 | Carbodiimide method | pAb | n.a. | n.a. | 10 ng/mL | [166] |
| Target | Sequence (5′-3′) | KD | Reference |
|---|---|---|---|
| AFB1 | GT TGG GCA CGT GTT GTC TCT CTG TGT CTC GTG CCC TTC GCT AGG CCC ACA | n.a. * | [181] |
| AFM1 | ACT GCT AGA GAT TTT CCA CAT | n.a. | [190] |
| OTA | GAT CGG GTG TGG GTG GCG TAA AGG GAG CAT CGG ACA | 0.2 μM | [180] |
| FB1 | ATA CCA GCT TAT TCA ATT AAT CGC ATT ACC TTA TAC CAG CTT ATT CAA TTA CGT CTG CAC ATA CCA GCT TAT TCA ATT AGA TAG TAA GTG CAA TCT | 100 ± 30 nM | [191] |
| ZEN | TCATCTATCTATGGTACATTACTATCTGTAATGTGATATG | 41 ± 5 nM | [192] |
| DON | GCATCACTACAGTCATTACGCATCGTAGGGGGGATCGTTAAGGAAGTGCCCGGAGGCGGTATCGTGTGAAGTGCTGTCCC | n.a. | [185] |
| PAT | GGCCCGCCAACCCGCATCATCTACACTGATATTTTACCTT | 21.83 ± 5.022 nM | [187] |
| T-2 | GTATATCAAGCATCGCGTGTTTACACATGCGAGAGGTGAA | 20.8 ± 3.1 nM | [188] |
| Ergot alkaloids | ACTCATCTGTGAAGAGAAGCAGCACAGAGGTCAGATGTCCGTCAGCCCCGATCGCCATCCAGGGACTCCCCCCTATGCCTATGCGTGCTACCGTGAA | 44 nM 2 | [189] |
| Mycotoxin | U.S.: Company/location/website * | ||||||
| Eurofins Abraxis Warminster, PA USA abraxis.eurofins-technologies.com | Neogen Lansing, MI U.S. www.neogen.com | Beacon Analytical Systems, Inc. Saco, ME USA www.beaconkits.com | Envirologix Inc. Portland, Main USA www.envirologix.com | Charm Sciences, Inc. Lawrence, MA USA www.charm.com | Vicam Milford, MA USA www.vicam.com | Romer Labs, Inc Newark, DE USA www.romerlabs.com | |
| AFB1 | n.a. | n.a. | n.a. | n.a. | n.a. | IAC | ELISA, LFD |
| AFB2 | n.a. | n.a. | n.a. | n.a. | n.a. | IAC | n.a. |
| AFG1 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| AFG2 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| Total AFs | ELISA | ELISA, IAC, LFD | ELISA (plate, tube) | LFD | LFD | IAC, LFD | ELISA, LFD |
| AFM1 | ELISA | ELISA, LFD | ELISA | n.a. | LFD | IAC, LFD | ELISA |
| OTA | ELISA (OTA, B, C) | ELISA, LFD | n.a. | LFD | n.a. | IAC | ELISA, LFD |
| ZEN | IAC, LFD | n.a. | ELISA (plate, tube) | LFD | LFD | IAC, LFD | ELISA, LFD |
| DON | ELISA | ELISA, IAC, LFD | ELISA | LFD | LFD | IAC, LFD | ELISA, LFD |
| FB1 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | LFD |
| Total FMs | n.a. | ELISA, LFD | ELISA | LFD | LFD | IAC, LFD | ELISA, LFD |
| T-2 | n.a. | n.a. | ELISA | n.a. | n.a. | IAC, LFD | ELISA |
| T-2/HT-2 | n.a. | ELISA, LFD | ELISA | LFD | LFD | IAC, LFD | n.a. |
| PAT | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| CIT | n.a. | n.a. | ELISA | n.a. | n.a. | IAC | n.a. |
| Ergot alkaloids | n.a. | LFD | n.a. | n.a. | n.a. | n.a. | n.a. |
| Mycotoxin | Europe: Company/location/website | ||||||
| Biosense Laboratories AS Bergen Norway www.biosense.com | R-Biopharm Darmstadt Germany r-biopharm.com | Abcam, Inc. Cambridge U.K. www.abcam.com (abcam.cn abcam.jp) | antibodies-online GmbH Aachen# Germany www.antikoerper-online.de | Agrisera AG Umea Sweden www.agrisera.com | Biomol Hamburg Germany www.biomol.com | Aokin AG Berlin Germany www.aokin.com | |
| AFB1 | n.a. | ELISA | mAb | pAb, mAb, ELISA | ELISA | mAb, ELISA, LFD | mAb |
| AFB2 | n.a. | n.a. | n.a. | n.a. | n.a. | mAb | n.a. |
| AFG1 | n.a. | n.a. | n.a. | n.a. | n.a. | mAb | n.a. |
| AFG2 | n.a. | n.a. | mAb | n.a. | n.a. | mAb | n.a. |
| Total AFs | ELISA | ELISA, LFD, IAC | n.a. | mAb, ELISA | ELISA | mAb, ELISA, LFD | IAC |
| AFM1 | n.a. | ELISA | n.a. | ELISA | pAb | mAb, ELISA, LFD | mAb, IAC |
| OTA | n.a. | ELISA, IAC | pAb, mAb | ELISA | pAb | mAb, ELISA, LFD | mAb, IAC |
| ZEN | ELISA | ELISA, LFD, IAC | mAb | pAb, mAb, ELISA | ELISA | mAb, ELISA, LFD | mAb, IAC |
| DON | ELISA | ELISA, LFD, IAC | pAb | pAb, mAb, ELISA | pAb | mAb, ELISA, LFD | mAb, IAC |
| FB1 | n.a. | ELISA, LFD, IAC | mAb | ELISA | n.a. | mAb, ELISA, LFD | mAb, IAC |
| Total FMs | ELISA | n.a. | n.a. | n.a. | n.a. | mAb | n.a. |
| T-2 | ELISA | ELISA, IAC | n.a. | ELISA | n.a. | ELISA, LFD | mAb |
| T-2/HT-2 | n.a. | ELISA, LFD, IAC | n.a. | n.a. | n.a. | n.a. | mAb, IAC |
| PAT | n.a. | MISPE | n.a. | pAb | pAb | n.a. | n.a. |
| CIT | n.a. | ELISA, IAC | n.a. | n.a. | n.a. | n.a. | n.a. |
| Ergot alkaloids | n.a. | n.a. | n.a. | n.a. | n.a. | mAb | mAb |
| Mycotoxin | China: Company/location/website | ||||||
| Cusabio Technology Co., Ltd. Wuhan cusabio.cn | Lvdu Bio-sciences 6 Technology Co., Ltd. Binzhou, Shandong lvdu.net | Jiangsu Suwei Microbiological Research Co., Ltd. Wuxi jssuwei.com | Beijing WDWK Biotechnology Co., Ltd. Beijing wdwkbio.com | Nankai Biotech Co. Ltd. Hangzhou nkbiotech.com | Beijing KWINBON Biotechnology Co., Ltd. Beijing kwinbon.com | Shandong Meizheng Bio-Tech Co., Ltd. Rizhao, Shandong meizhengbio.com | |
| AFB1 | ELISA | mAb, IAC, LFD, ELISA | ELISA, LFD, IAC | ELISA, LFD | LFD | ELISA, LFD, IAC | ELISA, LFD, IAC |
| AFB2 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| AFG1 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| AFG2 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| Total AFs | ELISA | IAC, LFD | ELISA, IAC | n.a. | n.a. | ELISA | ELISA, LFD, IAC |
| AFM1 | ELISA | mAb, IAC, LFD, ELISA | ELISA, LFD, IAC | ELISA, LFD | LFD | ELISA, LFD | ELISA, LFD, IAC |
| OTA | ELISA | mAb, IAC, LFD, ELISA | ELISA, LFD, IAC | ELISA, LFD | LFD | ELISA, IAC | ELISA |
| ZEN | ELISA | mAb, IAC, LFD, ELISA | ELISA, LFD, IAC | ELISA, LFD | LFD | ELISA, LFD, IAC | ELISA, LFD |
| DON | ELISA | mAb, IAC, LFD, ELISA | ELISA, LFD, IAC | ELISA, LFD | LFD | ELISA, LFD, IAC | ELISA, LFD, IAC |
| FB1 | ELISA | mAb, IAC, ELISA | n.a. | n.a. | n.a. | n.a. | n.a. |
| Total FMs | n.a. | LFD, ELISA | ELISA | ELISA, LFD | LFD | ELISA, LFD | ELISA, LFD |
| T-2 | n.a. | mAb, IAC, LFD, ELISA | ELISA | ELISA, LFD | LFD | ELISA | ELISA |
| T-2/HT-2 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| PAT | n.a. | mAb | n.a. | n.a. | n.a. | n.a. | n.a. |
| Mycotoxin | China: Company/location/website | ||||||
| Cusabio Technology Co., Ltd. Wuhan cusabio.cn | Lvdu Bio-sciences 6 Technology Co., Ltd. Binzhou, Shandong lvdu.net | Jiangsu Suwei Microbiological Research Co., Ltd. Wuxi jssuwei.com | Beijing WDWK Biotechnology Co., Ltd. Beijing wdwkbio.com | Nankai Biotech Co. Ltd. Hangzhou nkbiotech.com | Beijing KWINBON Biotechnology Co., Ltd. Beijing kwinbon.com | Shandong Meizheng Bio-Tech Co., Ltd. Rizhao, Shandong meizhengbio.com | |
| CIT | n.a. | IAC | n.a. | n.a. | n.a. | n.a. | n.a. |
| Ergot alkaloids | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. |
| Provider | Principle | Internet Address * |
|---|---|---|
| Luminex Corporation, Austin, TX, USA | suspension assay | www.luminexcorp.com |
| Becton Dickinson Biosciences, Franklin Lakes, NJ, USA | suspension assay | www.bdbiosciences.com |
| Quanterix Corp., Lexington, MA, USA | suspension assay | www.Quanterix.com |
| Merck Millipore, Burlington, MA, USA | suspension assay | www.merckmillipore.com |
| Bio-Rad-Laboratories, Hercules, CA, USA | suspension assay | www.bio-rad.com |
| SAFIA Technologies GmbH, Berlin, Germany | suspension assay | www.safia.tech |
| Foss GmbH, Hamburg, Germany | suspension assay | www.fossanalytics.com |
| Unisensor Seraing, Belgium | suspension assay | www.unisensor.be |
| Randox-Laboratories, Crumlin, UK | planar array (biochip) | www.randoxfood.com |
| GWK Präzisionstechnik GmbH, München, Germany | planar array (biochip) | www.gwk-munich.com |
| Provider | Principle 1 | Internet Address * |
|---|---|---|
| GE Healthcare, Chicago, IL, USA | SPR | www.biacore.com/lifesciences |
| Biolin Scientific, Gothenburg, Sweden | QCM | www.biolinscientific.com/qsense |
| Micromotive GmbH, Mainz Germany | Microcantilever array | www.micromotive.de |
| 2bind GmbH, Regensburg, Germany | Bio-layer-interferometrie | www.2bind.com |
| Dynamic Biosensors GmbH, München, Germany 2 | ESB | www.dynamic-biosensors.com |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, C.; Wang, J.; Knopp, D. Recent Progress in Rapid Determination of Mycotoxins Based on Emerging Biorecognition Molecules: A Review. Toxins 2022, 14, 73. https://doi.org/10.3390/toxins14020073
Wang Y, Zhang C, Wang J, Knopp D. Recent Progress in Rapid Determination of Mycotoxins Based on Emerging Biorecognition Molecules: A Review. Toxins. 2022; 14(2):73. https://doi.org/10.3390/toxins14020073
Chicago/Turabian StyleWang, Yanru, Cui Zhang, Jianlong Wang, and Dietmar Knopp. 2022. "Recent Progress in Rapid Determination of Mycotoxins Based on Emerging Biorecognition Molecules: A Review" Toxins 14, no. 2: 73. https://doi.org/10.3390/toxins14020073
APA StyleWang, Y., Zhang, C., Wang, J., & Knopp, D. (2022). Recent Progress in Rapid Determination of Mycotoxins Based on Emerging Biorecognition Molecules: A Review. Toxins, 14(2), 73. https://doi.org/10.3390/toxins14020073