Comparative Cytotoxic Effects and Possible Mechanisms of Deoxynivalenol, Zearalenone and T-2 Toxin Exposure to Porcine Leydig Cells In Vitro


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
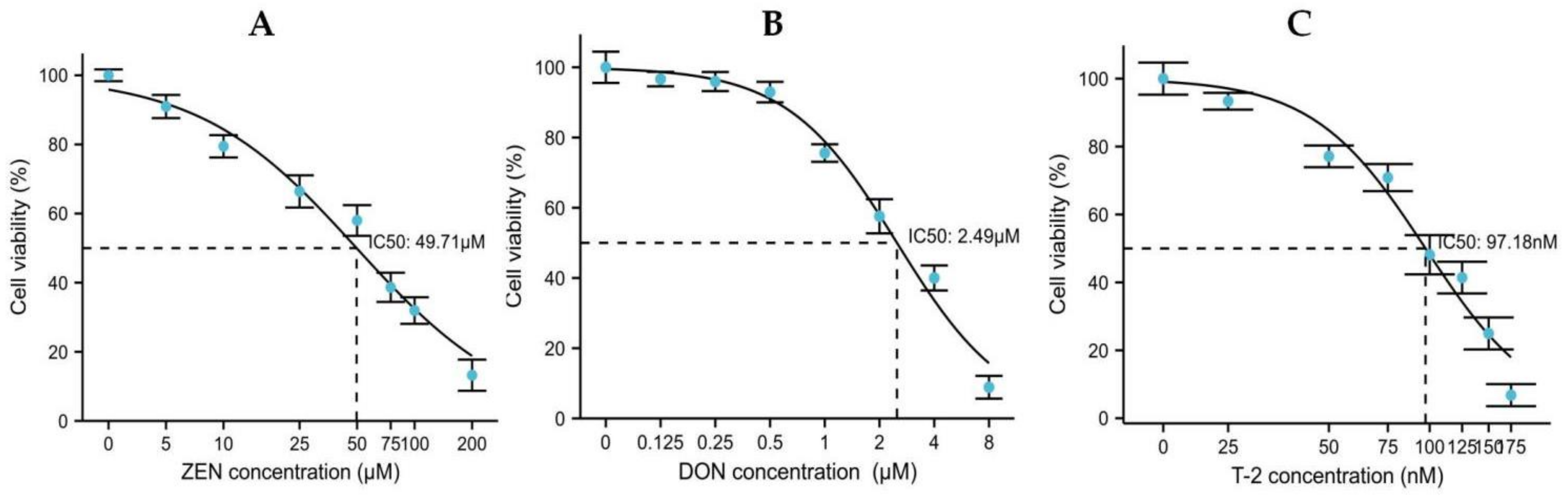
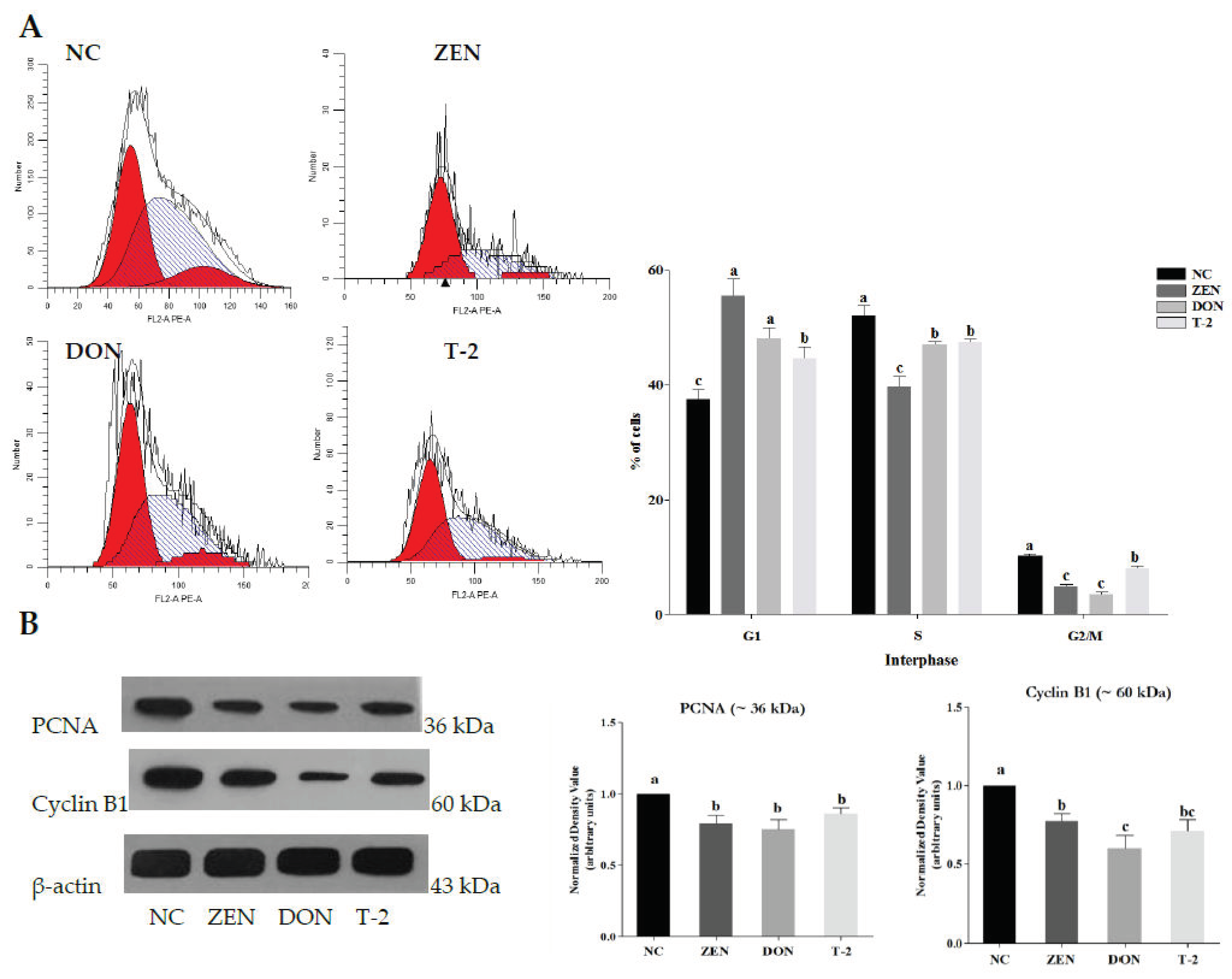
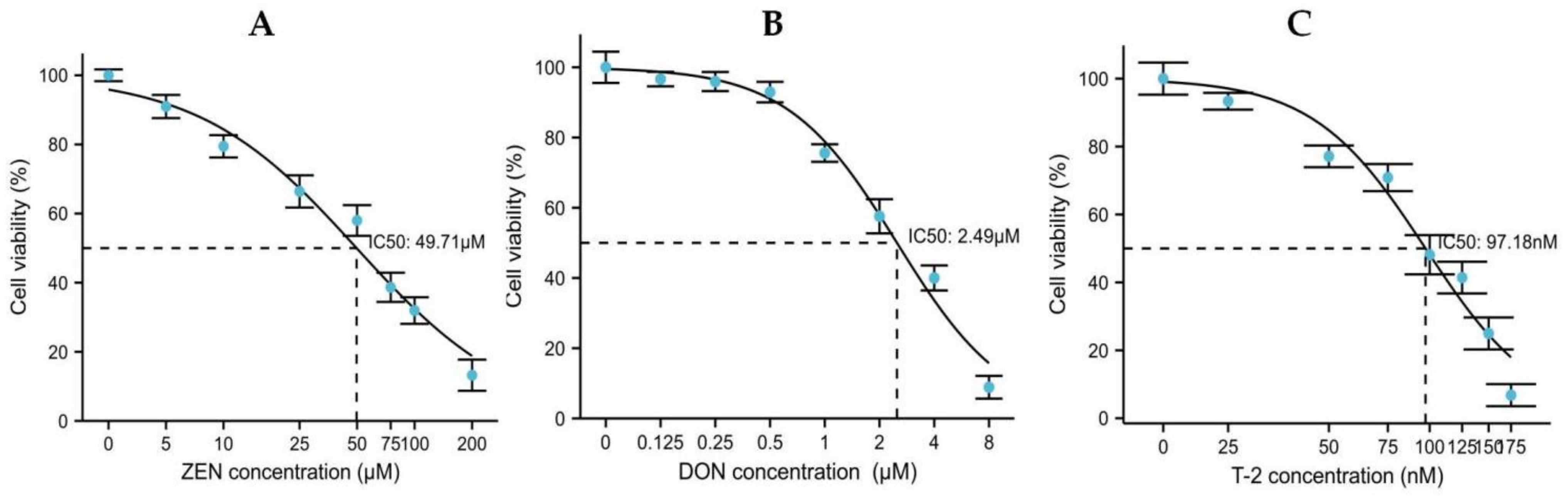
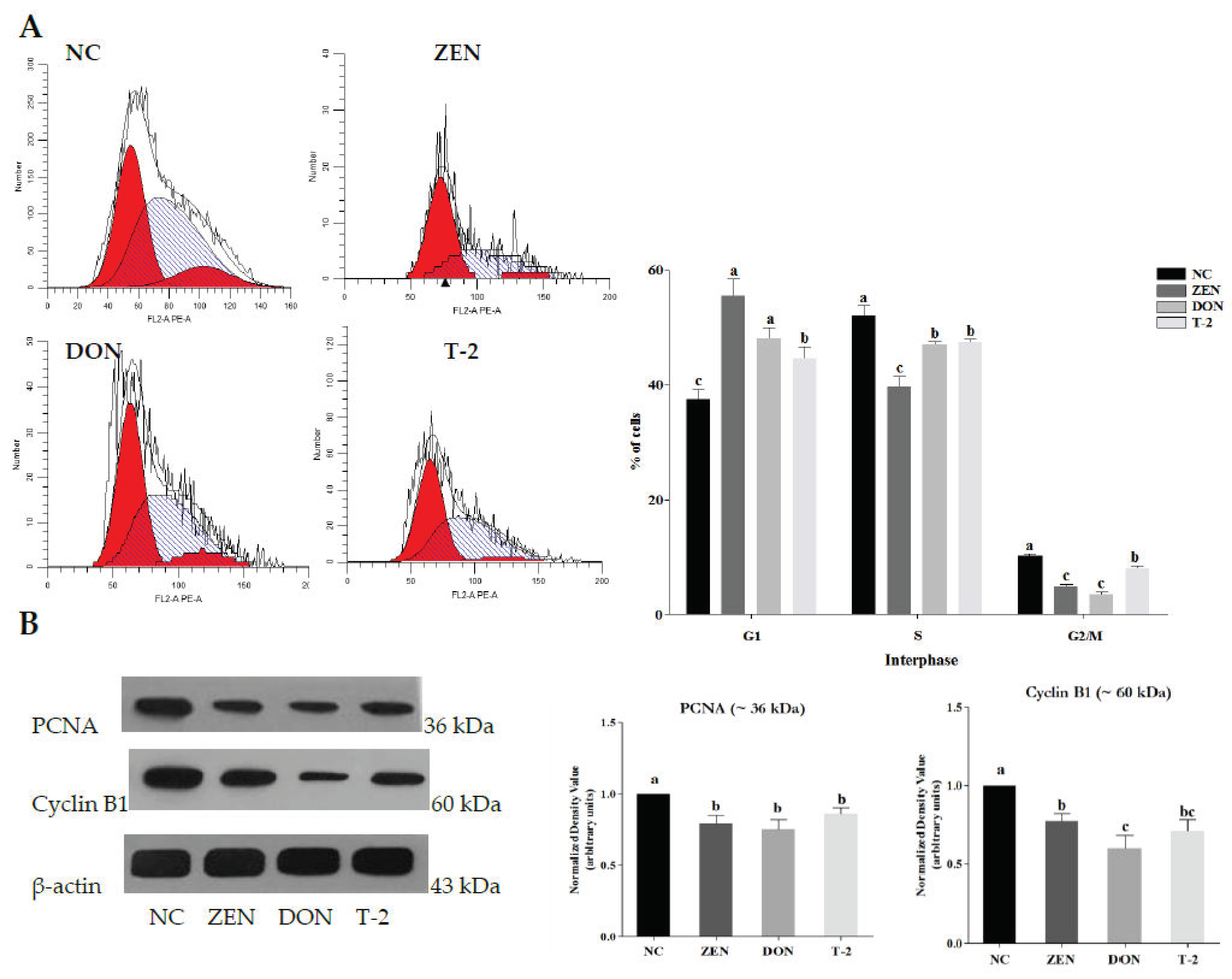
2.1. Effects of ZEN, DON and T-2 on Cell Proliferation
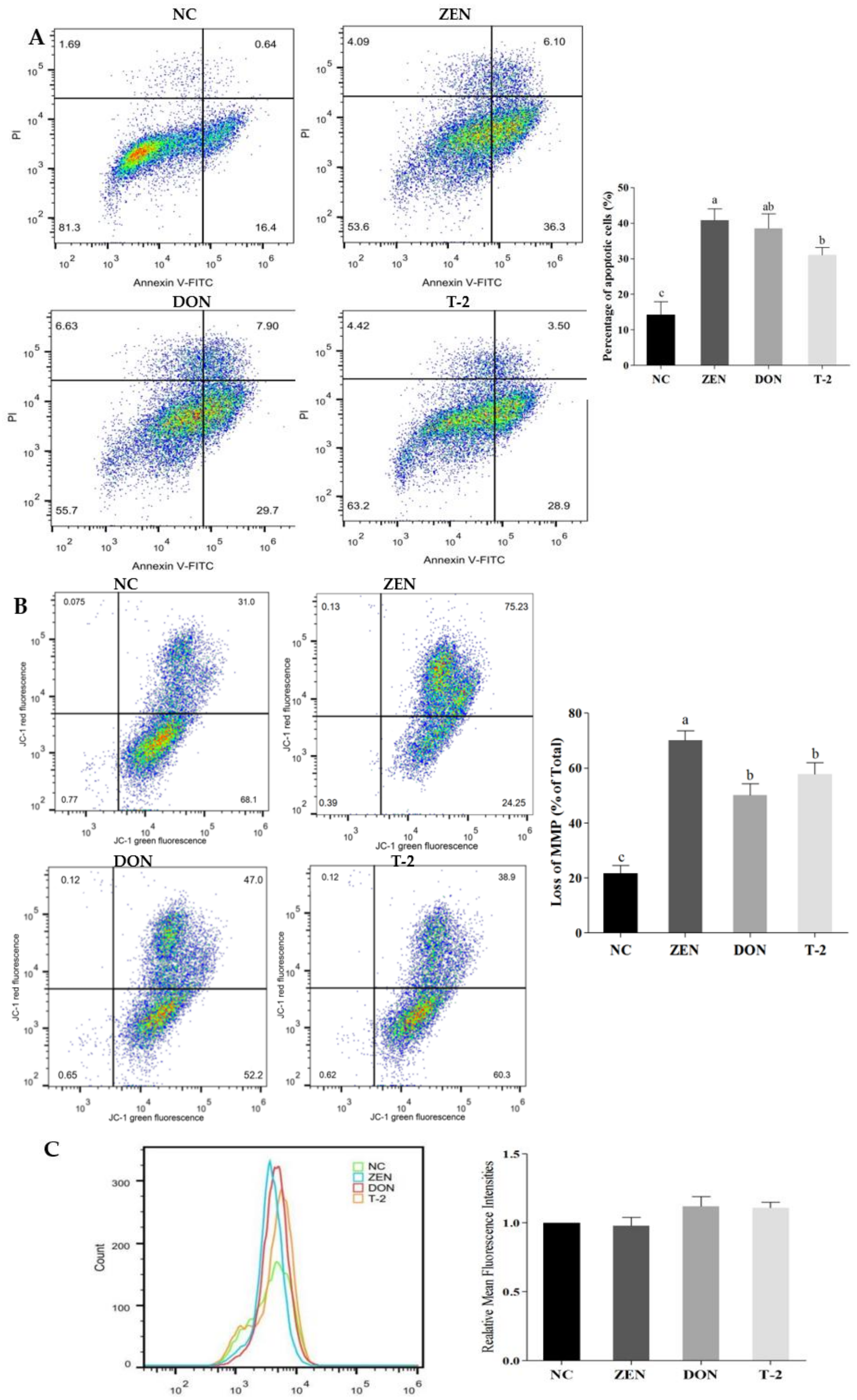
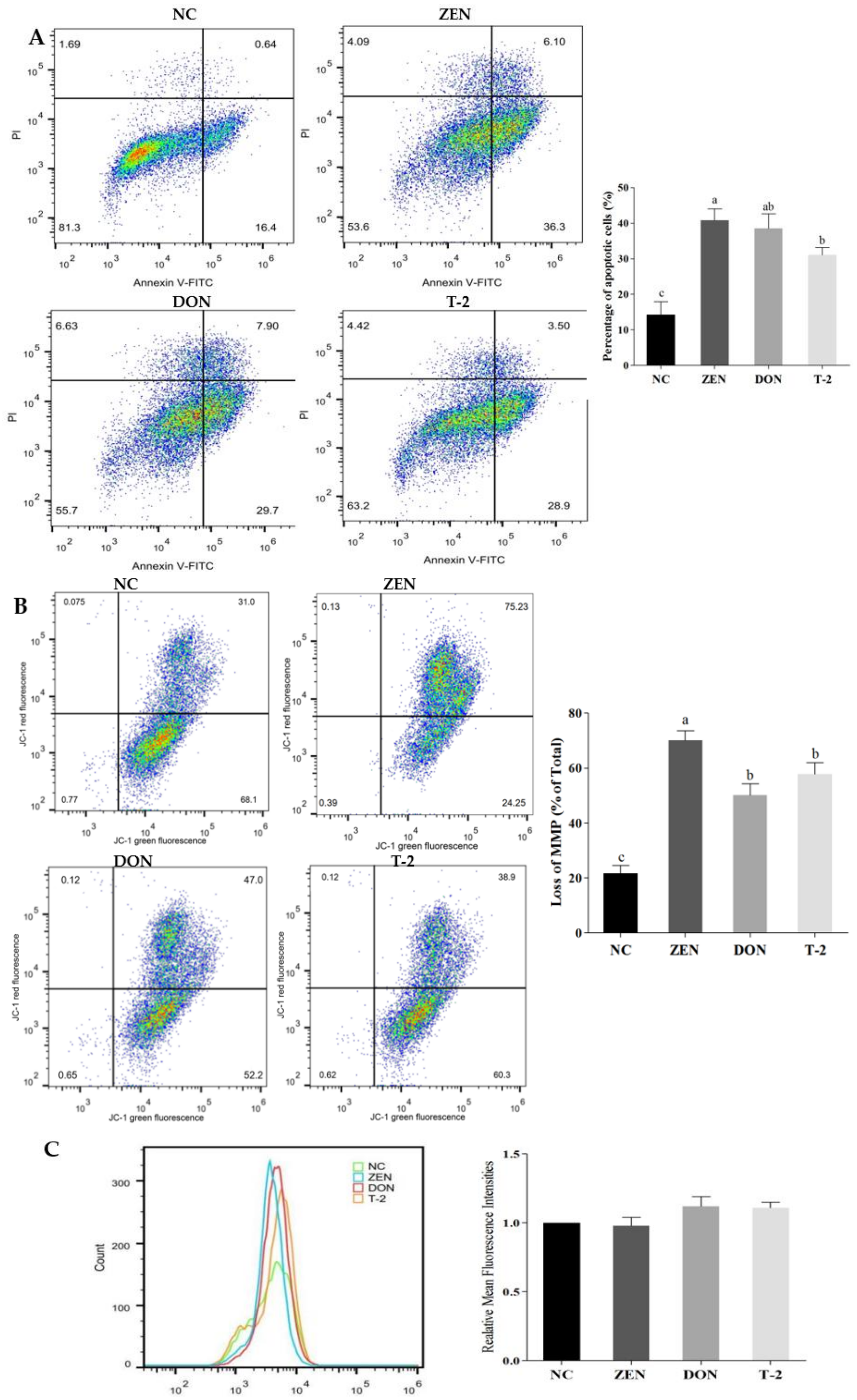
2.2. Effects of ZEN, DON and T-2 on Cell Apoptosis
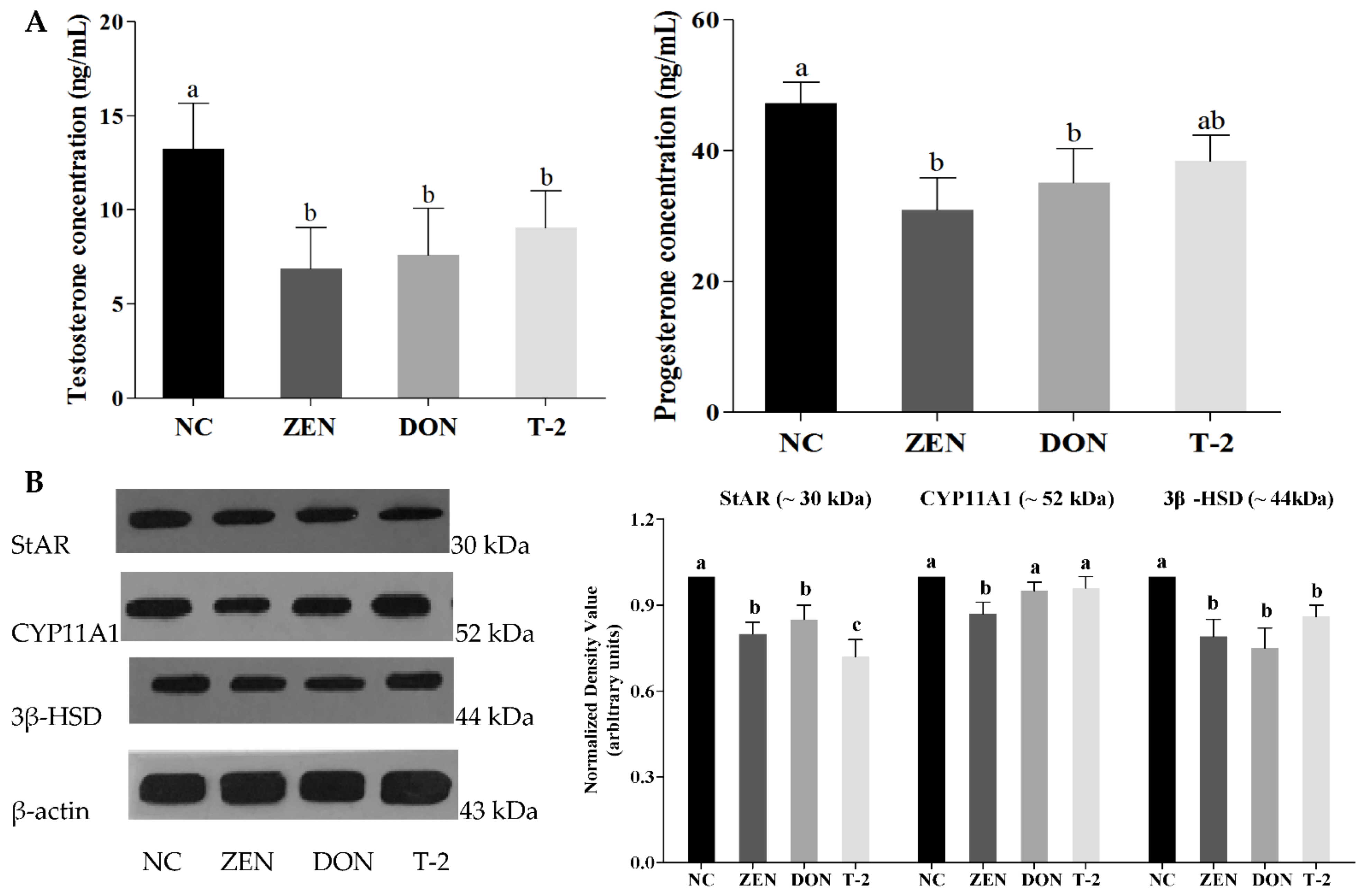
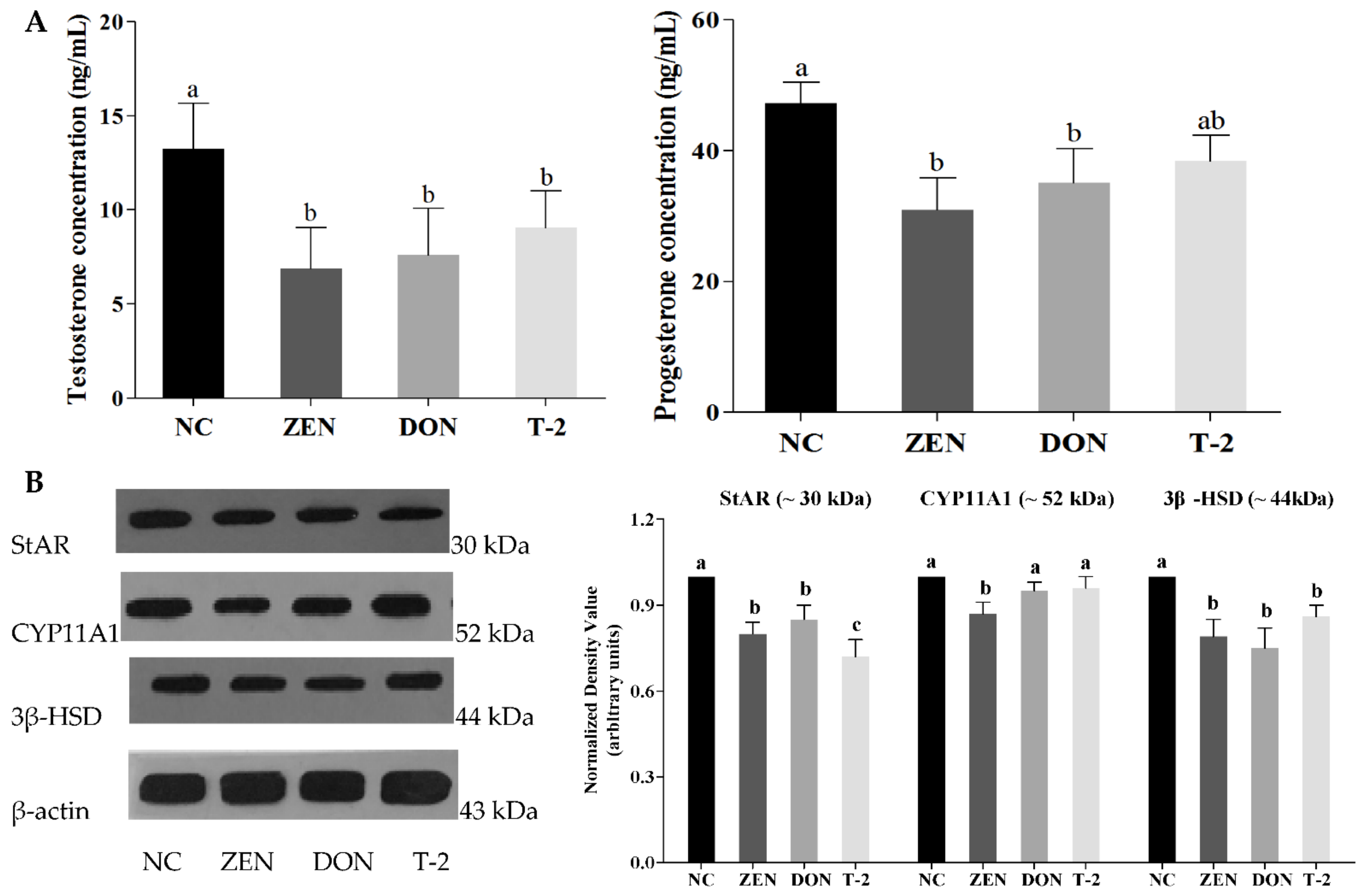
2.3. Effects of ZEN, DON and T-2 on Steroid Hormone Biosynthesis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Animals
5.3. Isolation and Culture of Porcine Leydig Cell
5.4. Cell Treatment
5.5. Cell Proliferation Assay
5.6. Cell Cycle Assay
5.7. Cell Apoptosis Assay
5.8. MMP Assay
5.9. ROS Production Assay
5.10. Steroid Hormone Quantifications
5.11. Western Blot Analysis
5.12. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tremblay, J.J. Molecular regulation of steroidogenesis in endocrine Leydig cells. Steroids 2015, 103, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Bai, S.; Bai, W.; Zou, F.; Zhang, L.; Su, Z.; Zhang, Q.; Ou, S.; Huang, Y. Toxic mechanisms of 3-monochloropropane-1,2-diol on progesterone production in r2c rat Leydig cells. J. Agric. Food Chem. 2013, 61, 9955–9960. [Google Scholar] [CrossRef] [PubMed]
- Savard, C.; Nogues, P.; Boyer, A.; Chorfi, Y. Prevention of deoxynivalenol- and zearalenone-associated oxidative stress does not restore ma-10 Leydig cell functions. Toxicology 2016, 341, 17–27. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, Y.; Gu, J.; Yuan, Y.; Liu, X.; Zheng, W.; Huang, Q.; Liu, Z.; Bian, J. Zearalenone inhibits testosterone biosynthesis in mouse Leydig cells via the crosstalk of estrogen receptor signaling and orphan nuclear receptor nur77 expression. Toxicol. In Vitro 2014, 28, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Pailhoux, E.; Mandonpepin, B.; Cotinot, C.; Geisert, R.D.; Niemann, H.; Doberska, C. Mammalian gonadal differentiation: The pig model. Reprod. Suppl. 2001, 58, 65–80. [Google Scholar] [CrossRef]
- Lejeune, H.; Habert, R.; Saez, J.M. Origin, proliferation and differentiation of Leydig cells. J. Mol. Endocrinol. 1998, 20, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Ki, M.; Vuli, A.; Kudumija, N.; Arkanj, B.; Pleadin, J. A two-year occurrence of fusarium T-2 and HT-2 toxin in Croatian cereals relative of the regional weather. Toxins 2021, 13, 39. [Google Scholar]
- Crudo, F.; Varga, E.; Aichinger, G.; Galaverna, G.; Marko, D.; Dall’Asta, C.; Dellafiora, L. Co-occurrence and combinatory effects of alternaria mycotoxins and other xenobiotics of food origin: Current scenario and future perspectives. Toxins 2019, 11, 640. [Google Scholar] [CrossRef] [Green Version]
- Tran, V.N.; Viktorova, J.; Augustynkova, K.; Jelenova, N.; Dobiasova, S.; Rehorova, K.; Fenclova, M.; Stranska-Zachariasova, M.; Vitek, L.; Hajslova, J. In silico and in vitro studies of mycotoxins and their cocktails; their toxicity and its mitigation by silibinin pre-treatment. Toxins 2020, 12, 148. [Google Scholar] [CrossRef] [Green Version]
- Ficheux, A.S.; Sibiril, Y.; Parent-Massin, D. Co-exposure of fusarium mycotoxins: In vitro myelotoxicity assessment on human hematopoietic progenitors. Toxicon 2012, 211, 1171–1179. [Google Scholar] [CrossRef]
- Kondreddy, R.; Song, J.; Hyun-Jeong, L.; Minseok, K.; Dong-Wook, K.; Hyun, J.; Bumseok, K.; Yookyung, L.; Yu, D.; Dong-Woon, K. Effects of high levels of deoxynivalenol and zearalenone on growth performance, and hematological and immunological parameters in pigs. Toxins 2018, 10, 114. [Google Scholar]
- Bonde, J.P. Male reproductive organs are at risk from environmental hazards. Asian J. Androl. 2010, 12, 152–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rykaczewska, A.; Gajęcka, M.; Onyszek, E.; Cieplińska, K.; Zielonka, Ł. Imbalance in the blood concentrations of selected steroids in pre-pubertal gilts depending on the time of exposure to low doses of zearalenone. Toxins 2019, 11, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.J.; Xiao, Y.; Li, C.; Zhang, J.; Zhang, Y.; Wu, M.; Ma, T.; Yang, L.; Wang, X.; Jiang, H.; et al. Zearalenone perturbs the circadian clock and inhibits testosterone synthesis in mouse Leydig cells. J. Toxicol. Environ. Health A 2021, 84, 112–124. [Google Scholar] [CrossRef]
- Chen, J.; Wang, M.; Wang, H.; Long, M. Zearalenone promotes apoptosis of mouse Leydig cells by targeting phosphatase and tensin homolog and thus inhibiting the PI3K/AKT signal pathway. Environ. Sci. Pollut. Res. Int. 2021, 28, 67779–67787. [Google Scholar] [CrossRef]
- Li, L.; Zhang, T.; Ren, X.; Li, B.; Wang, S. Male reproductive toxicity of zearalenone-meta-analysis with mechanism review. Ecotoxicol. Environ. Saf. 2021, 221, 112457. [Google Scholar] [CrossRef]
- Sprando, R.L.; Collins, T.F.; Black, T.N.; Olejnik, N.; Rorie, J.I.; Eppley, R.M.; Ruggles, D.I. Characterization of the effect of deoxynivalenol on selected male reproductive endpoints. Food Chem. Toxicol. 2005, 43, 623–635. [Google Scholar] [CrossRef]
- Jian, Y.Y.; Yong, F.Z.; Ai, M.L.; Xiang, F.K.; Yuan, X.L.; Kai, W.M.; Ai, H.J.; Shu, Y.F.; Xiao, L.Q. Toxic effects of T-2 toxin on reproductive system in male mice. Toxicol. Ind. Health 2010, 26, 25–31. [Google Scholar] [CrossRef]
- Granum, C.; Anchersen, S.; Karlsson, C.; Berg, V.; Olsaker, I.; Verhaegen, S.; Ropstad, E. Steroidogenic differential effects in neonatal porcine Leydig cells exposed to persistent organic pollutants derived from cod liver oil. Reprod. Toxicol. 2015, 11, 130–139. [Google Scholar] [CrossRef]
- Taroncher, M.; Rodríguez-Carrasco, Y.; Aspevik, T.; Kousoulaki, K.; Ruiz, M.-J. Cytoprotective effects of fish protein hydrolysates against H2O2-induced oxidative stress and mycotoxins in Caco-2/tc7 cells. Antioxidants 2021, 10, 975. [Google Scholar] [CrossRef]
- Wu, J.; Huang, W.; Xiao, H.; Xie, Y.; Yuan, Z.; Yi, J.; Chen, J.; Tu, D.; Tian, Y. Procyanidins B2 reverses the T-2 toxin-induced mitochondrial apoptosis in TM3 Leydig cells. J. Funct. Foods 2018, 45, 118–128. [Google Scholar] [CrossRef]
- Knigs, M.; Lenczyk, M.; Schwerdt, G.; Holzinger, H.; Humpf, H.U. Cytotoxicity, metabolism and cellular uptake of the mycotoxin deoxynivalenol in human proximal tubule cells and lung fibroblasts in primary culture. Toxicology 2007, 240, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Vejdovszky, K.; Warth, B.; Sulyok, M.; Marko, D. Non-synergistic cytotoxic effects of Fusarium and Alternaria toxin combinations in Caco-2 cells. Toxicol. Lett. 2016, 241, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, W.; Bian, X.; Yuan, Y.; Gu, J.; Liu, X.; Liu, Z.; Bian, J. Zearalenone induces apoptosis and cytoprotective autophagy in primary Leydig cells. Toxicol. Lett. 2014, 226, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Matias, F.B.; Yi, J.E.; Wu, J. T-2 toxin-induced cytotoxicity and damage on TM3 Leydig cells. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2016, 181, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, P.; Rong, J.; Ge, Y.; Shi, W. Small molecule CDS-3078 induces G2/M phase arrest and mitochondria-mediated apoptosis in HeLa cells. Exp. Ther. Med. 2020, 20, 284. [Google Scholar] [CrossRef]
- Ab Id-Essefi, S.; Baudrimont, I.; Hassen, W.; Ouanes, Z.; Mobio, T.A.; Anane, R.; Creppy, E.E.; Bacha, H. DNA fragmentation, apoptosis and cell cycle arrest induced by zearalenone in cultured DOK, Vero and Caco-2 cells: Prevention by Vitamin E. Toxicology 2003, 192, 237–248. [Google Scholar] [CrossRef]
- Liu, L.; Wang, D.; Li, L.; Ding, X.; Ma, H. Dehydroepiandrosterone inhibits cell proliferation and improves viability by regulating S phase and mitochondrial permeability in primary rat Leydig cells. Mol. Med. Rep. 2016, 14, 705–714. [Google Scholar] [CrossRef]
- Bartolowits, M.; Gast, J.; Hasler, A.J.; Cirrincione, A.M.; Davisson, V.J. Discovery of inhibitors for proliferating cell nuclear antigen using a computational-based linked-multiple-fragment screen. ACS Omega 2019, 4, 15181–15196. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Zhang, Y.; Su, J.; Yang, S.; Qiao, W.; Li, X.; Lei, Z.; Cheng, L.; An, N.; Wang, W. Effects of neuromedin B on steroidogenesis, cell proliferation, and apoptosis in porcine Leydig cell. J. Mol. Endocrinol. 2018, 61, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.; Chen, F.; Sun, J.; Zhou, J.; Wang, X.; Wang, N.; Li, X.; Zhang, Z.; Wang, A.; Jin, Y. Mycotoxin zearalenone induces apoptosis in mouse Leydig cells via an endoplasmic reticulum stress-dependent signalling pathway. Reprod. Toxicol. 2015, 52, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.F.; Yang, J.Y.; Li, Y.K.; Zhou, W. Toxicity and oxidative stress induced by t-2 toxin in cultured mouse Leydig cells. Toxicol. Mech. Methods 2016, 27, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Netro, H.M.; Chorfi, Y.; Price, C.A. Effects of the mycotoxin deoxynivalenol on steroidogenesis and apoptosis in granulosa cells. Reproduction 2015, 6, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, L.; Jiang, Y.; Zhu, L.; Xu, W.; Wang, X. Deoxynivalenol induces caspase-8-mediated apoptosis through the mitochondrial pathway in hippocampal nerve cells of piglet. Toxins 2021, 13, 73. [Google Scholar] [CrossRef]
- Wojtacha, P.; Trybowski, W.; Podlasz, P.; Migrodzka, M.; Obremski, K. Effects of a low dose of t-2 toxin on the percentage of T and B lymphocytes and cytokine secretion in the porcine ileal wall. Toxins 2021, 13, 277. [Google Scholar] [CrossRef]
- Oettel, M.; Mukhopadhyay, A.K. Progesterone: The forgotten hormone in men? Aging Male 2010, 7, 236–257. [Google Scholar] [CrossRef]
- Liu, Z.J.; Liu, Y.H.; Huang, S.Y.; Zang, Z.J. Insights into the regulation on proliferation and differentiation of stem Leydig cells. Stem Cell Rev. Rep. 2021, 17, 1521–1533. [Google Scholar] [CrossRef]
- Jian, Y.; Yong, F.; Na, N.A.; Wen, P.; Jian, F.; Xiang, P.; Xiao, L. Protective effects of l-arginine against testosterone synthesis decreased by t-2 toxin in mouse Leydig cells. Theriogenology 2019, 134, 98–103. [Google Scholar]
- Salva, D.A.; Klinefelter, G.R.; Hardy, M.P. Purification of rat Leydig cells: Increased yields after unit-gravity sedimentation of collagenase-dispersed interstitial cells. J. Androl. 2001, 22, 665–671. [Google Scholar]
- Lervik, S.; Krogh, K.V.; Karlsson, C.; Olsaker, I.; Andresen, Ø.; Dahl, E.; Verhaegen, S.; Ropstad, E. Steroidogenesis in primary cultures of neonatal porcine Leydig cells from Duroc and Norwegian Landrace breeds. Theriogenology 2011, 76, 1058–1069. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Dai, J.; Xu, J.; Yang, J.; Zhang, D. Comparative Cytotoxic Effects and Possible Mechanisms of Deoxynivalenol, Zearalenone and T-2 Toxin Exposure to Porcine Leydig Cells In Vitro. Toxins 2022, 14, 113. https://doi.org/10.3390/toxins14020113
Sun L, Dai J, Xu J, Yang J, Zhang D. Comparative Cytotoxic Effects and Possible Mechanisms of Deoxynivalenol, Zearalenone and T-2 Toxin Exposure to Porcine Leydig Cells In Vitro. Toxins. 2022; 14(2):113. https://doi.org/10.3390/toxins14020113
Chicago/Turabian StyleSun, Lingwei, Jianjun Dai, Jiehuan Xu, Junhua Yang, and Defu Zhang. 2022. "Comparative Cytotoxic Effects and Possible Mechanisms of Deoxynivalenol, Zearalenone and T-2 Toxin Exposure to Porcine Leydig Cells In Vitro" Toxins 14, no. 2: 113. https://doi.org/10.3390/toxins14020113
APA StyleSun, L., Dai, J., Xu, J., Yang, J., & Zhang, D. (2022). Comparative Cytotoxic Effects and Possible Mechanisms of Deoxynivalenol, Zearalenone and T-2 Toxin Exposure to Porcine Leydig Cells In Vitro. Toxins, 14(2), 113. https://doi.org/10.3390/toxins14020113