The Effect of Low Doses of Zearalenone (ZEN) on the Bone Marrow Microenvironment and Haematological Parameters of Blood Plasma in Pre-Pubertal Gilts

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
2.1. Experimental Feed
2.2. Clinical Observations
2.3. Concentrations of Zearalenone and Its Metabolites in the Bone Marrow Microenvironment
2.4. Carry-OVER Factor (CF)
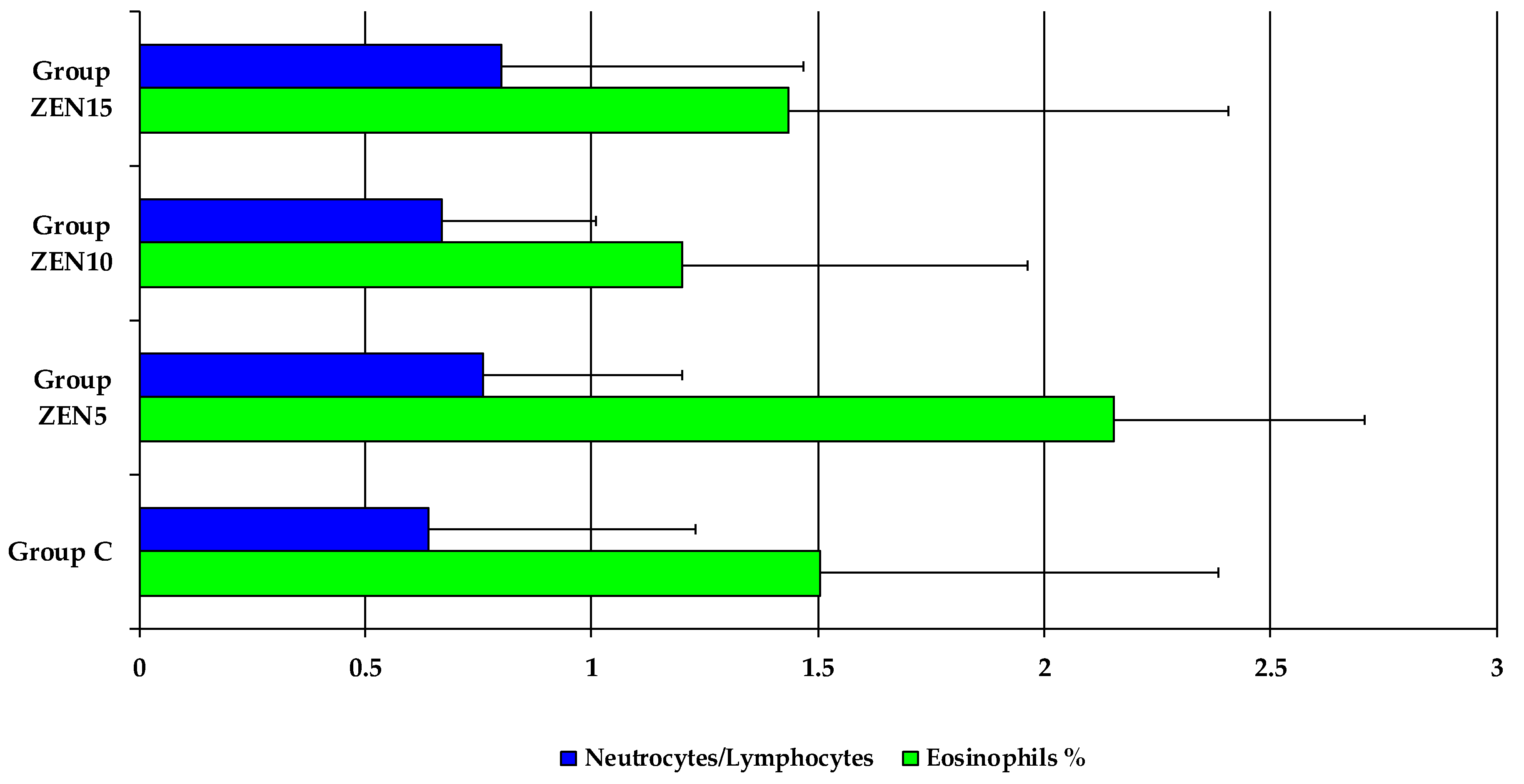
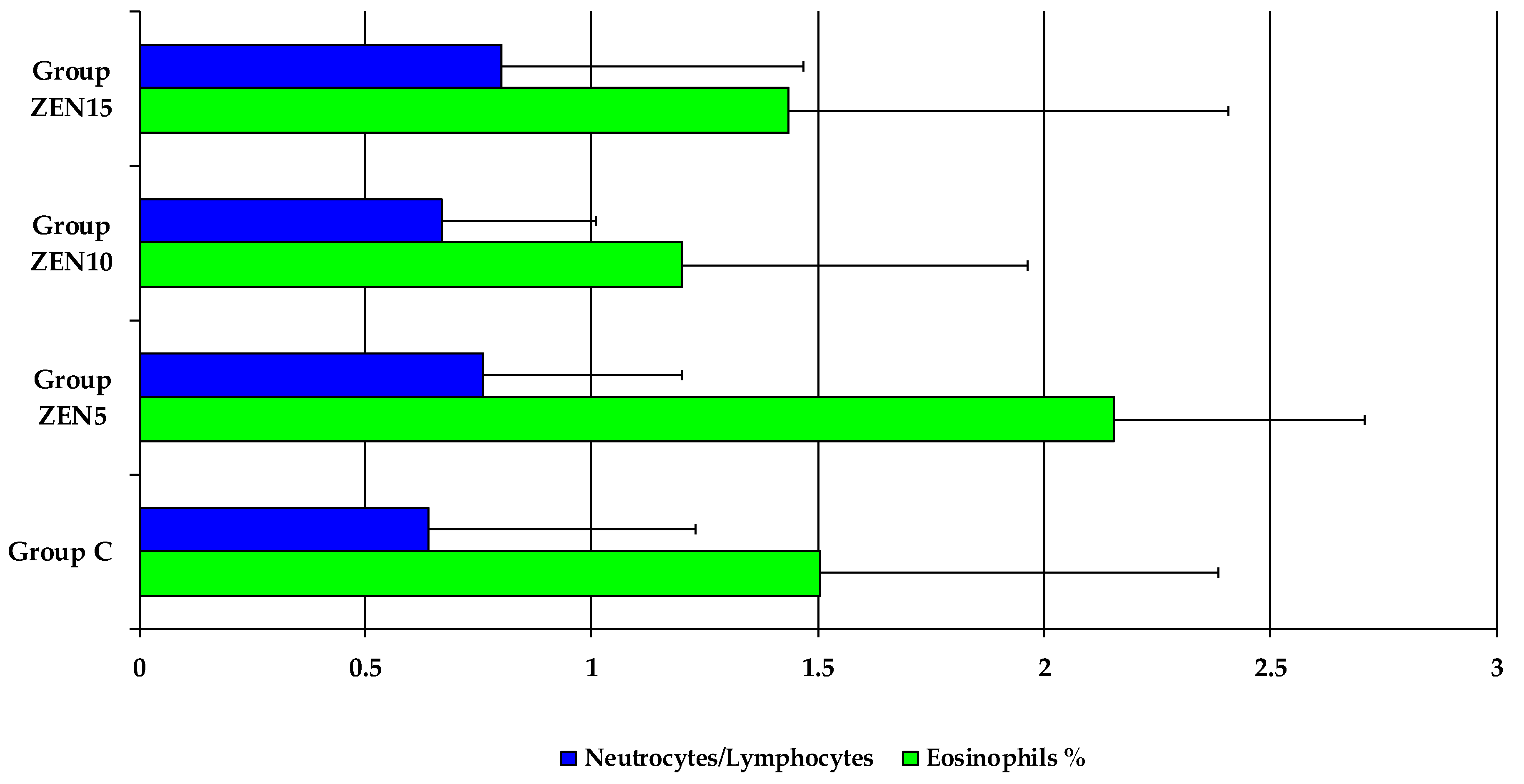
2.5. Results of Haematological Analyses
2.6. Haematological Analyses
2.6.1. General Analysis
2.6.2. Accompanying Factors
2.7. Correlation Coefficients
3. Discussion
3.1. Zearalenone and Its Metabolites in the Bone Marrow Microenvironment
3.2. White Blood Cells
3.3. Red Blood Cells
3.4. Platelets
3.5. Summary and Conclusions
4. Materials and Methods
4.1. General Information
4.2. Experimental Feed
Toxicological Analysis in Feed
4.3. Experimental Animals
4.3.1. Toxicological Analysis of Bone Marrow Microenvironment
Tissues Samples
Extraction Procedure
Chromatographic Quantification of ZEN and Its Metabolites
Carryover Factor
Statistical Analysis
4.4. Blood Sampling for Metabolic Profile Analysis
4.5. Haematology Tests
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wielogórska, E.; Elliott, C.T.; Danaher, M.; Connolly, L. Validation and application of a reporter gene assay for the determination of estrogenic endocrine disruptor activity in milk. Food Chem. Toxicol. 2014, 69, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ding, N.; Zhang, L.; Zhao, X.; Yang, Y.; Qu, H.; Fang, X. Biological Databases for Hematology Research. Genomics 2016, 14, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Flores-Flores, M.E.; Lizarraga, E.; de Cerain, A.L.; González-Peñas, E. Presence of mycotoxins in animal milk: A review. Food Control 2015, 53, 163–176. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; De Baere, S.; De Backer, P.; Croubels, S. Modified Fusarium mycotoxins unmasked: From occurrence in cereals to animal and human excretion. Food Chem. Toxicol. 2015, 80, 17–31. [Google Scholar] [CrossRef]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Piastowska-Ciesielska, A. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef]
- Dunbar, B.; Patel, M.; Fahey, J.; Wira, C. Endocrine control of mucosal immunity in the female reproductive tract: Impact of environmental disruptors. Mol. Cell. Endocrinol. 2012, 354, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Rykaczewska, A.; Gajęcka, M.; Dąbrowski, M.; Wiśniewska, A.; Szcześniewska, J.; Gajęcki, M.T.; Zielonka, Ł. Growth performance, selected blood biochemical parameters and body weights of pre-pubertal gilts fed diets supplemented with different doses of zearalenone (ZEN). Toxicon 2018, 152, 84–94. [Google Scholar] [CrossRef]
- Zachariasova, M.; Dzumana, Z.; Veprikova, Z.; Hajkovaa, K.; Jiru, M.; Vaclavikova, M.; Zachariasova, A.; Pospichalova, M.; Florian, M.; Hajslova, J. Occurrence of multiple mycotoxins in European feeding stuffs, assessment of dietary intake by farm animals. Anim. Feed Sci. Technol. 2014, 193, 124–140. [Google Scholar] [CrossRef]
- Frizzell, C.; Ndossi, D.; Verhaegen, S.; Dahl, E.; Eriksen, G.; Sørlie, M.; Ropstad, E.; Muller, M.; Elliott, C.T.; Connolly, L. Endocrine disrupting effects of zearalenone, alpha- and beta-zearalenol at the level of nuclear receptor binding and steroidogenesis. Toxicol. Lett. 2011, 206, 210–217. [Google Scholar] [CrossRef]
- Gajęcka, M.; Tarasiuk, M.; Zielonka, Ł.; Dąbrowski, M.; Gajęcki, M. Risk assessment for changes in metabolic profile and body weight of pre-pubertal gilts during long-term monotonic exposure to low doses of zearalenone (ZEN). Res. Vet. Sci. 2016, 109, 169–180. [Google Scholar] [CrossRef]
- Zielonka, Ł.; Waśkiewicz, A.; Beszterda, M.; Kostecki, M.; Dąbrowski, M.; Obremski, K.; Goliński, P.; Gajęcki, M. Zearalenone in the Intestinal Tissues of Immature Gilts Exposed per os to Mycotoxins. Toxins 2015, 7, 3210–3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, 4851. [Google Scholar] [CrossRef] [Green Version]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. Activity of zearalenone in the porcine intestinal tract. Molecules 2017, 22, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, E.J. Paradigm lost, paradigm found: The re-emergence of hormesis as a fundamental dose response model in the toxicological sciences. Environ. Pollut. 2005, 138, 378–411. [Google Scholar] [CrossRef] [PubMed]
- Freire, L.; Sant’Ana, A.S. Modified mycotoxins: An updated review on their formation, detection, occurrence, and toxic effects. Food Chem. Toxicol. 2018, 111, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Cieplińska, K.; Gajęcka, M.; Nowak, A.; Dąbrowski, M.; Zielonka, Ł.; Gajęcki, M.T. The gentoxicity of caecal water in gilts exposed to low doses of zearalenone. Toxins 2018, 10, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.-H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endoc. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajęcka, M.; Majewski, M.S.; Zielonka, Ł.; Grzegorzewski, W.; Onyszek, E.; Lisieska-Zołnierczyk, S.; Juśkiewicz, J.; Babuchowski, A.; Gajęcki, M.T. Concentration of Zearalenone, Alpha-Zearalenol and Beta-Zearalenol in the Myocardium and the Results of Isometric Analyses of the Coronary Artery in Prepubertal Gilts. Toxins 2021, 13, 396. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the risks for public health related to the presence of zearalenone in food. EFSA J. 2011, 9, 2197. [Google Scholar] [CrossRef]
- Pastoor, T.P.; Bachman, A.N.; Bell, D.R.; Cohen, S.M.; Dellarco, M.; Dewhurst, I.C.; Doe, J.E.; Doerrer, N.G.; Embry, M.R.; Hines, R.N.; et al. A 21st century roadmap for human health risk assessment. Crit. Rev. Toxicol. 2014, 44, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.S.; Goyal, N. Estrogens as regulator of hematopoietic stem cell, immune cells and bone biology. Life Sci. 2021, 269, 119091. [Google Scholar] [CrossRef] [PubMed]
- Hamza, E.; Metzinger, L.; Metzinger-Le Meuth, V. Uremic Toxins Affect Erythropoiesis during the Course of Chronic Kidney Disease: A Review. Cells 2020, 9, 2039. [Google Scholar] [CrossRef]
- Molica, F.; Stierlin, F.B.; Fontana, P.; Kwak, B.R. Pannexin- and Connexin-Mediated Intercellular Communication in Platelet Function. Int. J. Mol. Sci. 2017, 18, 850. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.B.; Zhang, J.; Lucas, D. The Role of the Bone Marrow Microenvironment in the Response to Infection. Front. Immunol. 2020, 11, 585402. [Google Scholar] [CrossRef]
- Tjon, J.M.-L.; Langemeijer, S.M.C.; Halkes, C.J.M. Anti Thymocyte Globulin-Based Treatment for Acquired Bone Marrow Failure in Adults. Cells 2021, 10, 2905. [Google Scholar] [CrossRef] [PubMed]
- Yu, V.W.C.; Scadden, D.T. Heterogeneity of the bone marrow niche. Curr. Opin. Hematol. 2016, 23, 331–338. [Google Scholar] [CrossRef]
- Rykaczewska, A.; Gajęcka, M.; Onyszek, E.; Cieplińska, K.; Dąbrowski, M.; Lisieska-Żołnierczyk, S.; Bulińska, M.; Babuchowski, A.; Gajęcki, M.T.; Zielonka, Ł. Imbalance in the Blood Concentrations of Selected Steroids in Prepubertal Gilts Depending on the Time of Exposure to Low Doses of Zearalenone. Toxins 2019, 11, 561. [Google Scholar] [CrossRef] [Green Version]
- Cieplińska, K.; Gajęcka, M.; Dąbrowski, M.; Rykaczewska, A.; Zielonka, Ł.; Lisieska-Żołnierczyk, S.; Bulińska, M.; Gajęcki, M.T. Time-dependent changes in the intestinal microbiome of gilts exposed to low zearalenone doses. Toxins 2019, 11, 296. [Google Scholar] [CrossRef] [Green Version]
- Gajęcka, M.; Tarasiuk, M.; Zielonka, Ł.; Dąbrowski, M.; Nicpoń, J.; Baranowski, M.; Gajęcki, M.T. Changes in the metabolic profile and body weight of pre-pubertal gilts during prolonged monotonic exposure to low doses of zearalenone and deoxynivalenol. Toxicon 2017, 125, 32–43. [Google Scholar] [CrossRef]
- Etim, N.N.; Williams, M.E.; Akpabio, U.; Offiong, E.E. Haematological parameters and factors affecting their values. Agric. Sci. 2014, 2, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Goyarts, T.; Dänicke, S.; Brüssow, K.P.; Valenta, H.; Ueberschär, K.H.; Tiemann, U. On the transfer of the Fusarium toxins deoxynivalenol (DON) and zearalenone (ZON) from sows to their fetuses during days 35–70 of gestation. Toxicol. Lett. 2007, 171, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Winnicka, A. Reference Values of Basic Laboratory Tests in Veterinary Medicine; SGGW Publishers: Warszawa, Poland, 2008; ISBN 978-83-7244-974-0. [Google Scholar]
- Li, Q.; Patience, J.F. Factors involved in the regulation of feed and energy intake of pigs. Anim. Feed Sci. Technol. 2016, 233, 22–33. [Google Scholar] [CrossRef]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Lin, K.S.; Uemura, S.; Thwin, K.K.M.; Nakatani, N.; Ishida, T.; Yamamoto, N.; Tamura, A.; Saito, A.; Mori, T.; Hasegawa, D.; et al. Minimal residual disease in high-risk neuroblastoma shows a dynamic and disease burden-dependent correlation between bone marrow and peripheral blood. Transl. Oncol. 2021, 14, 101019. [Google Scholar] [CrossRef]
- Benagiano, M.; Bianchi, P.; D’Elios, M.M.; Brosens, I.; Benagiano, G. Autoimmune diseases: Role of steroid hormones. Best Pr. Res. Clin. Obstet. Gynaecol. 2019, 60, 24–34. [Google Scholar] [CrossRef]
- Wilkenfeld, S.R.; Linc, C.; Frigo, D.E. Communication between genomic and non-genomic signaling events coordinate steroid hormone actions. Steroids 2018, 133, 2–7. [Google Scholar] [CrossRef]
- Gajęcka, M.; Otrocka-Domagała, I. Immunocytochemical expression of 3β- and 17β-hydroxysteroid dehydrogenase in bitch ovaries exposed to low doses of zearalenone. Pol. J. Vet. Sci. 2013, 16, 55–62. [Google Scholar] [CrossRef]
- Lawrenz, B.; Melado, L.; Fatemi, H. Premature progesterone rise in ART-cycles. Reprod. Biol. 2018, 18, 1–4. [Google Scholar] [CrossRef]
- Nakada, D.; Oguro, H.; Levi, B.P.; Ryan, N.; Kitano, A.; Saitoh, Y.; Takeichi, M.; Wendt, G.R.; Morrison, S.J. Estrogen increases haematopoietic stem cell self-renewal in females and during pregnancy. Nature 2014, 505, 555–558. [Google Scholar] [CrossRef]
- Balla, B.; Sárvári, M.; Kósa, J.P.; Kocsis-Deák, B.; Tobiás, B.; Árvai, K.; Takács, I.; Podani, J.; Liposits, Z.; Lakatos, P. Long-term selective estrogen receptor-beta agonist treatment modulates gene expression in bone and bone marrow of ovariectomized rats. J. Steroid Biochem. 2019, 188, 185–194. [Google Scholar] [CrossRef]
- Pinkas, J.; Gujski, M.; Wierzbińska-Stępniak, A.; Owoc, A.; Bojar, I. The polymorphism of oestrogen receptor alpha is important for metabolic consequences associated with menopause. Endokrynol. Polska 2016, 67, 608–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satirapod, C.; Wang, N.; MacDonald, J.A.; Sun, M.; Woods, D.C.; Tilly, J.L. Estrogen regulation of germline stem cell differentiation as a mechanism contributing to female reproductive aging. Aging 2020, 12, 7313–7333. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Feng, N.; Wang, Y.; Noll, L.; Xu, S.; Liu, X.; Lu, N.; Zou, H.; Gu, J.; Yuan, Y.; et al. Effects of zearalenone and its derivatives on the synthesis and secretion of mammalian sex steroid hormones: A review. Food Chem. Toxicol. 2019, 126, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Jiang, T.; Lin, P.; Chen, H.; Wang, L.; Wang, N.; Zhao, F.; Tang, K.; Zhou, D.; Wang, A.; et al. Apoptosis inducing factor gene depletion inhibits zearalenone-induced cell death in a goat Leydig cell line. Reprod. Toxicol. 2017, 67, 129–139. [Google Scholar] [CrossRef]
- Hennig-Pauka, I.; Koch, F.J.; Schaumberger, S.; Woechtl, B.; Novak, J.; Sulyok, M.; Nagl, V. Current challenges in the diagnosis of zearalenone toxicosis as illustrated by a field case of hyperestrogenism in suckling piglets. Porc. Health Manag. 2018, 4, 18. [Google Scholar] [CrossRef]
- Fañanas-Baquero, S.; Orman, I.; Aparicio, F.B.; de Miguel, S.B.; Merino, J.G.; Yañez, R.; Sainz, Y.F.; Sánchez, R.; Dessy-Rodríguez, M.; Alberquilla, O.; et al. Natural estrogens enhance the engraftment of human hematopoietic stem and progenitor cells in immunodeficient mice. Haematologica 2021, 106, 1659–1670. [Google Scholar] [CrossRef]
- Luisetto, M.; Almukhtar, N.; Ahmadabadi, B.N.; Hamid, G.A.; Mashori, G.R.; Khan, K.R.; Khan, F.A.; Cabianca, L. Endogenus Toxicology: Modern Physio- Pathological Aspects and Relationship with New Therapeutic Strategies. An Integrative Discipline Incorporating Concepts from Different Research Discipline Like Biochemistry, Pharmacology and Toxicology. Arch Cancer Sci Ther. 2019, 3, 001–004. [Google Scholar] [CrossRef] [Green Version]
- Elegido, A.; Graell, M.; Andrés, P.; Gheorghe, A.; Marcos, A.; Nova, E. Increased naive CD4+ and B lymphocyte subsets are associated with body mass loss and drive relative lymphocytosis in anorexia nervosa patients. Nutr. Res. 2017, 39, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Deng, H.; Deng, Y.; Liang, Z.; Deng, J.; Zuo, Z.; Hu, Y.; Shen, L.; Yu, S.; Cao, S. Combined effects of deoxynivalenol and zearalenone on oxidative injury and apoptosis in porcine splenic lymphocytes in vitro. Exp. Toxicol. Pathol. 2017, 69, 612–617. [Google Scholar] [CrossRef]
- Etim, N.A.N.; Offiong, E.E.A.; Williams, M.E.; Asuquo, L.E. Influence of nutrition on blood parameters of pigs. Am. J. Biol. Life Sci. 2014, 2, 46–52. Available online: http://www.openscienceonline.com/journal/ajbls (accessed on 10 January 2022).
- Sutherland, M.A.; Bryer, P.J.; Krebs, N.; McGlone, J.J. Tail docking in pigs: Acute physiological and behavioural responses. Animal 2008, 2, 292–297. [Google Scholar] [CrossRef] [Green Version]
- Gajęcki, M.; Gajęcka, M.; Zielonka, Ł.; Jakimiuk, E.; Obremski, K. Zearalenone as a potential allergen in the alimentary tract—A review. Pol. J. Food Nutr. Sci. 2006, 56, 263–268. [Google Scholar]
- Gajęcka, M.; Przybylska-Gornowicz, B. The low doses effect of experimental zearalenone (ZEN) intoxication on the presence of Ca2+ in selected ovarian cells from pre-pubertal bitches. Pol. J. Vet. Sci. 2012, 15, 711–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przybylska-Gornowicz, B.; Tarasiuk, M.; Lewczuk, B.; Prusik, M.; Ziółkowska, N.; Zielonka, Ł.; Gajęcki, M.; Gajęcka, M. The Effects of Low Doses of Two Fusarium Toxins, Zearalenone and Deoxynivalenol, on the Pig Jejunum. A Light and Electron Microscopic Study. Toxins 2015, 7, 4684–4705. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Wan, M.; Gu, Y.; Song, Y.; Zhang, Q.; Liu, L.; Meng, H.; Xia, Y.; Shi, H.B.; Su, Q.; et al. Red cell distribution width is associated with hemoglobin A1C elevation, but not glucose elevation. J. Diabetes Complicat. 2017, 31, 1544–1548. [Google Scholar] [CrossRef]
- Paiva-Martins, F.; Barbosa, S.; Silva, M.; Monteiro, D.; Pinheiro, V.; Mourão, J.L.; Fernandes, J.; Rocha, S.; Belo, L.; Santos-Silva, A. The effect of olive leaf supplementation on the constituents of blood and oxidative stability of red blood cells. J. Funct. Foods 2014, 9, 271–279. [Google Scholar] [CrossRef]
- Tatay, E.; Espín, S.; García-Fernández, A.J.; Ruiz, M.J. Oxidative damage and disturbance of antioxidant capacity by zearalenone and its metabolites in human cells. Toxicol. Vitro 2017, 45, 334–339. [Google Scholar] [CrossRef]
- Jilani, K.; Lang, F. Ca(2+)-dependent suicidal erythrocyte death following zearalenone exposure. Arch. Toxicol. 2013, 87, 1821–1828. [Google Scholar] [CrossRef]
- Pyrshev, K.A.; Klymchenko, A.S.; Csúcs, G.; Demchenko, A.P. Apoptosis and eryptosis: Striking differences on biomembrane level. BBA-Biomembrane 2018, 1860, 1362–1371. [Google Scholar] [CrossRef]
- Qadri, S.M.; Bissinger, R.; Solh, Z.; Oldenborg, P.A. Eryptosis in health and disease: A paradigm shift towards understanding the (patho)physiological implications of programmed cell death of erythrocytes. Blood Rev. 2017, 31, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Lang, E.; Lang, F. Mechanisms and pathophysiological significance of eryptosis, the suicidal erythrocyte death. Semin. Cell Dev. Biol. 2015, 39, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.Y.; Lau, P.M.; Yeung, K.W.; Kong, S.K. The second generation tyrosine kinase inhibitor dasatinib induced eryptosis in human erythrocytes-An in vitro study. Toxicol. Lett. 2018, 295, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Rybarczyk, L.; Jakimiuk, E.; Zielonka, Ł.; Obremski, K.; Zwierzchowski, W.; Gajęcki, M. The effect of experimental long-term exposure to low-dose zearalenone on uterine histology in sexually immature gilts. Exp. Toxicol. Pathol. 2012, 64, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Gajęcka, M.; Zielonka, Ł.; Dąbrowski, M.; Mróz, M.; Gajęcki, M. The effect of low doses of zearalenone and its metabolites on progesterone and 17β-estradiol concentrations in peripheral blood and body weights of pre-pubertal female Beagle dogs. Toxicon 2013, 76, 260–269. [Google Scholar] [CrossRef]
- Yorbik, O.; Mutlu, C.; Tanju, I.A.; Celik, D.; Ozcan, O. Mean platelet volume in children with attention deficit hyperactivity disorder. Med. Hypotheses 2014, 82, 341–345. [Google Scholar] [CrossRef]
- Dai, Q.; Zhang, G.; Lai, C.; Du, Z.; Chen, L.; Chen, Q.; Peng, L.; Wang, Y.; Yang, H.; Ye, L.; et al. Two cases of false platelet clumps flagged by the automated haematology analyser Sysmex XE-2100. Clin. Chim. Acta. 2014, 429, 152–156. [Google Scholar] [CrossRef]
- Meyer, J.; Lejmi, E.; Fontana, P.; Morel, P.; Gonelle-Gispert, C.; Bühler, L. A focus on the role of platelets in liver regeneration: Do platelet-endothelial cell interactions initiate the regenerative process? J. Hepatol. 2015, 63, 1263–1271. [Google Scholar] [CrossRef]
- Wysokiński, A.; Szczepocka, E. Platelet parameters (PLT, MPV, P-LCR) in patients with schizophrenia, unipolar depression and bipolar disorder. Psychiatry Res. 2016, 237, 238–245. [Google Scholar] [CrossRef]
- Heberer, T.; Lahrssen-Wiederholt, M.; Schafft, H.; Abraham, K.; Pzyrembel, H.; Henning, K.J.; Schauzu, M.; Braeunig, J.; Goetz, M.; Niemann, L.; et al. Zero tolerances in food and animal feed-Are there any scientific alternatives? A European point of view on an international controversy. Toxicol. Lett. 2007, 175, 118–135. [Google Scholar] [CrossRef]
- Smith, D.; Combes, R.; Depelchin, O.; Jacobsen, S.D.; Hack, R.; Luft, J.; Lammens, L.; von Landenberg, F.; Phillips, B.; Pfister, R.; et al. Optimising the design of preliminary toxicity studies for pharmaceutical safety testing in the dog. Regul. Toxicol. Pharm. 2005, 41, 95–101. [Google Scholar] [CrossRef]
- Meerpoel, C.; Vidal, A.; Tangni, E.K.; Huybrechts, B.; Couck, L.; De Rycke, R.; De Bels, L.; De Saeger, S.; Van den Broeck, W.; Devreese, M.; et al. A Study of Carry-Over and Histopathological Effects after Chronic Dietary Intake of Citrinin in Pigs, Broiler Chickens and Laying Hens. Toxins 2020, 12, 719. [Google Scholar] [CrossRef]
- Kowalski, A.; Kaleczyc, J.; Gajęcki, M.; Zieliński, H. Adrenaline, noradrenaline and cortisol levels in pigs during blood collection (In Polish). Med. Weter. 1996, 52, 716–718. [Google Scholar]
- Williams, M.S.; Ebel, E.D. Estimating correlation of prevalence at two locations in the farm-to-table continuum using qualitative test data. Int. J. Food Microbiol. 2017, 245, 29–37. [Google Scholar] [CrossRef]
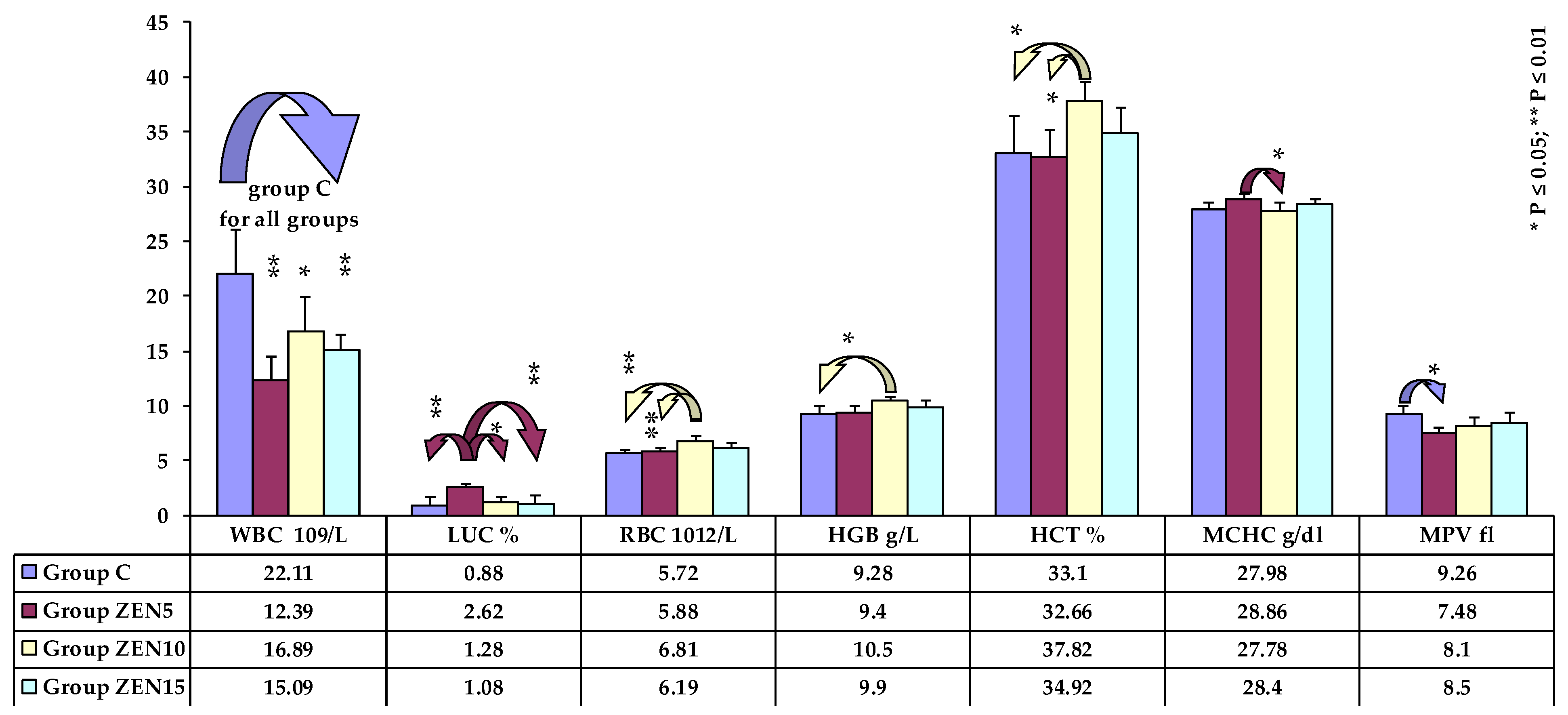


{kind=link}
{kind=link}
{kind=link}
| Exposure Date | Feed Intake [kg/Day] | Total ZEN Doses in Groups Respectively [µg/kg BW] | Group ZEN5 [ng/g] | CF Group ZEN5 | Group ZEN10 [ng/g] | CF Group ZEN10 | Group ZEN15 [ng/g] | CF Group ZEN15 |
|---|---|---|---|---|---|---|---|---|
| Zearalenone | ||||||||
| D1 | 0.8 | 80.5/161.9/242.7 | 5.64 ± 3.24 | 7 × 10−5 | 6.35 ± 3.56 | 39 × 10−6 | 8.17 ± 0.45 | 33 × 10−6 |
| D2 | 1.1 | 101.01/196.9/298.2 | 4.69 ± 4.28 | 46 × 10−6 | 7.08 ± 0.10 | 35 × 10−6 | 6.84 ± 0.18 ●● | 22 × 10−6 |
| D3 | 1.6 | 128.3/481.4/716.7 | 7.74 ± 0.26 | 6 × 10−5 | 7.35 ± 0.25 * | 15 × 10−6 | 7.03 ± 0.14 **,●● | 98 × 10−7 |
| α-ZEL and β-ZEL | ||||||||
| D1–D3 | not applicable | 0 | ||||||
| Blood Collection Dates | WBC 109/L | MONO % | LUC % | MPV fl |
|---|---|---|---|---|
| 1 | 15.52 ± 7.58 | 5.32 ± 1.06 | 0.88 ± 0.63 bb | 7.36 ± 1.61 cc,ff |
| 2 | 18.8 ± 4.84 | 4.94 ± 1.07 | 1.72 ± 0.99 | 8.94 ± 0.99 a |
| 3 | 18.8 ± 2.71 | 6.04 ± 3.69 | 1.42 ± 0.94 | 7.68 ± 0.55 c,f |
| 4 | 12.39 ± 2.1 ee | 4.3 ± 1.02 | 2.62 ± 0.31 | 7.48 ± 0.54 c,ff |
| 5 | 17.23 ± 0.57 | 3.56 ± 0.55 | 1.26 ± 0.38 | 9.44 ± 1.44 |
| 6 | 15.45 ± 1.41 | 6.78 ± 0.91 | 0.84 ± 0.76 bb | 8.14 ± 0.59 |
| 7 | 12.46 ± 1.37 ee | 3.84 ± 0.89 | 1.44 ± 0.73 | 8.44 ± 0.87 |
| 8 | 15.04 ± 10.37 | 6.4 ± 2.59 | 0.8 ± 0.53 b | 8.82 ± 1.42 |
| 9 | 24.55 ± 5.39 | 5.78 ± 1.18 | 0.6 ± 0.35 bb | 8.82 ± 1.35 |
| 10 | 10.44 ± 3 ee | 2.82 ± 0.84 d | 0.72 ± 0.37 bb | 9.66 ± 1.88 |
| Blood Collection Dates | WBC 109/L | NEUT % | LYMPH % | LUC % | RBC 1012/L | HGB g/L | HCT % | MCV fl | MCHC g/dL | HDW g/dL |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 21.86 ± 2.67 d | 37.68 ± 3.89 | 55.7 ± 3.89 | 0.8 ± 0.64 | 6.68 ± 0.54 | 10.79 ± 0.65 | 37.08 ± 2.19 | 55.6 ± 1.81 | 29.1 ± 1.23 | 1.58 ± 0.13 aa,bb |
| 2 | 22.74 ± 3.6 a,d | 43.7 ± 12.26 a | 50.02 ± 12.12 a | 0.86 ± 0.69 | 6.57 ± 0.38 c | 10.22 ± 0.36 b,cc | 34.98 ± 1.27 bb,cc | 53.3 ± 2.52 | 29.12 ± 0.57 a | 1.57 ± 0.04 a,b |
| 3 | 19.55 ± 3.03 | 45.14 ± 5.14 a | 46.94 ± 3.13 aa | 0.98 ± 0.62 | 6.56 ± 0.17 c | 10.18 ± 0.34 b,cc | 35.48 ± 0.74 bb,cc | 54.14 ± 2.49 | 28.7 ± 0.51 | 1.6 ± 0.08 a,bb |
| 4 | 16.89 ± 2.94 | 29.58 ± 7.88 | 63.9 ± 7.68 | 1.28 ± 0.37 | 6.81 ± 0.39 | 10.5 ± 0.36 | 37.82 ± 1.63 | 55.6 ± 1.75 | 27.78 ± 0.8 | 1.54 ± 0.09 a |
| 5 | 22.6 ± 6.49 a,d | 43.34 ± 11.1 | 48.62 ± 10.43 a | 1.86 ± 0.34 c | 6.83 ± 0.46 | 10.28 ± 0.76 b,c | 35.7 ± 2.87 bb,cc | 52.22 ± 1.6 a | 28.72 ± 0.38 | 1.54 ± 0.05 a |
| 6 | 15.36 ± 3.68 | 33.7 ± 6.15 | 57.92 ± 6.89 | 1.22 ± 0.61 | 6.71 ± 0.33 | 10.62 ± 0.97 | 38.06 ± 2.54 | 56.68 ± 1.98 | 27.86 ± 0.86 | 1.55 ± 0.07 a,b |
| 7 | 13.59 ± 2.68 | 27.28 ± 8.23 | 66.96 ± 8.07 | 0.66 ± 0.45 | 6.51 ± 0.42 b,c | 10.34 ± 0.28 c | 37.56 ± 1.11 | 57.86 ± 2.94 | 27.48 ± 0.54 | 1.43 ± 0.06 |
| 8 | 15.43 ± 5.68 | 34.08 ± 5.43 | 58.78 ± 5.47 | 1.64 ± 0.01 | 7.35 ± 0.33 | 11.54 ± 0.6 | 40.54 ± 2.56 | 55.18 ± 3.32 | 28.44 ± 0.6 | 1.44 ± 0.05 |
| 9 | 19.81 ± 3.64 | 39.6 ± 5.69 | 53.58 ± 6.26 | 0.52 ± 0.15 | 7.47 ± 0.41 | 11.7 ± 0.43 | 40.92 ± 0.86 | 54.84 ± 2.3 | 28.54 ± 0.94 | 1.53 ± 0.06 a |
| 10 | 12.85 ± 3.75 | 40.4 ± 6.31 | 53.52 ± 6.83 | 0.88 ± 0.21 | 6.83 ± 0.4 | 10.7 ± 0.55 | 37.34 ± 1.23 | 54.74 ± 2.65 | 28.62 ± 0.79 | 1.54 ± 0.08 a |
| Blood Collection Dates | WBC 109/L | NEUT % | LYMPH % | EOS % | MPV fl |
|---|---|---|---|---|---|
| 1 | 22.01 ± 3.85 ee,ff | 52.92 ± 7.91 | 39.96 ± 8.22 e | 2.4 ± 1.47 | 8.22 ± 1.65 |
| 2 | 19.83 ± 1.35 e,f | 43.7 ± 6.64 | 48.62 ± 5.62 | 1.32 ± 0.52 | 7.68 ± 1.55 a |
| 3 | 20.73 ± 2.21 e,f | 44.92 ± 7.66 | 46.42 ± 6.85 | 1.96 ± 0.65 | 7.50 ± 0.45 |
| 4 | 15.09 ± 14.43 | 38.54 ± 7.92 | 55.38 ± 7.23 | 0.64 ± 0.27 a | 8.51 ± 1.12 |
| 5 | 22.33 ± 5.96 ee,ff | 48.16 ± 12.12 | 45.62 ± 10.12 | 1.22 ± 0.43 | 7.58 ± 1.05 aa,b,c |
| 6 | 16.66 ± 3.96 | 37 ± 5.58 | 54.14 ± 5.95 | 1.6 ± 0.41 | 7.84 ± 1.09 |
| 7 | 10.79 ± 2.53 | 26.44 ± 6.28 a,d | 66.12 ± 4.45 | 0.74 ± 0.23 a | 7.44 ± 1.32 |
| 8 | 19.04 ± 2.47 | 41.32 ± 11.48 | 49.64 ± 11.23 | 1.9 ± 0.76 | 8.38 ± 2.72 |
| 9 | 14.8 ± 10.25 | 29.02 ± 20.07 a | 40.37 ± 27.41 e | 1.15 ± 0.82 | 8.85 ± 2.46 |
| 10 | 10.73 ± 1.78 | 39.68 ± 6.79 | 53.94 ± 7.3 | 1.44 ± 1.05 | 8.28 ± 1.05 aa,b,cc |
| Blood Collection Dates | PLT 109/L Group ZEN5 | PLT 109/L Group ZEN10 | PLT Clumps % Group ZEN10 | PLT 109/L Group ZEN15 | PLT Clumps % Group ZEN15 |
|---|---|---|---|---|---|
| 1 | 405.4 ± 295.39 | 690.2 ± 121.87 e | 26.4 ± 14.75 | 479.8 ± 214.68 | 19.8 ± 18.07 |
| 2 | 522.6 ± 112.81 | 763 ± 107.01 d,ee | 19.8 ± 18.07 | 668 ± 144.82 d,e | 19.8 ± 18.07 |
| 3 | 745 ± 54.41 e | 813.2 ± 149.05 c,dd,ee | 0 ± 0 | 753.8 ± 167.69 a,dd,ee | 0 ± 0 |
| 4 | 741.4 ± 89.6 e | 734.6 ± 131.75 d,e | 26.4 ± 14.75 | 501.6 ± 62.13 b | 33 ± 0 b |
| 5 | 517.2 ± 162.41 | 800.2 ± 212.97 c,dd,ee | 33 ± 0 bb | 718.2 ± 129.56 a,d,ee | 26.4 ± 14.75 |
| 6 | 645.2 ± 140.47 | 650.6 ± 238.35 | 19.8 ± 18.07 | 635.4 ± 38.01 e | 19.8 ± 18.07 |
| 7 | 463.6 ± 83.09 | 569.4 ± 132.14 | 26.4 ± 14.75 | 607.8 ± 203.15 | 13.2 ± 18.07 |
| 8 | 454 ± 317.27 | 617.2 ± 180.88 | 13.2 ± 18.07 | 626.6 ± 263.92 e | 19.8 ± 18.07 |
| 9 | 595.6 ± 172.33 | 497.2 ± 101.9 | 33 ± 0 bb | 394 ± 294.02 | 24.75 ± 16.5 |
| 10 | 369.4 ± 118.85 | 478.2 ± 151.9 | 33 ± 0 bb | 386.6 ± 158.7 | 33 ± 0 b |
| Exposure Date | Experimental Groups | PLT | WBC | RBC |
|---|---|---|---|---|
| D1 | Group ZEN5 | −0.739 | −0.362 | −0.456 |
| Group ZEN10 | 0.685 | −0.502 | −0.189 | |
| Group ZEN15 | −0.590 | −0.336 | −0.869 | |
| D2 | Group ZEN5 | −0.251 | −0.851 | −0.424 |
| Group ZEN10 | −0.559 | −0.325 | −0.832 | |
| Group ZEN15 | −0.682 | −0.797 | −0.565 | |
| D3 | Group ZEN5 | −0.772 | −0.230 | −0.752 |
| Group ZEN10 | −0.731 | −0.451 | −0.300 | |
| Group ZEN15 | −0.436 | −0.128 | −0.738 |
| Parameters | Composition Declared by the Manufacturer (%) |
|---|---|
| Soybean meal | 16 |
| Wheat | 55 |
| Barley | 22 |
| Wheat bran | 4.0 |
| Chalk | 0.3 |
| Zitrosan | 0.2 |
| Vitamin-mineral premix 1 | 2.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mróz, M.; Gajęcka, M.; Przybyłowicz, K.E.; Sawicki, T.; Lisieska-Żołnierczyk, S.; Zielonka, Ł.; Gajęcki, M.T. The Effect of Low Doses of Zearalenone (ZEN) on the Bone Marrow Microenvironment and Haematological Parameters of Blood Plasma in Pre-Pubertal Gilts. Toxins 2022, 14, 105. https://doi.org/10.3390/toxins14020105
Mróz M, Gajęcka M, Przybyłowicz KE, Sawicki T, Lisieska-Żołnierczyk S, Zielonka Ł, Gajęcki MT. The Effect of Low Doses of Zearalenone (ZEN) on the Bone Marrow Microenvironment and Haematological Parameters of Blood Plasma in Pre-Pubertal Gilts. Toxins. 2022; 14(2):105. https://doi.org/10.3390/toxins14020105
Chicago/Turabian StyleMróz, Magdalena, Magdalena Gajęcka, Katarzyna E. Przybyłowicz, Tomasz Sawicki, Sylwia Lisieska-Żołnierczyk, Łukasz Zielonka, and Maciej Tadeusz Gajęcki. 2022. "The Effect of Low Doses of Zearalenone (ZEN) on the Bone Marrow Microenvironment and Haematological Parameters of Blood Plasma in Pre-Pubertal Gilts" Toxins 14, no. 2: 105. https://doi.org/10.3390/toxins14020105
APA StyleMróz, M., Gajęcka, M., Przybyłowicz, K. E., Sawicki, T., Lisieska-Żołnierczyk, S., Zielonka, Ł., & Gajęcki, M. T. (2022). The Effect of Low Doses of Zearalenone (ZEN) on the Bone Marrow Microenvironment and Haematological Parameters of Blood Plasma in Pre-Pubertal Gilts. Toxins, 14(2), 105. https://doi.org/10.3390/toxins14020105