Prevalence of Fusarium fungi and Deoxynivalenol Levels in Winter Wheat Grain in Different Climatic Regions of Poland

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Culture-Based Morphological Method for the Identification of Fungi
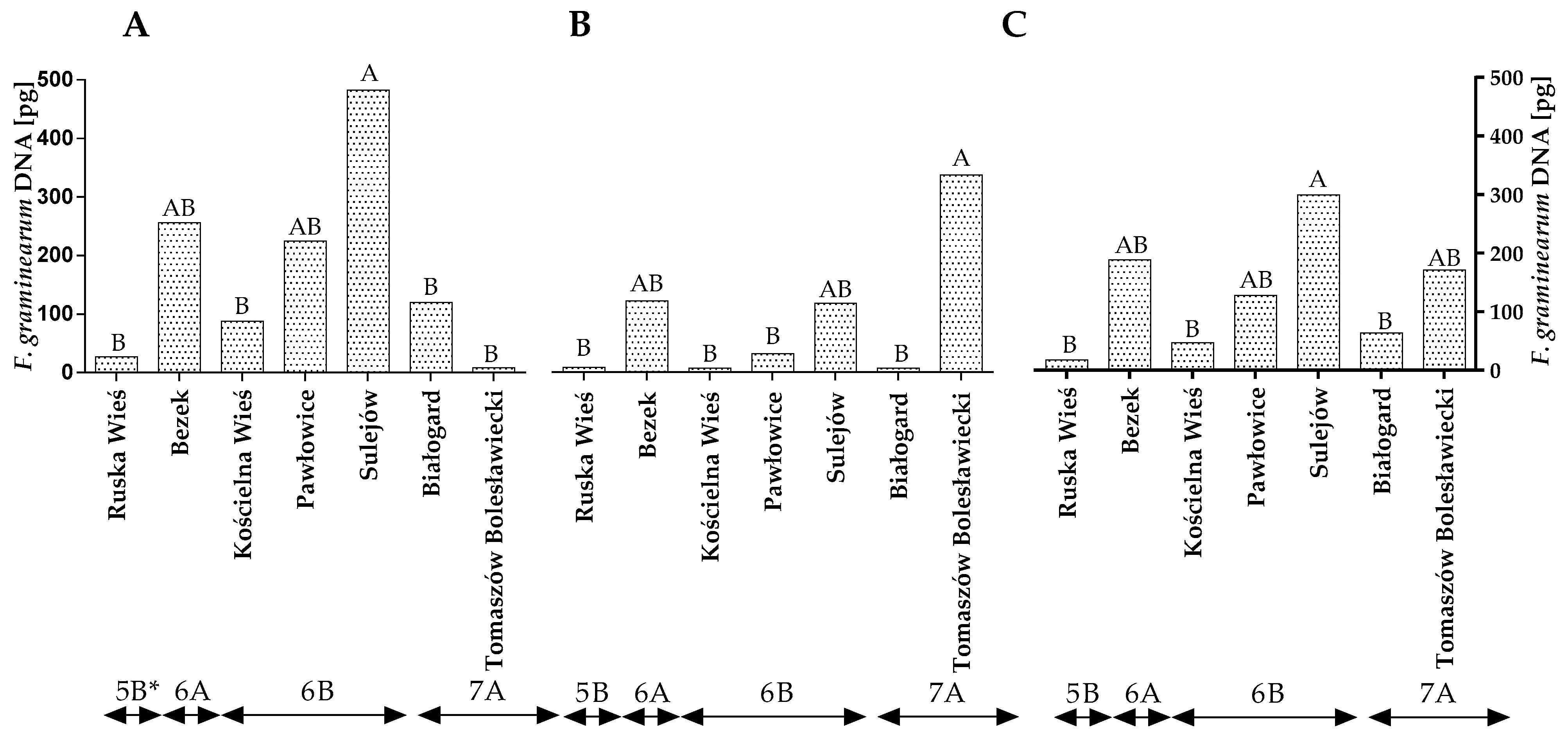
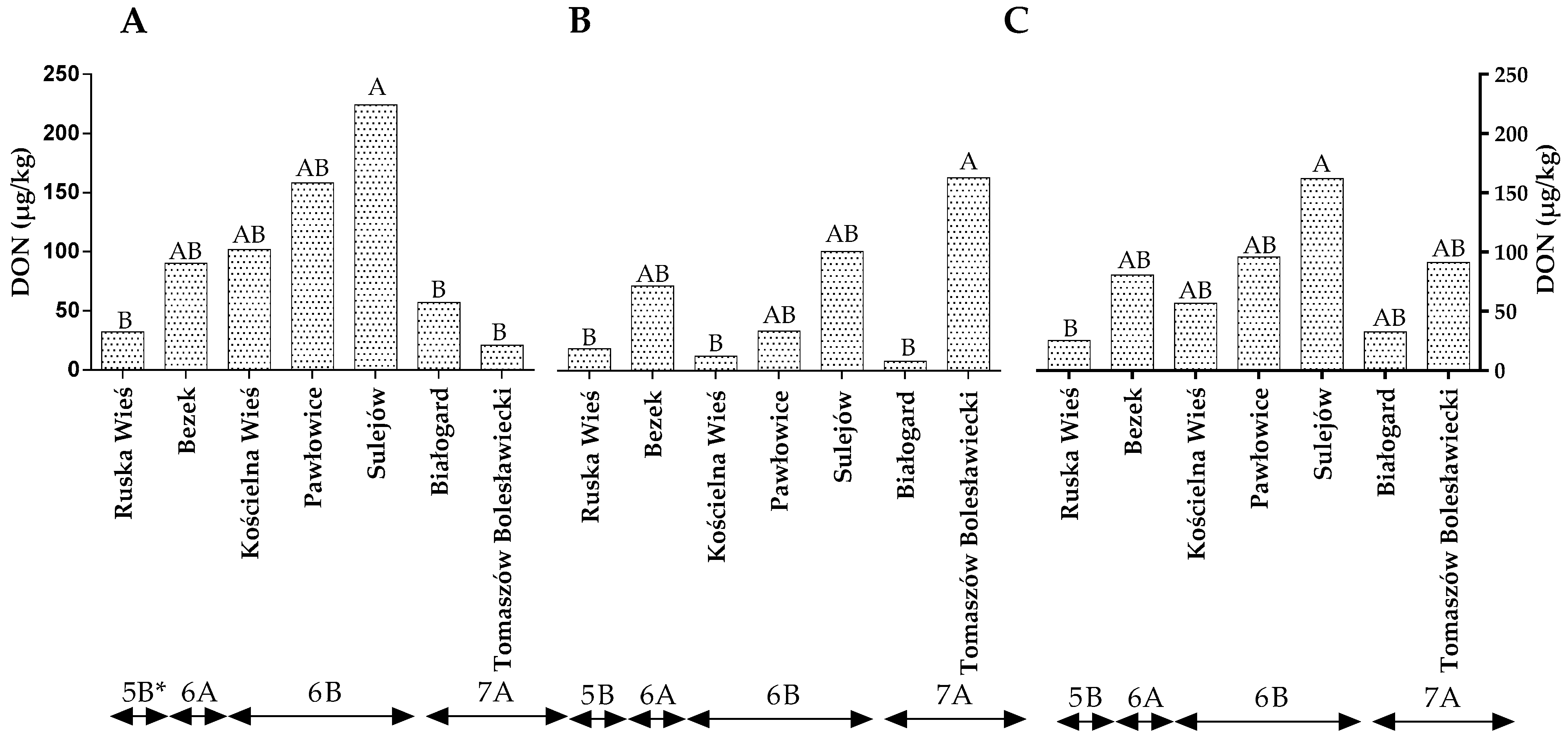
2.2. Quantification of F. culmorum and F. graminearum DNA by qPCR and Quantification of DON by HPLC
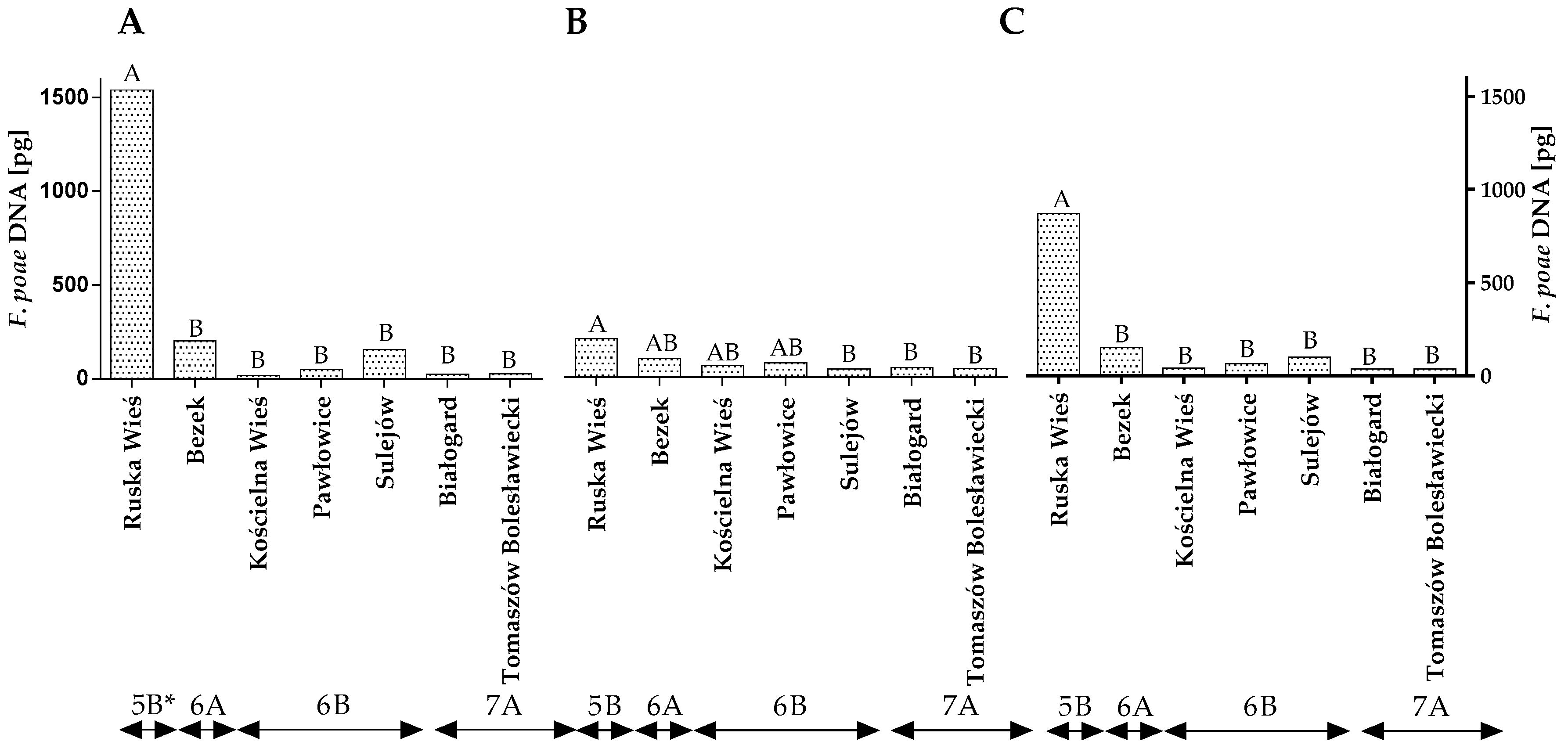
2.3. DNA Quantification by qPCR in Other Fungal Species
2.4. The Influence of Climate on the Prevalence of Fungi
2.5. Correlation between the Number of Fungal Isolates Identified by the Cultural Method and the Quantity of the DNA of Fungal Species
3. Discussion
4. Conclusions
- Fusarium avenaceum and F. graminearum were the predominant fungal species in the winter wheat grains cultivated in most of the analyzed locations in Poland.
- The prevalence of fungal pathogens was influenced by weather conditions during the growing season of winter wheat, and DON-producing species were predominant in climatic regions 6A and 6B, which also contributed to higher grain contamination with DON.
- Fusarium avenaceum, F. poae and P. verrucosum were the predominant species in the winter wheat grains grown in the coldest climatic region.
- Mathematical analyses revealed a positive correlation between the quantity of F. avenaceum DNA (pg) and temperatures (°C) in the first ten days of May, as well as humidity (mm) between 11 and 20 May.
- Spearman’s rank correlation analysis confirmed the presence of competitive interactions between enniatin-producing F. avenaceum and DON-producing F. culmorum and F. graminearum.
- A positive correlation was observed between the presence of F. culmorum and F. graminearum DNA and DON contamination of winter wheat grains.
5. Materials and Methods
5.1. Field Experiment
5.2. Isolation and Identification of Fungi by the Culture-Based Method
5.3. Extraction and Quantification of Genomic DNA from Fusarium fungi Colonizing Winter Wheat Grain
5.4. Identification of Selected Fusarium Species and P. verrucosum by qPCR
5.5. Detection and Quantification of DON
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stuper-Szablewska, K.; Perkowski, J. Contamination of wheat grain with microscopic fungi and their metabolites in Poland in 2006–2009. Ann. Agric. Environ. Med. 2014, 21, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, M.; Backhouse, D.; Ponte, E.M.D. Climate change impacts on the ecology of Fusarium graminearum species complex and susceptibility of wheat to Fusarium head blight: A review. World Mycotoxin J. 2016, 9, 685–700. [Google Scholar] [CrossRef]
- Vujanovic, V.; Mavragani, D.; Hamel, C. Fungal communities associated with durum wheat production system: A characterization by growth stage, plant organ and preceding crop. Crop Prot. 2012, 37, 26–34. [Google Scholar] [CrossRef]
- Pastuszak, J.; Szczerba, A.; Dziurka, M.; Hornyák, M.; Kope’c, P.; Szklarczyk, M.; Płazek, A. Physiological and Biochemical Response to Fusarium culmorum Infection in Three Durum Wheat Genotypes at Seedling and Full Anthesis Stage. Int. J. Mol. Sci. 2021, 22, 7433. [Google Scholar] [CrossRef]
- Obanor, F.; Chakraborty, S. Aetiology and toxigenicity of Fusarium graminearum and F. pseudograminearum causing crown rot and head blight in Australia under natural and artificial infection. Plant Pathol. 2014, 63, 1218–1229. [Google Scholar] [CrossRef]
- Leslie, J.F.; Moretti, A.; Mesterházy, Á.; Ameye, M.; Audenaert, K.; Singh, P.K.; Richard-Forget, F.; Chulze, S.N.; Del Ponte, E.M.; Chala, A.; et al. Key Global Actions for Mycotoxin Management in Wheat and Other Small Grains. Toxins 2021, 13, 725. [Google Scholar] [CrossRef]
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of wheat diseases—A field perspective. Mol. Plant Pathol. 2018, 19, 1523–1536. [Google Scholar] [CrossRef]
- van der Fels-Klerx, H.J.; de Rijk, T.C.; Booij, C.J.H.; Goedhart, P.W.; Boers, E.A.M.; Zhao, C.; Waalwijk, C.; Mol, H.G.J.; van der Lee, T.A.J. Occurrence of Fusarium Head Blight species and Fusarium mycotoxins in winter wheat in The Netherlands in 2009. Food Add. Contam. Part A 2012, 29, 1716–1726. [Google Scholar] [CrossRef]
- Somma, S.; Petruzzella, A.L.; Logrieco, A.F.; Meca, G.; Cacciola, O.S.; Moretti, A. Phylogenetic analyses of Fusarium graminearum strains from cereals in Italy, and characterization of their molecular and chemical chemotypes. Crop. Past. Sci. 2014, 65, 52–61. [Google Scholar] [CrossRef]
- Lenc, L.; Czecholiński, G.; Wyczling, D.; Turów, T.; Kaźmierczak, A. Fusarium head blight (FHB) and Fusarium spp. on grain of spring wheat cultivars grown in Poland. J. Plant Protect. Res. 2015, 55, 266–277. [Google Scholar] [CrossRef]
- Okorski, A.; Polak-Śliwińska, M.; Karpiesiuk, K.; Pszczółkowska, A.; Kozera, W. Real time PCR: A good toll to estimate mycotoxin contamination in pig diets. World Mycotoxin J. 2017, 10, 219–228. [Google Scholar] [CrossRef]
- Pszczółkowska, A.; Okorski, A.; Olszewski, J.; Jarmołkowicz, J. Fungal pathogens of the genus Fusarium in winter wheat protected with fungicides in north-eastern Poland. Acta Agrobot. 2013, 66, 95–106. [Google Scholar] [CrossRef][Green Version]
- Sadowski, C.Z.; Lenc, L.; Kuś, J. Fusarium head blight and Fusarium spp. on grain of winter wheat, a mixture of cultivars and spelt grown in organic system. J. Res. Appl. Agric. Eng. 2010, 55, 79–83. [Google Scholar]
- Buerstmayr, H.; Adam, G.; Lemmens, M. Resistance to head blight caused by Fusarium spp. in Wheat’. In Disease Resistance in Wheat; Sharma, I., Ed.; MPG Books Group: Bodmin, UK, 2012; pp. 236–276. [Google Scholar]
- Geißinger, C.; Gastl, M.; Becker, T. Enzymes from Cereal and Fusarium Metabolism Involved in the Malting Process—A Review. J. Am. Soc. Brew. Chem. 2021, 80, 1–16. [Google Scholar] [CrossRef]
- National Oceanic & Atmospheric Administration—Earth System Research Laboratory. Trends in Atmospheric Carbon Dioxide. 2020. Available online: https://www.esrl.noaa.gov/gmd/ccgg/trends (accessed on 27 January 2022).
- Palit, P.; Kudapa, H.; Zougmore, R.; Kholova, J.; Whitbread, A.; Sharma, M.; Varshney, R.K. An integrated research framework combininggenomics, systems biology, physiology, modelling and breeding forlegume improvement in response to elevated CO2 under climatechange scenario. Curr. Plant Biol. 2020, 22, 10014. [Google Scholar] [CrossRef]
- Lamichaney, A.; Maity, A. Implications of rising atmospheric carbon dioxide concentration on seed quality. Int. J. Biometeorol. 2021, 65, 805–812. [Google Scholar] [CrossRef]
- Heinze, W.; Schreiber, D. Eine neue Kartierung der Winterhärtezonen für Gehölze in Europa (Anew mapping of winter hardiness zones for woody vegetation in Europe). Mitt. Deutsch. Dendrol. Gesellschaft. 1984, 75, 11–56. [Google Scholar]
- Dawson, I.K.; Russell, J.; Powell, W.; Steffenson, B.; Thomas, W.T.B.; Waugh, R. Barley: A Translational Model for Adaptation to Climate Change. New Phytol. 2015, 206, 913–931. [Google Scholar] [CrossRef]
- Linkmeyer, A.; Hofer, K.; Rychlik, M.; Herz, M.; Hausladen, H.; Huckelhoven, R.; Hess, M. Influence of Inoculum and Climatic Factors on the Severity of Fusarium Head Blight in German Spring and Winter Barley. Food Addit. Contam. Part A Chem. Anal. Control Expo Risk Assess. 2016, 33, 489–499. [Google Scholar] [CrossRef]
- Snowarski, M. Strefy mrozoodporności—Nowe mapy. Szkółkarstwo 2012, 6, 60–67. [Google Scholar]
- Wegulo, S. Factors influencing deoxynivalenol accumulation in small grain cereals. Toxins 2012, 4, 1157–1180. [Google Scholar] [CrossRef] [PubMed]
- Tola, M.; Kebede, B.; Yildiz, F. Occurrence, importance and control of mycotoxins: A review. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar] [CrossRef]
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A Unified Effort to Fight an Enemy of Wheat and Barley: Fusarium Head Blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [PubMed]
- Moretti, A.; Pascale, M.; Logrieco, A.F. Mycotoxin risks under a climate change scenario in Europe. Tr. Food Sci. Technol. 2019, 84, 38–40. [Google Scholar] [CrossRef]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional wheat. Food Addit. Contam. 2009, 26, 496–506. [Google Scholar] [CrossRef]
- Mielniczuk, E.; Skwaryło-Bednarz, B. Fusarium Head Blight, Mycotoxins and Strategies for Their Reduction. Agronomy 2020, 10, 509. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Beyer, M.; Pasquali, M.; Jenny, E.; Musa, T.; Bucheli, T.D.; Wettstein, F.E.; Forrer, H.-R. An eight-year survey of wheat shows distinctive effects of cropping factors on different Fusarium species and associated mycotoxins. Eur. J. Agron. 2019, 105, 62–77. [Google Scholar] [CrossRef]
- Piacentini, K.C.; Rocha, L.O.; Savi, G.D.; Carnielli-Queiroz, L.; De Carvalho Fontes, L.; Correa, B. Assessment of Toxigenic Fusarium Species and Their Mycotoxins in Brewing Barley Grains. Toxins 2019, 11, 31. [Google Scholar] [CrossRef]
- Timmusk, S.; Nevo, E.; Ayele, F.; Noe, S.; Niinemets, Y. Fighting Fusarium Pathogens in the Era of Climate Change: A Conceptual Approach. Pathogens 2020, 9, 419. [Google Scholar] [CrossRef]
- Nathanail, A.; Varga, E.; Meng-Reiterer, J.; Bueschl, C.; Michlmayr, H. Metabolism of Fusarium Mycotoxins T2 toxin, HT-2, and in wheat. J. Agric. Food Chem. 2015, 63, 7862–7872. [Google Scholar] [CrossRef]
- Smith, M.C.; Madec, S.; Coton, E. Natural Co-Occurrence of Mycotoxins in Foods and Feeds and Their in vitro Combined Toxicological Effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Teller, R.S.; Schmidt, R.J.; Whitlow, L.W. Effect of physical damage to ears of corn before harvest and treatment with various additives on the concentration of mycotoxins, silage fermentation, and aerobic stability of corn silage. J. Dairy Sci. 2012, 95, 1428–1436. [Google Scholar] [CrossRef] [PubMed]
- Spanic, V.; Horvat, D.; Drezner, G.; Zdunic, Z. Changes in protein composition in the grain and malt after Fusarium infection dependently of wheat resistance. Pathogens 2019, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Beccari, G.; Arellano, C.; Covarelli, L.; Tini, F.; Sulyok, M.; Cowger, C. Effect of wheat infection timing on Fusarium head blight causal agents and secondary metabolites in grain. Int. J. Food Microbiol. 2019, 90, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Juan, C.; Ferrer, E.; Mañes, J. Fusarium species, chemotype characterisation and trichothecene contamination of durum and soft wheat in an area of central Italy. J. Sci. Food Agric. 2015, 95, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Häller, G.B.; Munich, M.; Kleijer, G.; Mascher, F. Characterisation of kernel resistance against Fusarium infection in spring wheat by baking quality and mycotoxin assessments. Eur. J. Plant Pathol. 2008, 120, 61–68. [Google Scholar]
- Marzec-Schmidt, K.; Börjesson, T.; Suproniene, S.; Jędryczka, M.; Janavičienė, S.; Góral, T.; Karlsson, I.; Kochiieru, Y.; Ochodzki, P.; Mankevičienė, A.; et al. Modelling the Effects of Weather Conditions on Cereal Grain Contamination with Deoxynivalenol in the Baltic Sea Region. Toxins 2021, 13, 737. [Google Scholar] [CrossRef]
- Bryła, M.; Ksieniewicz-Woźniak, E.; Yoshinari, T.; Waśkiewicz, A.; Szymczyk, K. Contamination of Wheat Cultivated in Various Regions of Poland during 2017 and 2018 Agricultural Seasons with Selected Trichothecenes and Their Modified Forms. Toxins 2019, 11, 88. [Google Scholar] [CrossRef]
- Klem, K.; Vanova, M.; Hajslova, J.; Lancova, K.; Sehnalova, M. A neural network model for prediction of deoxynivalenol content in wheat grain based on weather data and preceding crop. Plant Soil Env. 2007, 53, 421–429. [Google Scholar] [CrossRef]
- Bensassi, F.; Mahdi, C.; Bacha, H.; Hajlaoui, M.R. Survey of the mycobiota of freshly harvested wheat grains in the main production areas of Tunisia. Afr. J. Food Sci. 2011, 5, 292–298. [Google Scholar]
- Desjardins, A.E. Fusarium mycotoxins. In Chemistry, Genetics, and Biology; APS Press: St. Paul, MN, USA, 2006. [Google Scholar]
- González-Osnaya, L.; Farrés, A. Deoxynivalenol and zearalenone in Fusarium-contaminated wheat in Mexico City. Food Addit. Contam. Part B. 2011, 4, 71–78. [Google Scholar] [CrossRef]
- Yoshizawa, T. Thirty-five Years of Research on Deoxynivalenol, a Trichothecene Mycotoxin: With Special Reference to Its Discovery and Co-occurrence with Nivalenol in Japan. Food Saf. 2013, 1, 2013002. [Google Scholar] [CrossRef]
- Winter, M.; Koopmann, B.; Döll, K.; Karlovsky, P.; Kropf, U.; Schlüter, K.; Tiedemann, A. Mechanisms regulating grain contamination with trichothecenes trans-located from the stem base of wheat (Triticum aestivum) infected with Fusarium culmorum. Phytopathology 2013, 103, 682–689. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2012, 57, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.M.; Nicholson, P.; Thomsett, M.A.; Simpson, D.; Cooke, B.M.; Doohan, F.M.; Brennan, J.; Monaghan, S.; Moretti, A.; Mule, G.; et al. Relationship between the fungal complex causing Fusarium head blight of wheat and environmental conditions. Phytopathology 2008, 98, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Siou, D.; Gélisse, S.; Laval, V.; Suffert, F.; Lannou, C. Mutual Exclusion between Fungal Species of the Fusarium Head Blight Complex in a Wheat Spike. Appl. Environ. Microbiol. 2015, 81, 4682–4689. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Mulè, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. Eur. J. Plant Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Ederli, L.; Beccari, G.; Tini, F.; Bergamini, I.; Bellezza, I.; Romani, R.; Covarelli, L. Enniatin B and Deoxynivalenol Activity on Bread Wheat and on Fusarium Species Development. Toxins 2021, 13, 728. [Google Scholar] [CrossRef] [PubMed]
- Eranthodi, A.; Schneiderman, D.; Harris, L.J.; Witte, T.E.; Sproule, A.; Hermans, A.; Overy, D.P.; Chatterton, S.; Liu, J.; Li, T.; et al. Enniatin production influences Fusarium avenaceum virulence on potato tubers, but not on durum wheat or peas. Pathogens 2020, 9, 75. [Google Scholar] [CrossRef]
- Pollard, A.T.; Okubara, P.A. Real-time PCR quantification of Fusarium avenaceum in soil and seeds. J. Microbiol. Methods 2019, 157, 21–30. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Musa, T.; Bänziger, I.; Kägi, A.; Bucheli, T.D.; Wettstein, F.E.; Pasquali, M.; Forrer, H.R. Fusarium Mycotoxins in Swiss Wheat: A Survey of Growers’ Samples between 2007 and 2014 Shows Strong Year and Minor Geographic Effects. Toxins 2017, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Hoboken, NY, USA, 2006; pp. 81–159. [Google Scholar]
- Uhlig, S.; Jestoi, M.; Parikka, P. Fusarium avenaceum- the north European situation. Int. J. Food Microbiol. 2007, 119, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Byamukama, E.; Shaukat, A.; Kleinjan, J.; Yabwalo, D.N.; Graham, C.; Caffe-Treml, M.; Mueller, N.D.; Rickertsen, R.; Berzonsky, W.A. Winter Wheat Grain Yield Response to Fungicide Application is Influenced by Cultivar and Rainfall. Plant Pathol. J. 2019, 35, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Doohan, F.M.; Brennan, J.; Cooke, B.M. Influence of climatic factors on Fusarium species pathogenic to cereals. Eur. J. Plant Pathol. 2003, 109, 755–768. [Google Scholar] [CrossRef]
- Reis, E.M.; Boareto, C.; Durante Danelli, A.L.; Zoldan, S.M. Anthesis, the infectious process and disease progress curves for Fusarium head blight in wheat. Summa Phytopathol. 2016, 42, 134–139. [Google Scholar] [CrossRef]
- Postic, J.; Cosic, J.; Vrandecic, K.; Jurkovic, D.; Saleh, A.A.; Leslie, J.F. Diversity of Fusarium Species Isolated from Weeds and Plant Debris in Croatia. J. Phytopathol. 2011, 160, 76–81. [Google Scholar] [CrossRef]
- Shah, L.; Ali, A.; Yahya, M.; Zhu, Y.; Wang, S.; Si, H.; Ma, C. Integrated control of Fusarium head blight and deoxynivalenol mycotoxin in wheat. Plant Pathol. 2017, 67, 532–548. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Tóth, B.; Szabó-Hevér, Á.; Varga, J.; Lehoczki-Krsjak, S. Node Infection Caused by Fusarium graminearum in Wheat. Cereal Res. Commun. 2008, 36, 471–475. [Google Scholar] [CrossRef]
- Manstretta, V.; Gourdain, E.; Rossi, V. Deposition patterns of Fusarium graminearum ascospores and conidia within a wheat canopy. Europ. J. Plant Pathol. 2015, 143, 873–880. [Google Scholar] [CrossRef]
- Scala, V.; Aureli, G.; Cesarano, G.; Incerti, G.; Fanelli, C.; Scala, F.; Reverberi, M.; Bonanomi, G. Climate, Soil Management, and Cultivar Affect Fusarium Head Blight Incidence and Deoxynivalenol Accumulation in Durum Wheat of Southern Italy. Front. Microbiol. 2014, 7, 1014. [Google Scholar] [CrossRef]
- Bankina, B.; Bimšteine, G.; Neusa-Luca, I.; Roga, A.; Fridmanis, D. What influences the composition of fungi in wheat grains. Acta Agrobot. 2017, 70, 1726. [Google Scholar] [CrossRef][Green Version]
- Kulik, T.; Jestoi, M. Quantification of Fusarium poae DNA and associated mycotoxins in asymptomatically contaminated wheat. Int. J. Food Microbiol. 2009, 130, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.-M.; Monger, W.; Ritieni, A.; Nicholson, P. Effect of temperature and duration of wetness during initial infection periods on disease development, fungal biomass and mycotoxin concentrations on wheat inoculated with single, or combinations of, Fusarium species. Plant Pathol. 2007, 56, 943–956. [Google Scholar] [CrossRef]
- Wang, Y.; Hou, Y.; Mao, X.; Liu, F.; Zhou, M. Temperature-Responded Biological Fitness of Carbendazim-Resistance Fusarium graminearum Mutants Conferring the F167Y, E198K, and E198L Substitutions. Plant Dis. 2021, 105, 3522–3530. [Google Scholar] [CrossRef] [PubMed]
- Wenda-Piesik, A.; Lemańczyk, G.; Twarużek, M.; Błajet-Kosicka, A.; Kazek, M.; Grajewski, J. Fusarium head blight incidence and detection of Fusarium toxins in wheat in relation to agronomic factors. Eur. J. Plant Pathol. 2017, 149, 515–531. [Google Scholar] [CrossRef]
- Drakopoulos, D.; Kägi, A.; Gimeno, A.; Six, J.; Jenny, E.; Forrer, H.-R.; Vogelgsang, S. Prevention of Fusarium head blight infection and mycotoxins in wheat with cut-and-carry biofumigation and botanicals. Field Crops Res. 2020, 246, 107681. [Google Scholar] [CrossRef]
- Rowlandson, T.; Gleason, M.; Sentelhas, P.; Gillespie, T.; Thomas, C.; Hornbuckle, B. Reconsidering leaf wetness duration determination for plant disease management. Plant Dis. 2015, 99, 310–319. [Google Scholar] [CrossRef]
- Hofer, K.; Huckelhoven, R.; Hess, M. Analysis of Archive Samples of Spring and Winter Barley Support an Increase in Individual Fusarium Species in Bavarian Barley Grain over the Last Decades. J. Plant Dis. Prot. 2019, 126, 247–254. [Google Scholar] [CrossRef]
- Pigati, R.L.; Dernoeden, P.H.; Grybauskas, A.P.; Momen, B. Simulated rainfall and mowing impact fungicide performance when targeting dollar spot in creeping bentgrass. Plant Dis. 2010, 94, 596–603. [Google Scholar] [CrossRef]
- Bryła, M.; Ksieniewicz-Woźniak, E.; Waśkiewicz, A.; Szymczyk, K.; Jędrzejczak, R. Natural Occurrence of Nivalenol, Deoxynivalenol, and Deoxynivalenol-3-Glucoside in Polish Winter Wheat. Toxins 2018, 10, 81. [Google Scholar] [CrossRef]
- ISO-24333; Cereal and Cereal Products—Sampling. ISO: Geneva, Switzerland, 2009.
- Gerlach, W.; Nirenberg, H. The Genus Fusarium—A Pictorial Atlas; Biologische Bundesanstalt für Land- und Forstwirtschaft: Berlin, Germany, 1982. [Google Scholar]
- Watanabe, I.; Kakishima, M.; Adachi, Y.; Nakajima, H. Potential mycotoxin productivity of Alternaria alternata isolated from garden trees. Mycotoxins 2007, 57, 34–45. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Kulik, T.; Jestoi, M.; Okorski, A. Development of TaqMan assays for the quantitative detection of Fusarium avenaceum/Fusarium tricinctum and Fusarium poae esyn1 genotypes from cereal grain. FEMS Microbiol. Lett. 2011, 314, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Nicolaisen, M.; Supronienė, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Justesen, A.F. Real-time PCR for quantification of eleven individual Fusarium species in cereals. J. Microbiol. Methods 2009, 76, 234–240. [Google Scholar] [CrossRef]
- Vegi, A.; Wolf-Hall, C.E. Multiplex real-time PCR method for detection and quantification of mycotoxigenic fungi belonging to three different genera. J. Food Sci. 2013, 78, M70–M76. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Location (L) | Variety (V) | L × V |
|---|---|---|---|
| F. avenaceum/F. tricinctum DNA | ** | ns | ** |
| F. culmorum DNA | * | ns | * |
| F. graminearum DNA | ** | ns | ** |
| F. poae DNA | *** | * | *** |
| P. verrucosum DNA | *** | ns | *** |
| DON (µg/kg) | ** | ns | * |
| Month | Days | DNA Quantity | DON (µg/kg) | ||||
|---|---|---|---|---|---|---|---|
| F. avenaceum/F. tricinctum | F. culmorum | F. graminearum | F. poae | P. verrucosum | |||
| May | 1–10 | 0.54 * | −0.20 | −0.16 | −0.19 | −0.23 | −0.22 |
| 11–20 | 0.09 | 0.18 | 0.31 | −0.22 | −0.30 | 0.18 | |
| 21–31 | 0.35 | −0.20 | −0.18 | −0.20 | −0.26 | −0.18 | |
| June | 1–10 | 0.06 | 0.19 | 0.18 | −0.09 | −0.28 | 0.16 |
| 11–20 | 0.03 | 0.12 | 0.20 | −0.13 | −0.25 | 0.16 | |
| 21–30 | 0.33 | −0.15 | −0.19 | −0.23 | −0.30 | −0.14 | |
| July | 1–10 | −0.39 | 0.17 | 0.37 | 0.06 | −0.01 | 0.16 |
| 11–20 | −0.07 | 0.21 | 0.35 | −0.30 | −0.29 | 0.26 | |
| 21–31 | 0.25 | 0.09 | 0.02 | −0.30 | −0.25 | 0.09 | |
| Month | Days | DNA Quantity | DON (µg/kg) | ||||
|---|---|---|---|---|---|---|---|
| F. avenaceum/F. tricinctum | F. culmorum | F. graminearum | F. poae | P. verrucosum | |||
| May | 1–10 | −0.02 | −0.04 | 0.09 | 0.25 | 0.18 | 0.08 |
| 11–20 | 0.50 * | −0.21 | −0.23 | 0.02 | −0.10 | −0.25 | |
| 21–31 | −0.07 | −0.28 | 0.15 | −0.06 | −0.21 | 0.01 | |
| June | 1–10 | 0.09 | −0.20 | −0.13 | −0.24 | 0.02 | −0.06 |
| 11–20 | 0.06 | −0.27 | 0.17 | −0.24 | −0.24 | 0.12 | |
| 21–30 | −0.22 | 0.08 | −0.22 | 0.01 | 0.17 | −0.18 | |
| July | 1–10 | 0.24 | −0.18 | −0.22 | −0.11 | −0.12 | −0.27 |
| 11–20 | 0.12 | −0.16 | 0.06 | −0.26 | −0.15 | 0.13 | |
| 21–31 | −0.02 | −0.19 | −0.22 | 0.13 | 0.02 | −0.27 | |
| DNA Quantity | Number of Isolates | ||||
|---|---|---|---|---|---|
| F. avenaceum | F. tricinctum | F. culmorum | F. graminearum | P. verrucosum | |
| F. avenaceum/F. tricinctum | 0.49 * | 0.26 | 0.15 | −0.11 | −0.19 |
| F. culmorum | −0.32 | −0.17 | 0.27 | −0.04 | 0.11 |
| F. graminearum | −0.30 | 0.07 | 0.11 | −0.15 | −0.06 |
| F. poae | 0.00 | 0.18 | −0.25 | 0.17 | −0.22 |
| P. verrucosum | 0.01 | 0.02 | −0.06 | 0.18 | 0.03 |
| DNA Quantity | qPCR Detection (DNA) | DON (µg/kg) | |||
|---|---|---|---|---|---|
| F. culmorum | F. graminearum | F. poae | P. verrucosum | ||
| F. avenaceum/F. tricinctum | −0.49 * | −0.59 * | −0.04 | −0.03 | −0.58 * |
| F. culmorum | - | 0.32 | −0.09 | 0.13 | 0.51 * |
| F. graminearum | - | - | −0.11 | −0.09 | 0.89 ** |
| F. poae | - | - | - | 0.09 | −0.14 |
| P. verrucosum | - | - | - | - | −0.01 |
| Location | GPS | Climatic Region |
|---|---|---|
| Białogard | φ = 54°00′, λ = 16°00′, H = 24 m a.s.l. | 7A |
| Bezek | φ = 51°11′, λ = 23°15′, H = 224 m a.s.l. | 6A |
| Kościelna Wieś | φ = 51°48′, λ = 18°01′, H = 120 m a.s.l. | 6B |
| Ruska Wieś | φ = 53°53′, λ = 22°28′, H = 130 m a.s.l. | 5B |
| Sulejów | φ = 51°35′, λ = 19°86′, H = 188 m a.s.l. | 6B |
| Tomaszów Bolesławiecki | φ = 51°17′, λ = 15°41′, H = 200 m a.s.l. | 7A |
| Pawłowice | φ = 50°28′, λ = 18°29′, H = 240 m a.s.l. | 6B |
| Genotype/Gene | Primer/Probe | Sequence (5′-3′) | Regression Equation, Efficiency of qPCR (E) | References |
|---|---|---|---|---|
| F. avenaceum/F. tricinctum | Avetric f | 5′-AGCAGTCGAGTTCGTCAACAGA-3′ | y = −3.35x + 37.21 | [80] |
| Esyn1 | Avetric r | 5′-GGCYTTTCCTGCGAACTTG-3′ | E = 98.5 | |
| Avetric probe | FAM—CCGTCGAGTCCTCT—MGB | R2 = 0.99 | ||
| F. culmorum | FculC561 fwd | 5′-CACCGTCATTGGTATGTTGTCACT-3′ | y = −3.49x + 35.45 | [81] |
| EF1α | FculC614 rev | 5′-CGGGAGCGTCTGATAGTCG-3′ | E = 93.6 R2 = 0.98 | |
| F. graminearum | FgramB379 fwd | 5′-CCATTCCCTGGGCGCT-3′ | y = −3.29x + 33.32 | [81] |
| EF1α | FgramB411 rev | 5′-CCTATTGACAGGTGGTTAGTGACTGG-3′ | E = 100 R2 = 0.97 | |
| F. poae | Poae f | 5′-GCGGCCGCTTTTGTCA-3′ | y = −3.2x + 33.85 | [80] |
| Esyn1 | Poae r | 5′-GCCTTTCCAGCAAGAGATGGT-3′ | E = 99.8 | |
| Poae probe | FAM—AAAGCGGTCGAGTCTG—MGB | R2 = 0.99 | ||
| P. verrucosum | rRNA forward | 5′-TAAGGTGCCGGAATACACGCTCAT-3′ | y = −3.53x + 22.27 | [82] |
| rRNA | rRNA reverse | 5′-TAGTTCATTCGGCCCGTGAGTTGT-3′ | E = 92.7 | |
| PV rRNA-Probe | Fam-TCTAGACAGCCCGACGGTGGCCATGGAAGT-Tamra | R2 = 0.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okorski, A.; Milewska, A.; Pszczółkowska, A.; Karpiesiuk, K.; Kozera, W.; Dąbrowska, J.A.; Radwińska, J. Prevalence of Fusarium fungi and Deoxynivalenol Levels in Winter Wheat Grain in Different Climatic Regions of Poland. Toxins 2022, 14, 102. https://doi.org/10.3390/toxins14020102
Okorski A, Milewska A, Pszczółkowska A, Karpiesiuk K, Kozera W, Dąbrowska JA, Radwińska J. Prevalence of Fusarium fungi and Deoxynivalenol Levels in Winter Wheat Grain in Different Climatic Regions of Poland. Toxins. 2022; 14(2):102. https://doi.org/10.3390/toxins14020102
Chicago/Turabian StyleOkorski, Adam, Alina Milewska, Agnieszka Pszczółkowska, Krzysztof Karpiesiuk, Wojciech Kozera, Joanna Agnieszka Dąbrowska, and Justyna Radwińska. 2022. "Prevalence of Fusarium fungi and Deoxynivalenol Levels in Winter Wheat Grain in Different Climatic Regions of Poland" Toxins 14, no. 2: 102. https://doi.org/10.3390/toxins14020102
APA StyleOkorski, A., Milewska, A., Pszczółkowska, A., Karpiesiuk, K., Kozera, W., Dąbrowska, J. A., & Radwińska, J. (2022). Prevalence of Fusarium fungi and Deoxynivalenol Levels in Winter Wheat Grain in Different Climatic Regions of Poland. Toxins, 14(2), 102. https://doi.org/10.3390/toxins14020102