Molecular Diversity of Linear Peptides Revealed by Transcriptomic Analysis of the Venom Gland of the Spider Lycosa poonaensis

 , , ,
, , ,
Abstract
1. Introduction
2. Results and Discussion
2.1. Transcriptomic Analysis of the Venom Gland of L. poonaensis
2.1.1. Peptides Other Than Linear Cytolytic Peptides
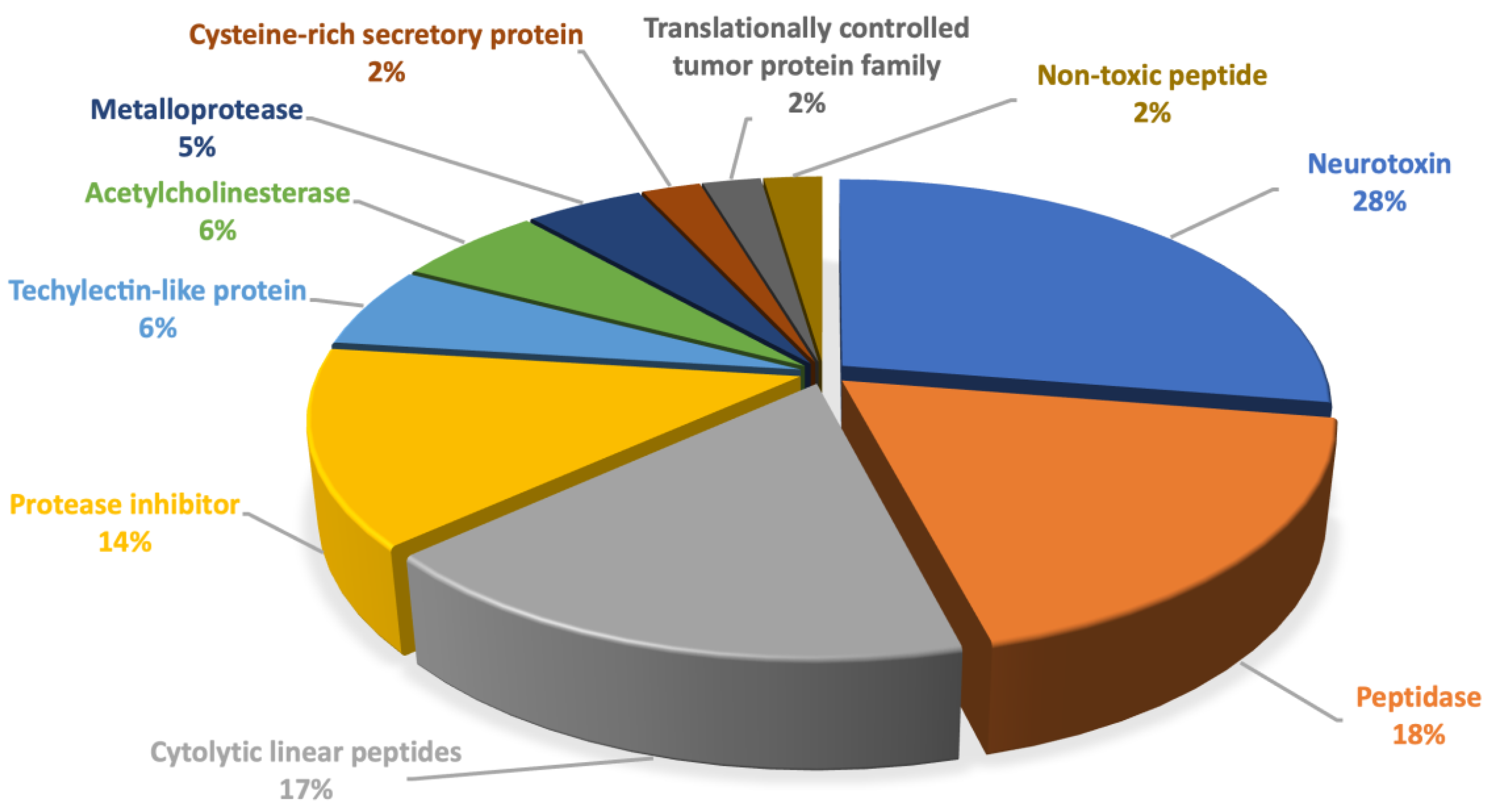
2.1.2. Enzymes and Proteins
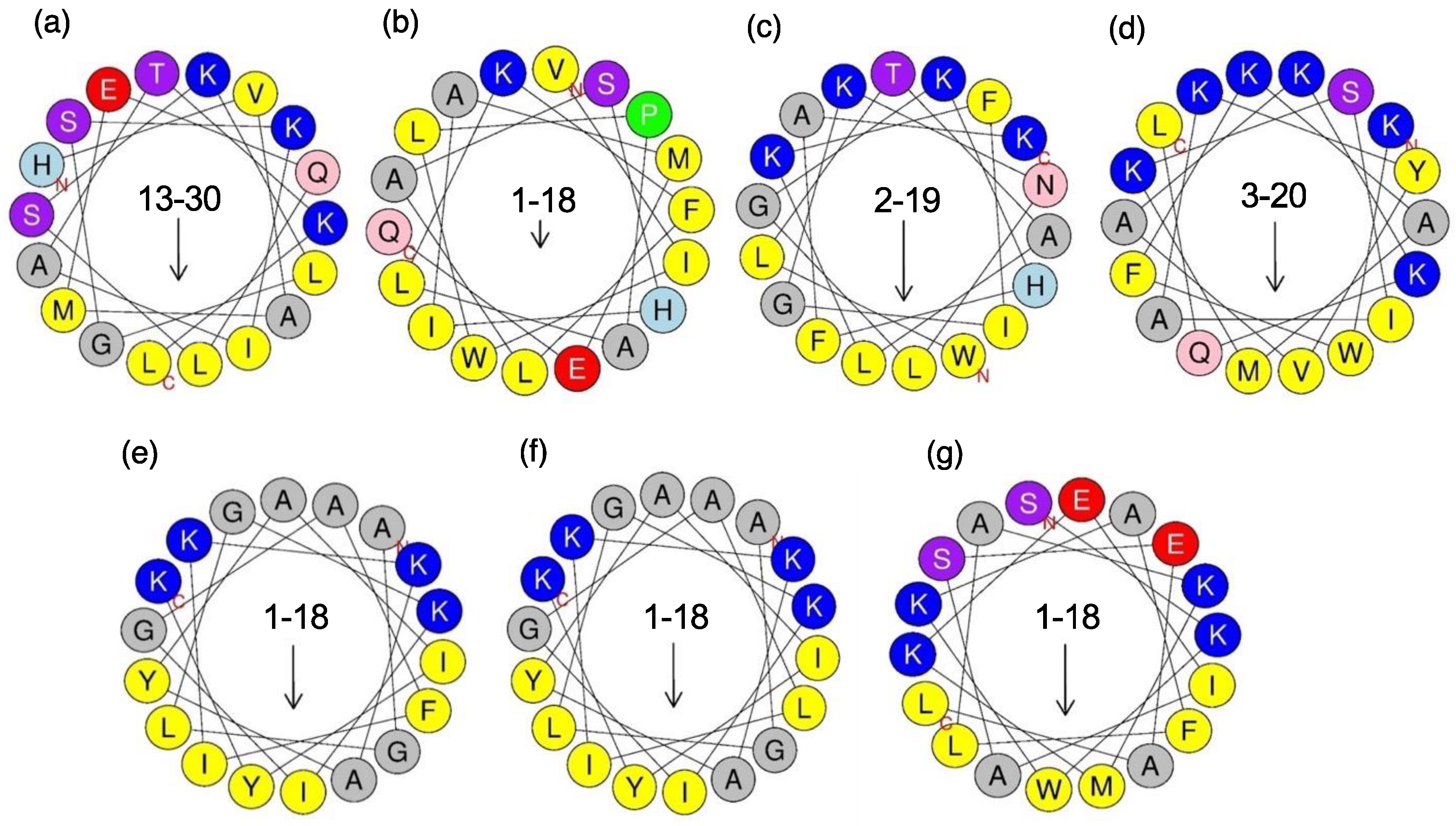
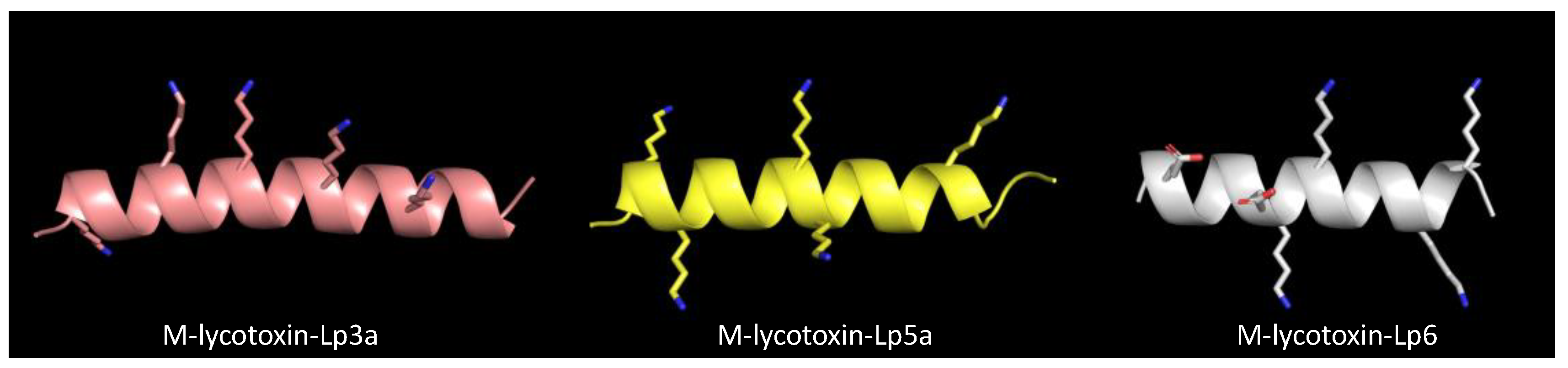
2.1.3. Cytolytic Linear Peptides
2.2. Biological Activities
3. Conclusions
4. Material and Methods
4.1. Biological Materials
4.2. RNA Extraction
4.3. Library Preparation and Sequencing
4.4. De Novo Assembly and Functional Annotations
4.5. Peptide Synthesis
4.6. Mass Spectrometric Analysis
4.7. Circular Dichroism (CD) Measurements
4.8. Antibacterial Activity
4.9. Insect Toxicity
4.10. Hemolytic Activity
4.11. 3D Structure Prediction
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Selden, P.A.; Shcherbakov, D.E.; Dunlop, J.A.; Eskov, K.Y. Arachnids from the Carboniferous of Russia and Ukraine, and the Permian of Kazakhstan. Palaeontol. Z. 2014, 88, 297–307. [Google Scholar] [CrossRef]
- Dimitrov, D.; Hormiga, G. Spider diversification through space and time. Annu. Rev. Entomol. 2021, 66, 225–241. [Google Scholar] [CrossRef]
- Langenegger, N.; Nentwig, W.; Kuhn-Nentwig, L. Spider venom: Components, modes of action, and novel strategies in transcriptomic and proteomic analyses. Toxins 2019, 11, 611. [Google Scholar] [CrossRef] [PubMed]
- Saez, N.J.; Herzig, V. Versatile spider venom peptides and their medical and agricultural applications. Toxicon 2019, 158, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Windley, M.J.; Herzig, V.; Dziemborowicz, S.A.; Hardy, M.C.; King, G.F.; Nicholson, G.M. Spider-venom peptides as bioinsecticides. Toxins 2012, 4, 191–227. [Google Scholar] [CrossRef] [PubMed]
- Gremski, L.H.; Trevisan-Silva, D.; Ferrer, V.P.; Matsubara, F.H.; Meissner, G.O.; Wille, A.C.M.; Vuitika, L.; Dias-Lopes, C.; Ullah, A.; de Moraes, F.R.; et al. Recent advances in the understanding of brown spider venoms: From the biology of spiders to the molecular mechanisms of toxins. Toxicon 2014, 83, 91–120. [Google Scholar] [CrossRef]
- Kuhn-Nentwig, L.; Stocklin, R.; Nentwig, W. Venom composition and strategies in spiders: Is everything possible? Adv. Insect Physiol. 2011, 40, 1–86. [Google Scholar] [CrossRef]
- Kuhn-Nentwig, L.; Lischer, H.E.L.; Pekar, S.; Langenegger, N.; Albo, M.J.; Isaia, M.; Nentwig, W. Linear peptides—A combinatorial innovation in the venom of some modern spiders. Front. Mol. Biosci. 2021, 8, 705141. [Google Scholar] [CrossRef]
- Gao, B.; Tian, C.H.; Zhu, S.Y. Inducible antibacterial response of scorpion venom gland. Peptides 2007, 28, 2299–2305. [Google Scholar] [CrossRef]
- Wang, X.; Wang, G. Insights into antimicrobial peptides from spiders and scorpions. Protein Peptide Lett. 2016, 23, 707–721. [Google Scholar] [CrossRef]
- Avci, F.G.; Akbulut, B.S.; Ozkirimli, E. Membrane active peptides and their biophysical characterization. Biomolecules 2018, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Wullschleger, B.; Nentwig, W.; Kuhn-Nentwig, L. Spider venom: Enhancement of venom efficacy mediated by different synergistic strategies in Cupiennius salei. J. Exp. Biol. 2005, 208, 2115–2121. [Google Scholar] [CrossRef] [PubMed]
- Megaly, A.M.A.; Yoshimoto, Y.; Tsunoda, Y.; Miyashita, M.; Abdel-Wahab, M.; Nakagawa, Y.; Miyagawa, H. Characterization of 2 linear peptides without disulfide bridges from the venom of the spider Lycosa poonaensis (Lycosidae). Biosci. Biotechnol. Biochem. 2021, 85, 1348–1356. [Google Scholar] [CrossRef]
- Pineda, S.S.; Chin, Y.K.Y.; Undheim, E.A.B.; Senff, S.; Mobli, M.; Dauly, C.; Escoubas, P.; Nicholson, G.M.; Kaas, Q.; Guo, S.D.; et al. Structural venomics reveals evolution of a complex venom by duplication and diversification of an ancient peptide-encoding gene. Proc. Natl. Acad. Sci. USA 2020, 117, 11399–11408. [Google Scholar] [CrossRef] [PubMed]
- King, G.F. Tying pest insects in knots: The deployment of spider-venom-derived knottins as bioinsecticides. Pest Manag. Sci. 2019, 75, 2437–2445. [Google Scholar] [CrossRef]
- Cassola, A.C.; Jaffe, H.; Fales, H.M.; Afeche, S.C.; Magnoli, F.; Cipolla-Neto, J. omega-Phonetoxin-IIA: A calcium channel blocker from the spider Phoneutria nigriventer. Pflug. Arch. Eur. J. Phys. 1998, 436, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Kabanova, N.V.; Vassilevski, A.A.; Rogachevskaja, O.A.; Bystrova, M.F.; Korolkova, Y.V.; Pluzhnikov, K.A.; Romanov, R.A.; Grishin, E.V.; Kolesnikov, S.S. Modulation of P2X3 receptors by spider toxins. BBA-Biomembranes 2012, 1818, 2868–2875. [Google Scholar] [CrossRef]
- Grishin, E.V.; Savchenko, G.A.; Vassilevski, A.A.; Korolkova, Y.V.; Boychuk, Y.A.; Viatchenko-Karpinski, V.Y.; Nadezhdin, K.D.; Arseniev, A.S.; Pluzhnikov, K.A.; Kulyk, V.B.; et al. Novel peptide from spider venom inhibits P2X3 receptors and inflammatory pain. Ann. Neurol. 2010, 67, 680–683. [Google Scholar] [CrossRef]
- Yuan, C.H.; He, Q.Y.; Peng, K.; Diao, J.B.; Jiang, L.P.; Tang, X.; Liang, S.P. Discovery of a distinct superfamily of Kunitz-type toxin (KTT) from tarantulas. PLoS ONE 2008, 3, e3414. [Google Scholar] [CrossRef]
- Wan, H.; Lee, K.S.; Kim, B.Y.; Zou, F.M.; Yoon, H.J.; Je, Y.H.; Li, J.H.; Jin, B.R. A spider-derived Kunitz-type serine protease inhibitor that acts as a plasmin inhibitor and an elastase inhibitor. PLoS ONE 2013, 8, e53343. [Google Scholar] [CrossRef]
- Undheim, E.A.B.; Sunagar, K.; Herzig, V.; Kely, L.; Low, D.H.W.; Jackson, T.N.W.; Jones, A.; Kurniawan, N.; King, G.F.; Ali, S.A.; et al. A proteomics and transcriptomics investigation of the venom from the Barychelid spider Trittame loki ( Brush- Foot Trapdoor). Toxins 2013, 5, 2488–2503. [Google Scholar] [CrossRef]
- Sade, Y.B.; Boia-Ferreira, M.; Gremski, L.H.; da Silveira, R.B.; Gremski, W.; Senff-Ribeiro, A.; Chaim, O.M.; Veiga, S.S. Molecular cloning, heterologous expression and functional characterization of a novel translationally-controlled tumor protein (TCTP) family member from Loxosceles intermedia (brown spider) venom. Int. J. Biochem. Cell Biol. 2012, 44, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Gremski, L.H.; Matsubara, F.H.; da Justa, H.C.; Schemczssen-Graeff, Z.; Baldissera, A.B.; Schluga, P.H.D.; Leite, I.D.; Boia-Ferreira, M.; Wille, A.C.M.; Senff-Ribeiro, A.; et al. Brown spider venom toxins: What are the functions of astacins, serine proteases, hyaluronidases, allergens, TCTP, serpins and knottins? J. Venom. Anim. Toxins 2021, 27, e20201088. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, R.B.; Wille, A.C.M.; Chaim, O.M.; Appel, M.H.; Silva, D.T.; Franco, C.R.C.; Toma, L.; Mangili, O.C.; Gremski, W.; Dietrich, C.P.; et al. Identification, cloning, expression and functional characterization of an astacin-like metalloprotease toxin from Loxosceles intermedia (brown spider) venom. Biochem. J. 2007, 406, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Langenegger, N.; Koua, D.; Schurch, S.; Heller, M.; Nentwig, W.; Kuhn-Nentwig, L. Identification of a precursor processing protease from the spider Cupiennius salei essential for venom neurotoxin maturation. J. Biol. Chem. 2018, 293, 2079–2090. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Nentwig, L.; Langenegger, N.; Heller, M.; Koua, D.; Nentwig, W. The dual prey-inactivation strategy of spiders-in-depth venomic analysis of Cupiennius salei. Toxins 2019, 11, 167. [Google Scholar] [CrossRef]
- Tadokoro, T.; Modahl, C.M.; Maenaka, K.; Aoki-Shioi, N. Cysteine-rich secretory proteins (CRISPs) from venomous snakes: An overview of the functional diversity in a large and underappreciated superfamily. Toxins 2020, 12, 175. [Google Scholar] [CrossRef]
- Santos, D.M.; Verly, R.M.; Pilo-Veloso, D.; de Maria, M.; de Carvalho, M.A.R.; Cisalpino, P.S.; Soares, B.M.; Diniz, C.G.; Farias, L.M.; Moreira, D.F.F.; et al. LyeTx I, a potent antimicrobial peptide from the venom of the spider Lycosa erythrognatha. Amino Acids 2010, 39, 135–144. [Google Scholar] [CrossRef]
- Adao, R.; Seixas, R.; Gomes, P.; Pessoa, J.C.; Bastos, M. Membrane structure and interactions of a short Lycotoxin I anaiogue. J. Pept. Sci. 2008, 14, 528–534. [Google Scholar] [CrossRef]
- Yans, L.Z.; Adams, M.E. Lycotoxins, antimicrobial peptides from venom of the wolf spider Lycosa carolinensis. J. Biol. Chem. 1998, 273, 2059–2066. [Google Scholar] [CrossRef]
- Kozlov, S.A.; Vassilevski, A.A.; Feofanov, A.V.; Surovoy, A.Y.; Karpunin, D.V.; Grishin, E.V. Latarcins, antimicrobial and cytolytic peptides from the venom of the spider Lachesana tarabaevi (Zodariidae) that exemplify biomolecular diversity. J. Biol. Chem. 2006, 281, 20983–20992. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, S.A.; Grishin, E.V. The universal algorithm of maturation for secretory and excretory protein precursors. Toxicon 2007, 49, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Nentwig, L. Complex precursor structures of cytolytic cupiennins identified in spider venom gland transcriptomes. Sci. Rep. 2021, 11, 4009. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, M.; Kitanaka, A.; Yakio, M.; Yamazaki, Y.; Nakagawa, Y.; Miyagawa, H. Complete de novo sequencing of antimicrobial peptides in the venom of the scorpion Isometrus maculatus. Toxicon 2017, 139, 1–12. [Google Scholar] [CrossRef] [PubMed]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef]
- Jiang, Z.; Hodges, R.S. Effects of net charge and the number of positively charged residues on the biological activity of amphipathic-helical cationic antimicrobial peptides. Biopolymers 2007, 88, 615. [Google Scholar]
- Lyu, Y.F.; Yang, Y.; Lyu, X.T.; Dong, N.; Shan, A.S. Antimicrobial activity, improved cell selectivity and mode of action of short PMAP-36-derived peptides against bacteria and Candida. Sci. Rep. 2016, 6, 27258. [Google Scholar] [CrossRef]
- Barreto-Santamaria, A.; Patarroyo, M.E.; Curtidor, H. Designing and optimizing new antimicrobial peptides: All targets are not the same. Crit. Rev. Clin. Lab. Sci. 2019, 56, 351–373. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Pathak, N.; Salasauvert, R.; Ruche, G.; Janna, M.H.; Mccarthy, D.; Harrison, R.G. Comparison of the effects of hydrophobicity, amphiphilicity, and alpha-helicity on the activities of antimicrobial peptides. Proteins 1995, 22, 182–186. [Google Scholar] [CrossRef]
- Rodriguez, A.; Villegas, E.; Satake, H.; Possani, L.D.; Corzo, G. Amino acid substitutions in an alpha-helical antimicrobial arachnid peptide affect its chemical properties and biological activity towards pathogenic bacteria but improves its therapeutic index. Amino Acids 2011, 40, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Nentwig, L. Antimicrobial and cytolytic peptides of venomous arthropods. Cell. Mol. Life Sci. 2003, 60, 2651–2668. [Google Scholar] [CrossRef]
- Dathe, M.; Wieprecht, T.; Nikolenko, H.; Handel, L.; Maloy, W.L.; MacDonald, D.L.; Beyermann, M.; Bienert, M. Hydrophobicity, hydrophobic moment and angle subtended by charged residues modulate antibacterial and haemolytic activity of amphipathic helical peptides. FEBS Lett. 1997, 403, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, L.; Wallace, B.A. Protein secondary structure analyses from circular dichroism spectroscopy: Methods and reference databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef]
- Mirdita, M.; Schutze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
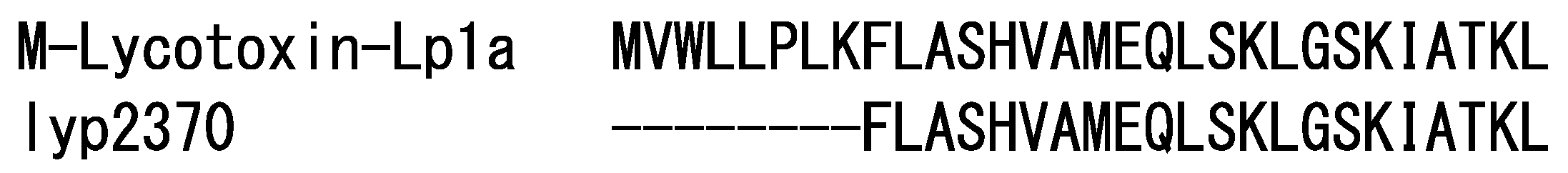{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Precursor Type | Contig | Structure of Precursor |
|---|---|---|
| Simple | m.2649 | <EEARIWLTALKFIGKNLGKHFAKQQLSKLGRSED> |
| m.2799 | <EEARIWLTALKFLGKNLGKHFAKQQLAKLGRSED> | |
| Binary | m.2698 | <EEARIWLTALKFIGKNLGKHFAKQQLSKLGRSEDMTVNDDEEARIWLTALKFIGKNLGKHFAKQQLSKLGRSED> |
| m.2695 | <EEARIWLTALKFIGKNLGKHFAKQQLSKLGRSEDMTVNDDEEARIWLTALKFLGKNLGLKFIGKNLGKHFAKQQLSKLGRSED> | |
| m.2781 | <EEARIWLTALKFLGKNLGKHFAKQQLAKLGRSEDISENLSADDDEEARIWLTALKFIGKNLLVNSIKIFR* | |
| m.2690 | <EEARIWLTALKFIGKNLGKHFAKQQLSKLGRSEDMTVNDDEEARIWLTALKFLGKNLGLKFIGKNLGKH* | |
| Complex | m.2667 | <EEARGRLQAFLAKMKEIAAQTLGREENLFANEEERVIWLPALKFLASHIAMEQLSKLGRNEQTPEEARIWLTALKFIGKNLGKHFAKQQLSKLGRNEQTPEEARIWLTALKQLSKLGRT* |
| m.2650 | <EEARGRLQAFLAKMKEIAAQTLGREENLFANEEERVIWLPALKFLASHIAMEQLSKLGRNEQTPEEARIWLTALNKFIGKNLGKHFAKQQLSKLGRNEQTPEEARIWLTALKQLSKLGRT* | |
| m.2660 | <EEARGRLQAFLAKMKEIAAQTLGREENLFANEEERVIWLPALKFLASHIAMEQLSKLGRNEQTPEEARIWLTALNKFLG* | |
| m.2706 | <EEARGRLQAFLAKMKEIAAQTLGREENLFANEEERVIWLPALKFLASHIAMEQLSKLGRNEQTPEEARIWLTALNNASEALGREEI> | |
| m.2678 | <EEARSKWKAFLAKMKEIASEALGREEGVSANEDEEARGRLQAFLAKMKEIAAQTLGREDSLSANEDEEARGRLQAFLAKMKEIAAQTLGREESLSENENEEARGRLQAFLAKMKEIAAQTLGREESVSANEDEEARSKWKAFLAKMKEIASEALGREEN> | |
| m.2715 | <EEARIWLTALKFIGKNLARIWLTALKFLGKNLGLKFIGKNLGKHFAKQQLSKLGRSEEARIWLTALKFIGKNLGKHLAKQQLSKLGRNEETPGSFSADDDELERAGLGKIGAFIKKAYAIYKAKAAGRNEQTPATVSANDDEEARKIKWFKAMKSIAKYVAKKQLKKHLGGEN* | |
| m.2734 | <EEARIWLTALKFIGKNLGKHFAKQQLSKLGRSEDMTVNDDEEARIWLTALKFIGKNLGKHFAKQQLSKLGRSEDMSANDDEEARIWLTALKFIGKNLGKHLAKQQLSKLGRNEETPGSFSADDDELERAGLGKIGAFIKKAYAIYKAKAAGRNEQTPATVSANDDEEARKIKWFKAMKSIAKYVAKKQLKKHLGGEN* | |
| m.2795 | <EEARIWLTALKFIGKNLGKHFAKQQLSKLGRSEDMTVNDDEEARIWLTALKFLGKNLGLKFIGKNLGKHFAKQQLSKLGRSEDMSANDDEEARIWLTALKFIGKNLGKHLAKQQLSKLGRNEETPGSFSADDDELERAGLGKIGAFIKKAYAIYKAKAAGRNEQTPATVSANDDEEARKIKWFKAMKSIAKYVAKKQLKKHLGGEN* | |
| m.2773 | <EEARGRLQAFLAKMKEIAAQTLGREESVSANEDEEARSKWKAFLAKMKEIASEALGREENLSANEDEEERMVWLLPLKFLASHVAMEQLSKLGSKIATKLGRNEQIPVISANEDEEERMVWLLPLKFLASHVAMEQLSKLGSKIATKLGRNEETPVSFFADDDGEERAGLGKIGALIKKAYAIYKAKAAGRNEQTPATVSANDDEEARKIKWFKAMKSIAKYVAKKQLKKHLGGEN* |
 Mature region;
Mature region;  PQM;
PQM;  iPQM;
iPQM;  Spacer or linker;
Spacer or linker;  C-terminal amidation; < Signal peptide and propeptide; > Truncated C-terminal sequence; * End of the sequence.
C-terminal amidation; < Signal peptide and propeptide; > Truncated C-terminal sequence; * End of the sequence.| Name | Sequence 1 | Detection in the Venom 2 |
|---|---|---|
| M-lycotoxin-Lp1a | MVWLLPLKFLASHVAMEQLSKLGSKIATKL-NH2 | F |
| M-lcotoxin-Lp1b | VIWLPALKFLASHIAMEQLSKL-NH2 | S |
| M-lcotoxin-Lp2 3 | GRLQAFLAKMKEIAAQTL-NH2 | F |
| M-lcotoxin-Lp3a | IWLTALKFIGKNLGKHFAKQQLSKL-NH2 | F |
| M-lcotoxin-Lp3b | IWLTALKFLGKNLGKHFAKQQLAKL-NH2 | n.d. |
| M-lcotoxin-Lp3c | IWLTALKFIGKNLGKHLAKQQLSKL-NH2 | S |
| M-lcotoxin-Lp3d | IWLTALNKFIGKNLGKHFAKQQLSKL-NH2 | S |
| M-lcotoxin-Lp3e | IWLTALKFLGKNLGLKFIGKNLGKHFAKQQLSKL-NH2 | n.d. |
| M-lcotoxin-Lp3f | IWLTALKFLGKNLGLKFIGKNLGKH | n.d. |
| M-lcotoxin-Lp3g | IWLTALKFIGKNLLVNSIKIFR | n.d. |
| M-lcotoxin-Lp3h | IWLTALKQLSKLGRT | n.d. |
| M-lcotoxin-Lp3i | IWLTALNNASEAL-NH2 | n.d. |
| M-lcotoxin-Lp3j | IWLTALNKFL-NH2 | n.d. |
| M-lcotoxin-Lp4 | KIKWFKAMKSIAKVAKKQLKKHLGGEN | F |
| M-lcotoxin-Lp5a | AGLGKIGAFIKKAAIKAKAA-NH2 | F |
| M-lcotoxin-Lp5b | AGLGKIGALIKKAAIKAKAA-NH2 | F |
| M-lcotoxin-Lp6 | SKWKAFLAKMKEIASEAL-NH2 | F |
| Name | Number of Residues | Molecular Mass | Net Charge | Hydrophobic Moment 1 | Hydrophobicity 1 |
|---|---|---|---|---|---|
| M-lycotoxin-Lp1a | 30 | 3350.9 | +4 | 0.416 | 0.358 |
| M-lycotoxin-Lp1b | 22 | 2506.4 | +2 | 0.176 | 0.838 |
| M-lycotoxin-Lp3a | 25 | 2880.7 | +6 | 0.538 | 0.510 |
| M-lycotoxin-Lp4 | 28 | 3271.9 | +8 | 0.455 | 0.316 |
| M-lycotoxin-Lp5a | 22 | 2251.3 | +6 | 0.427 | 0.449 |
| M-lycotoxin-Lp5b | 22 | 2217.3 | +6 | 0.425 | 0.444 |
| M-lycotoxin-Lp6 | 18 | 2049.1 | +3 | 0.482 | 0.355 |
| Peptides | Solvents 1 | Percentage of Secondary Structure 2 | |||
|---|---|---|---|---|---|
| α-Helix | β-Sheet | Turns | Unordered | ||
| M-lycotoxin-Lp1a | I | 15 | 26 | 25 | 35 |
| II | 61 | 10 | 11 | 19 | |
| M-lycotoxin-Lp1b | I | 10 | 31 | 26 | 33 |
| II | 57 | 9 | 12 | 22 | |
| M-lycotoxin-Lp3a | I | 11 | 29 | 26 | 34 |
| II | 59 | 16 | 8 | 16 | |
| M-lycotoxin-Lp4 | I | 8 | 33 | 26 | 33 |
| II | 51 | 11 | 13 | 23 | |
| M-lycotoxin-Lp5a | I | 15 | 23 | 28 | 33 |
| II | 55 | 17 | 10 | 18 | |
| M-lycotoxin-Lp5b | I | 7 | 33 | 25 | 34 |
| II | 51 | 14 | 11 | 23 | |
| M-lycotoxin-Lp6 | I | 20 | 22 | 25 | 33 |
| II | 55 | 14 | 8 | 22 | |
| Peptide | Antibacterial Activity MIC (µM) | Insect Toxicity 1 PD50 (µg/g Weight) | Hemolysis EC50 (µM) | ||
|---|---|---|---|---|---|
| E. coli | S. aureus | B. subtilis | |||
| M-lycotoxin-Lp1a | 6.2–12.5 | 12.5–25 | 12.5–25 | 35 (5.92–20.4) | >100 |
| M-lycotoxin-Lp1a(9-30) 2 | >100 | >100 | 100–200 | >1000 | >100 |
| M-lycotoxin-Lp1b | 50–100 | 3.1–6.2 | 6.2–12.5 | 31 (13.7–68.2) | >100 |
| M-lycotoxin-Lp3a | 0.8–1.6 | 0.4–0.8 | 0.8–1.6 | 70 (46.4–149) | >100 |
| M-lycotoxin-Lp4 | 0.4–0.8 | 1.5–3.1 | 3.1–6.2 | 10 (5.8–17.1) | >100 |
| M-lycotoxin-Lp5a | 1.25–2.5 | 50–100 | 1.25–2.5 | 50 (18.3–138) | >100 |
| M-lycotoxin-Lp5b | 1.25–2.5 | 50–100 | 1.25–2.5 | 44 (12.2–160) | >100 |
| M-lycotoxin-Lp6 | 5–12.5 | >100 | 5–12.5 | 94 (53.4–168) | >100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Megaly, A.M.A.; Miyashita, M.; Abdel-Wahab, M.; Nakagawa, Y.; Miyagawa, H. Molecular Diversity of Linear Peptides Revealed by Transcriptomic Analysis of the Venom Gland of the Spider Lycosa poonaensis. Toxins 2022, 14, 854. https://doi.org/10.3390/toxins14120854
Megaly AMA, Miyashita M, Abdel-Wahab M, Nakagawa Y, Miyagawa H. Molecular Diversity of Linear Peptides Revealed by Transcriptomic Analysis of the Venom Gland of the Spider Lycosa poonaensis. Toxins. 2022; 14(12):854. https://doi.org/10.3390/toxins14120854
Chicago/Turabian StyleMegaly, Alhussin Mohamed Abdelhakeem, Masahiro Miyashita, Mohammed Abdel-Wahab, Yoshiaki Nakagawa, and Hisashi Miyagawa. 2022. "Molecular Diversity of Linear Peptides Revealed by Transcriptomic Analysis of the Venom Gland of the Spider Lycosa poonaensis" Toxins 14, no. 12: 854. https://doi.org/10.3390/toxins14120854
APA StyleMegaly, A. M. A., Miyashita, M., Abdel-Wahab, M., Nakagawa, Y., & Miyagawa, H. (2022). Molecular Diversity of Linear Peptides Revealed by Transcriptomic Analysis of the Venom Gland of the Spider Lycosa poonaensis. Toxins, 14(12), 854. https://doi.org/10.3390/toxins14120854