Glutamine Synthetase Contributes to the Regulation of Growth, Conidiation, Sclerotia Development, and Resistance to Oxidative Stress in the Fungus Aspergillus flavus

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
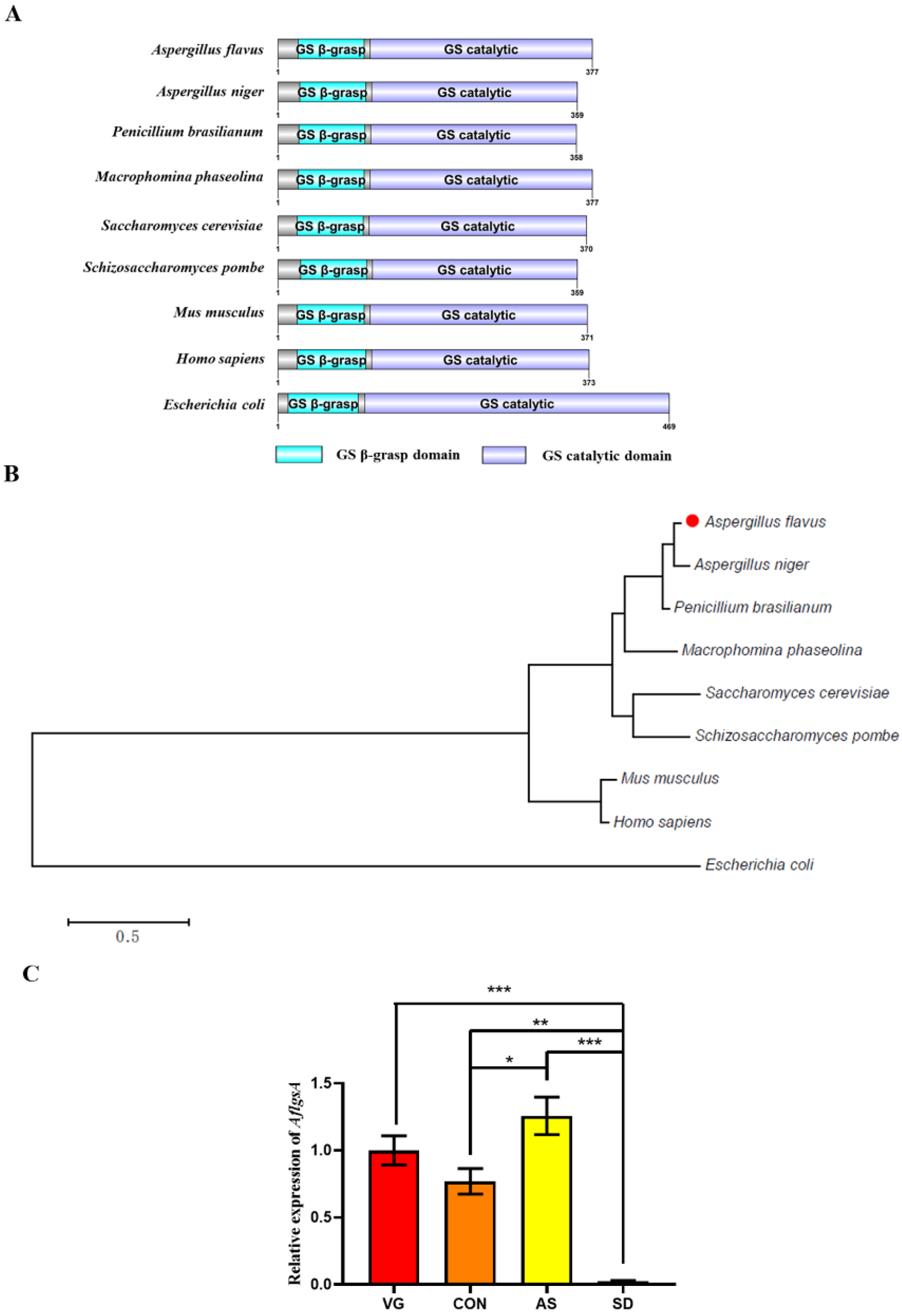
2.1. Identification of AflGsA in A. flavus
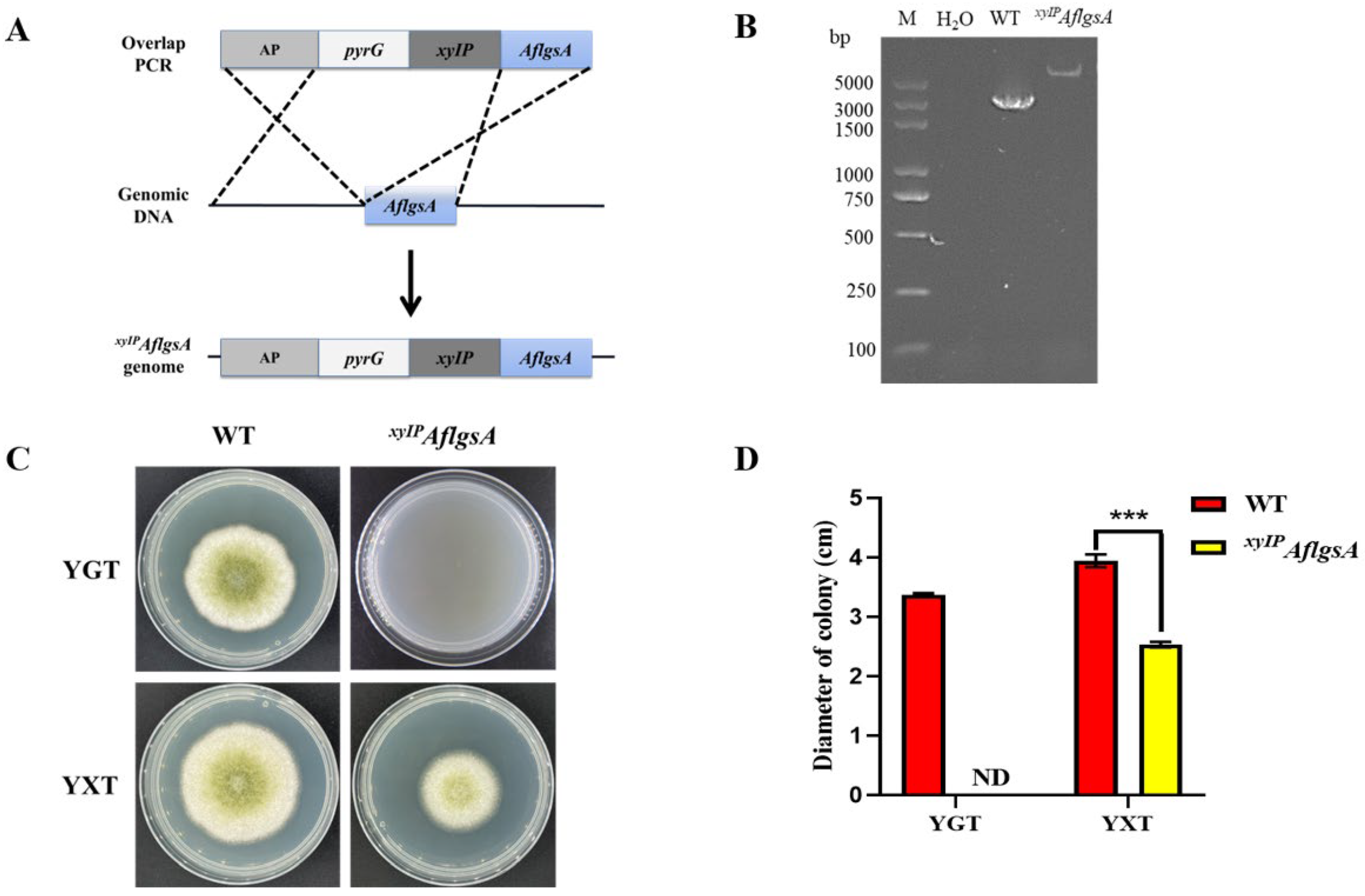
2.2. AflgsA Is an Essential Gene for the Growth of A. flavus
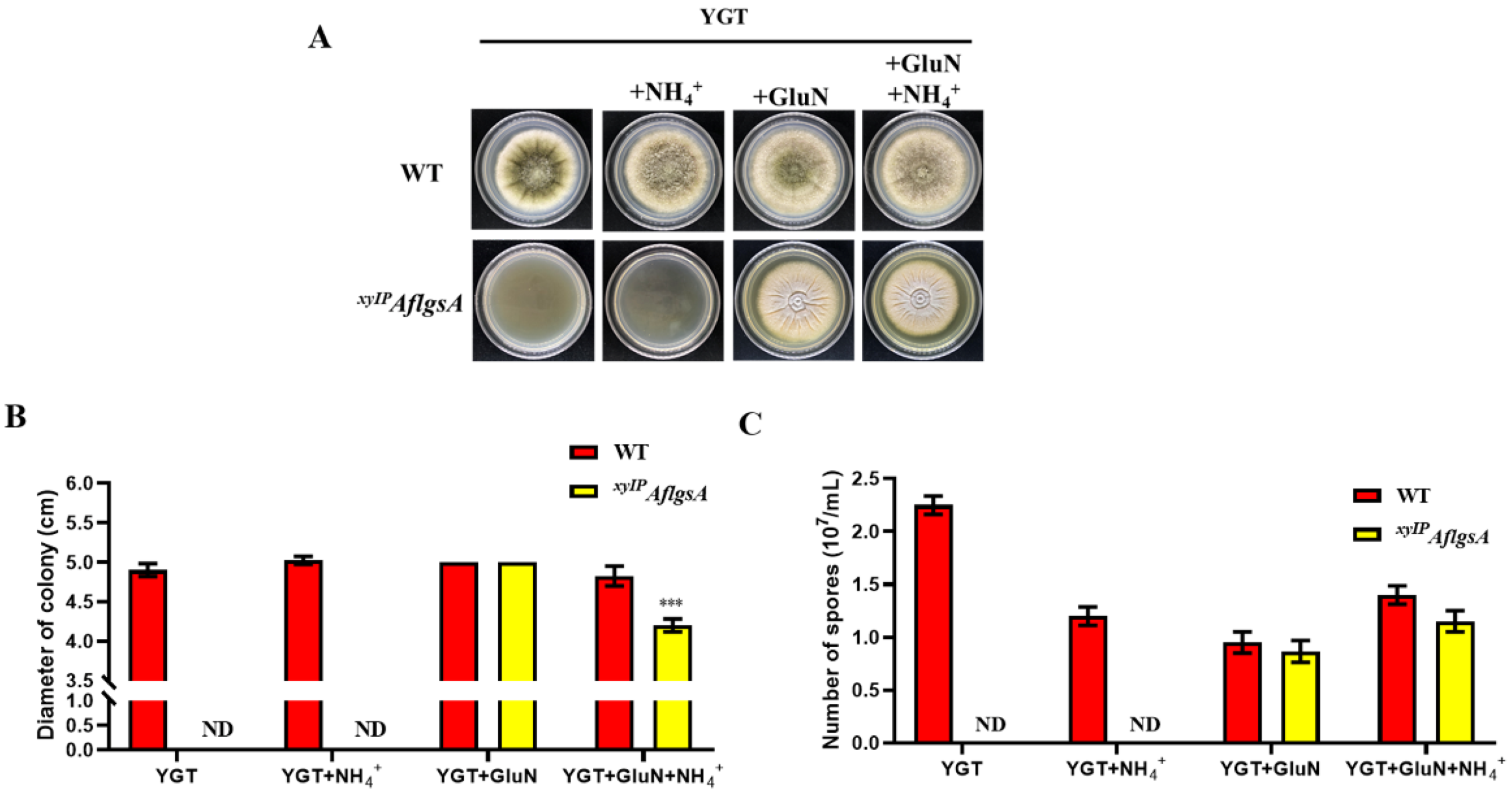
2.3. Glutamine Restores the Growth Defect of the xyIPAflgsA Strain
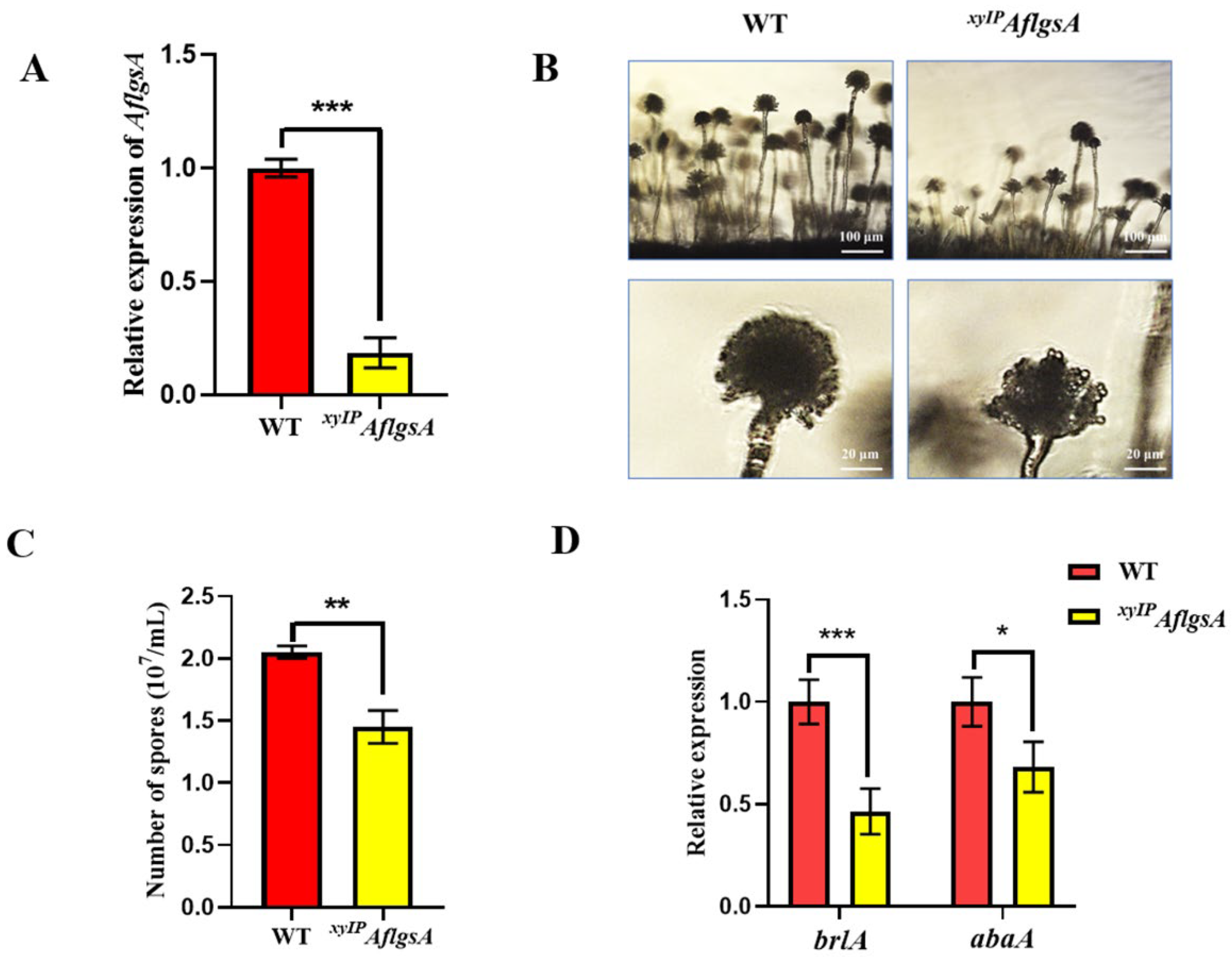
2.4. AflGsA Is Important for Conidia Development of A. flavus
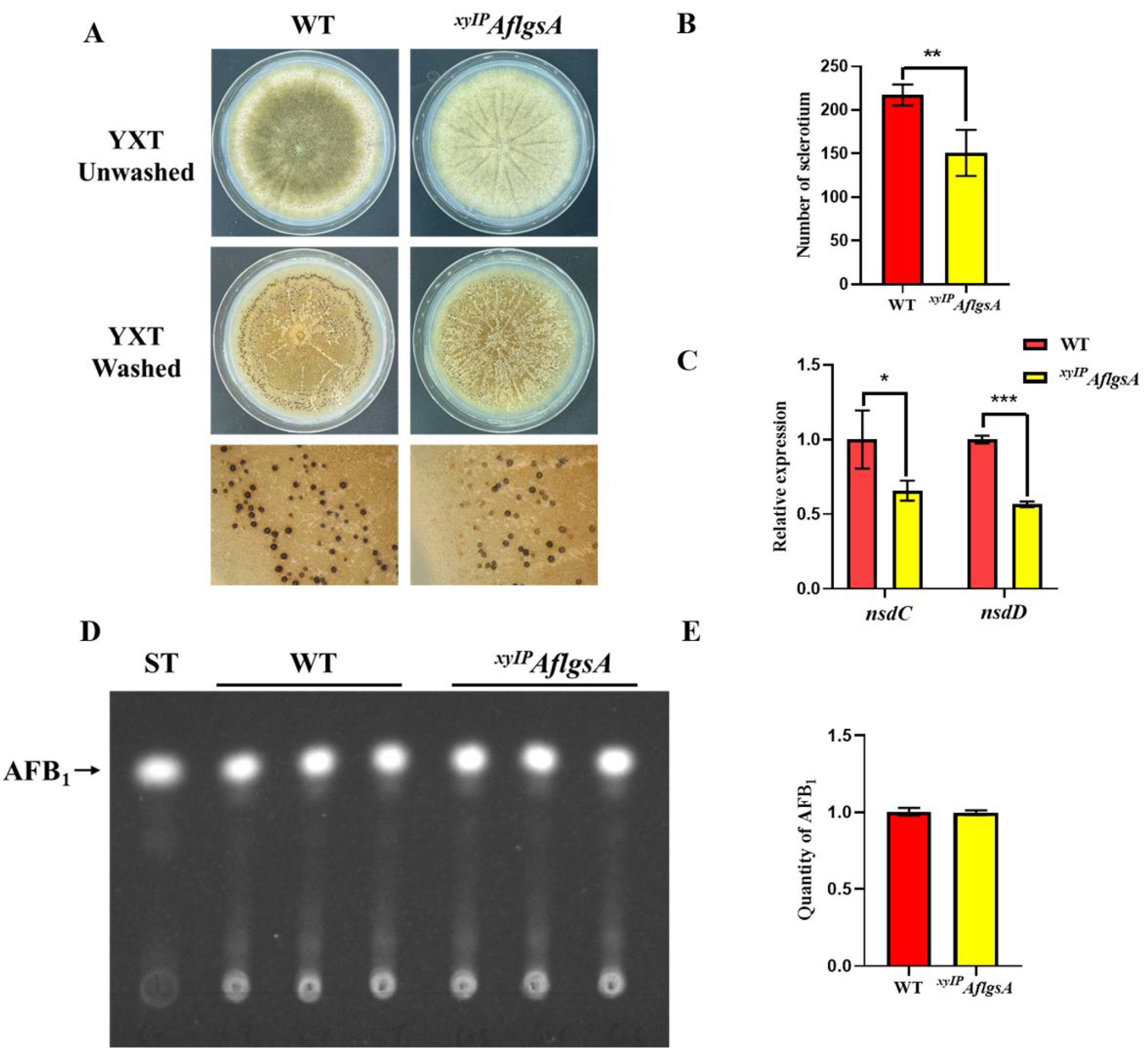
2.5. AflGsA Contributes to the Production of Sclerotia but Not to Toxin Synthesis in A. flavus
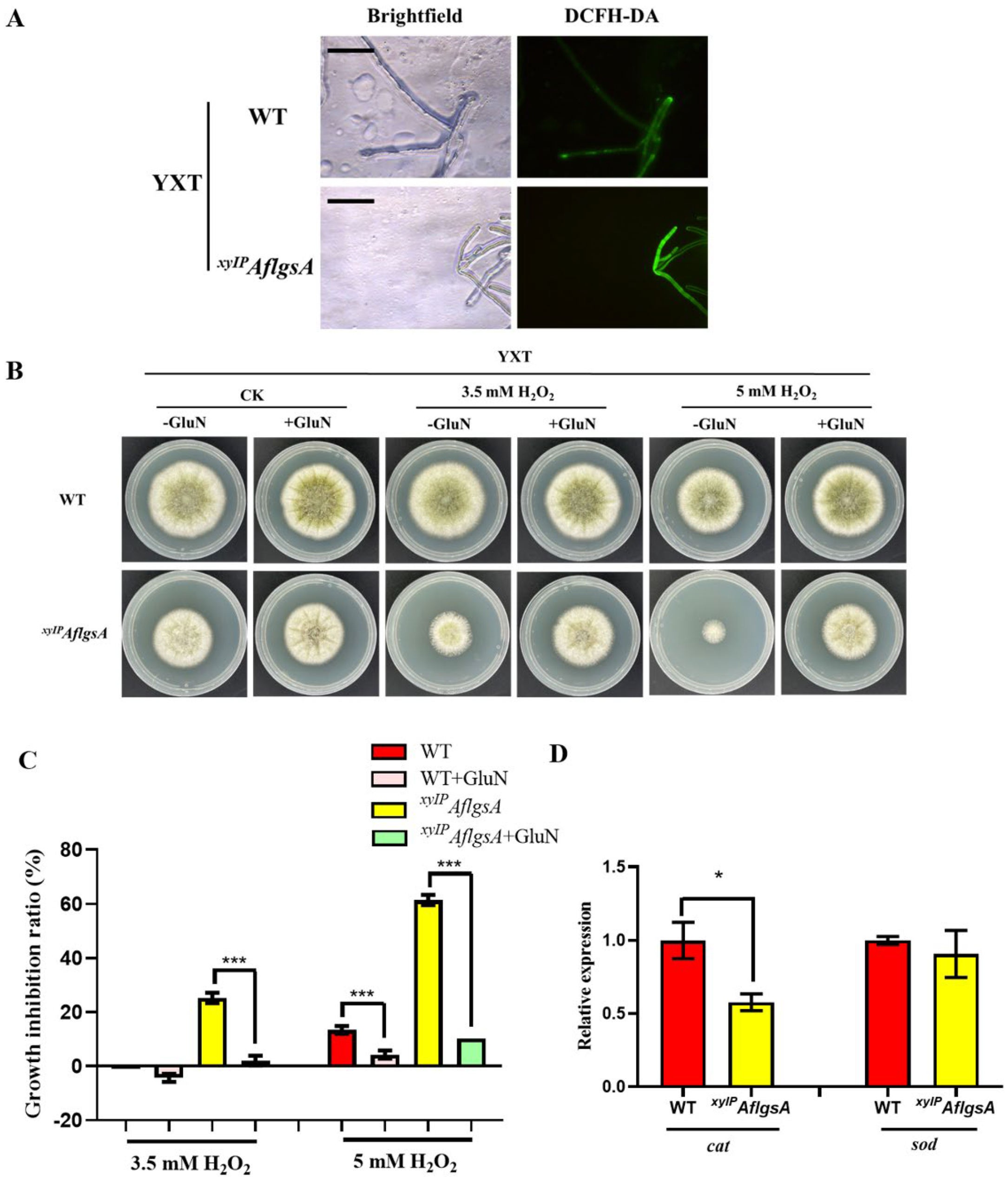
2.6. AflGsA Balances ROS and Resists Oxidative Stress in A. flavus
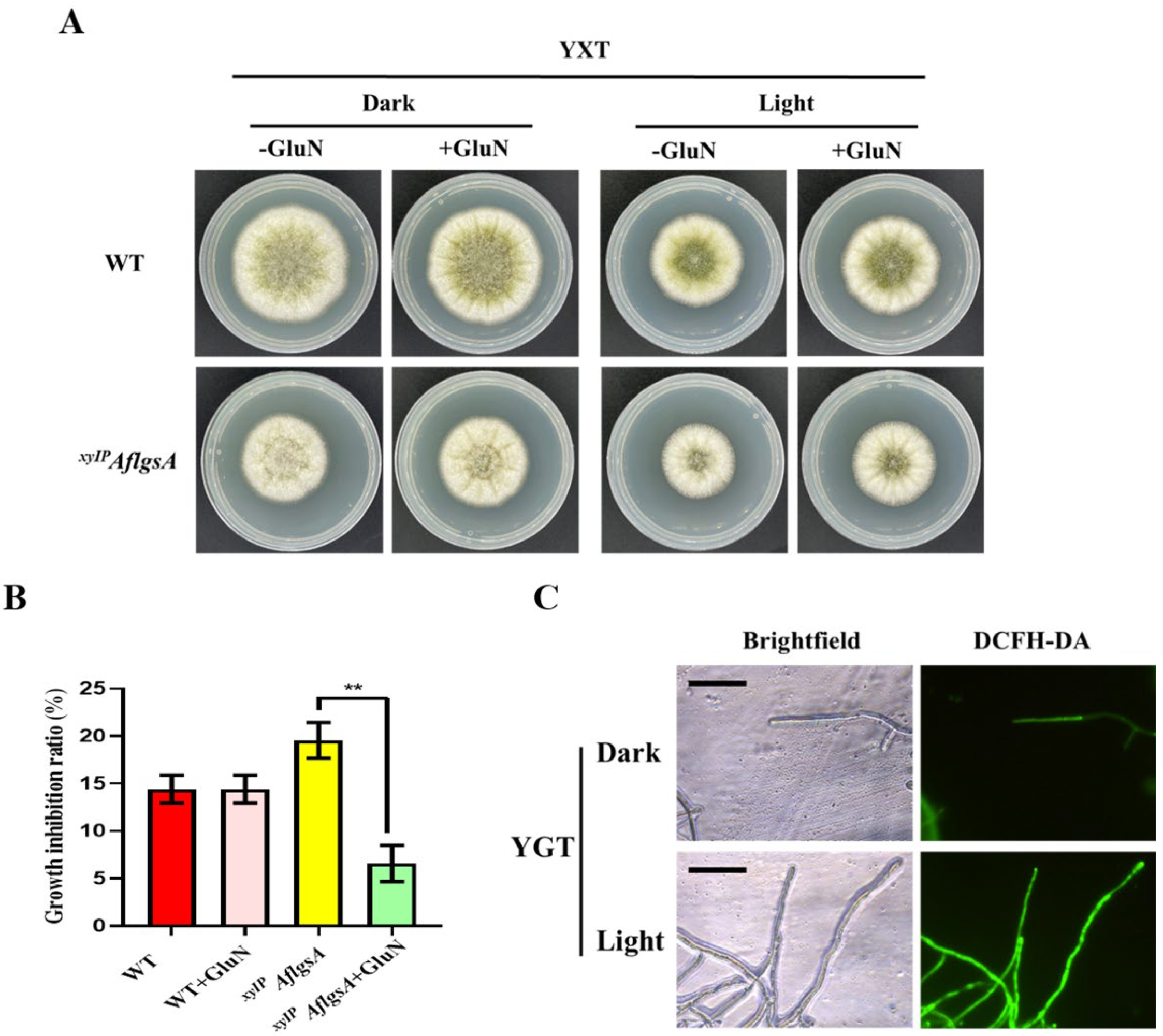
2.7. AflGsA Is Involved in Light Signaling Pathways in A. flavus
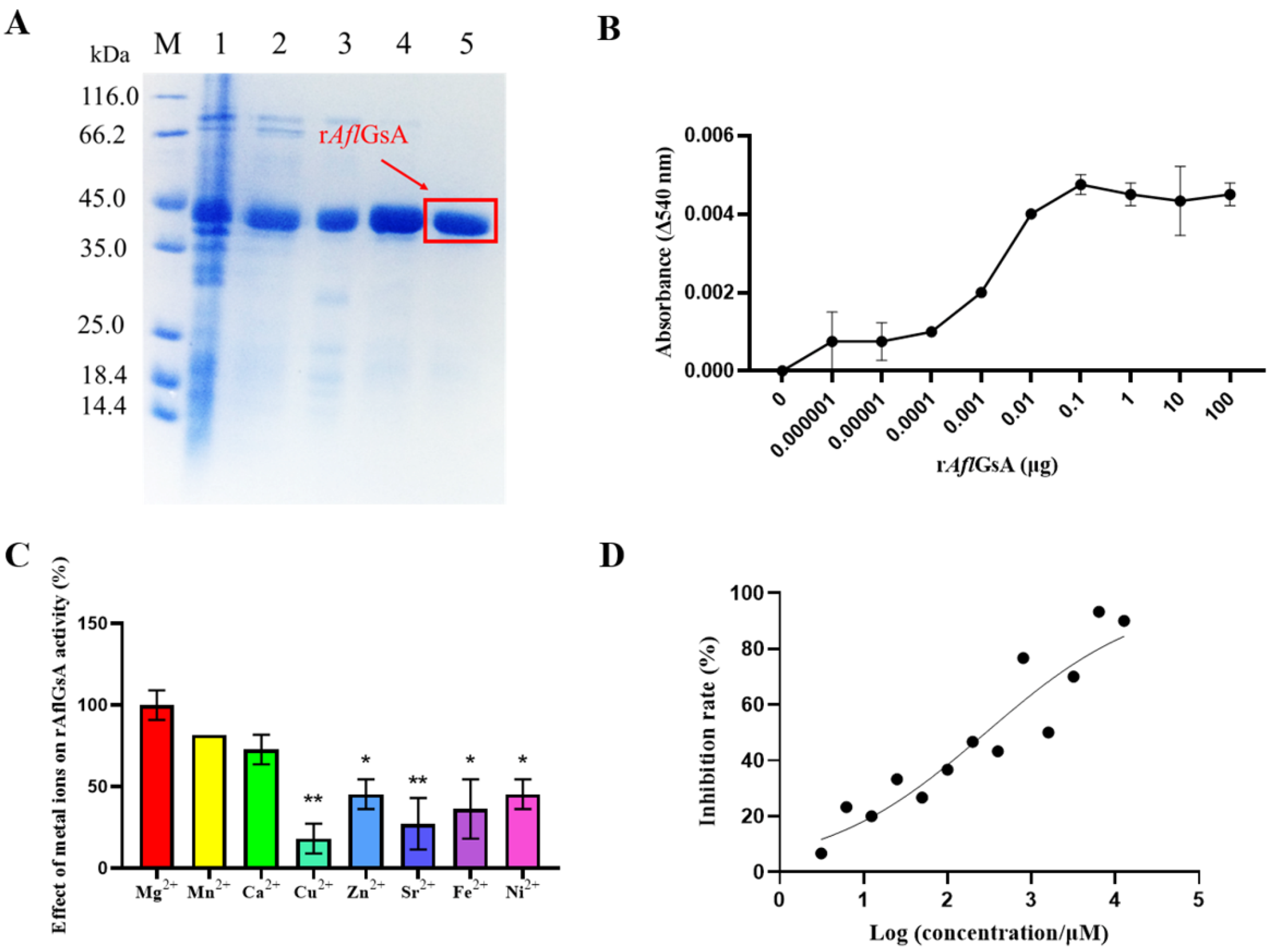
2.8. Recombinant AflGsA (rAflGsA) Has Glutamine Synthetase Activity
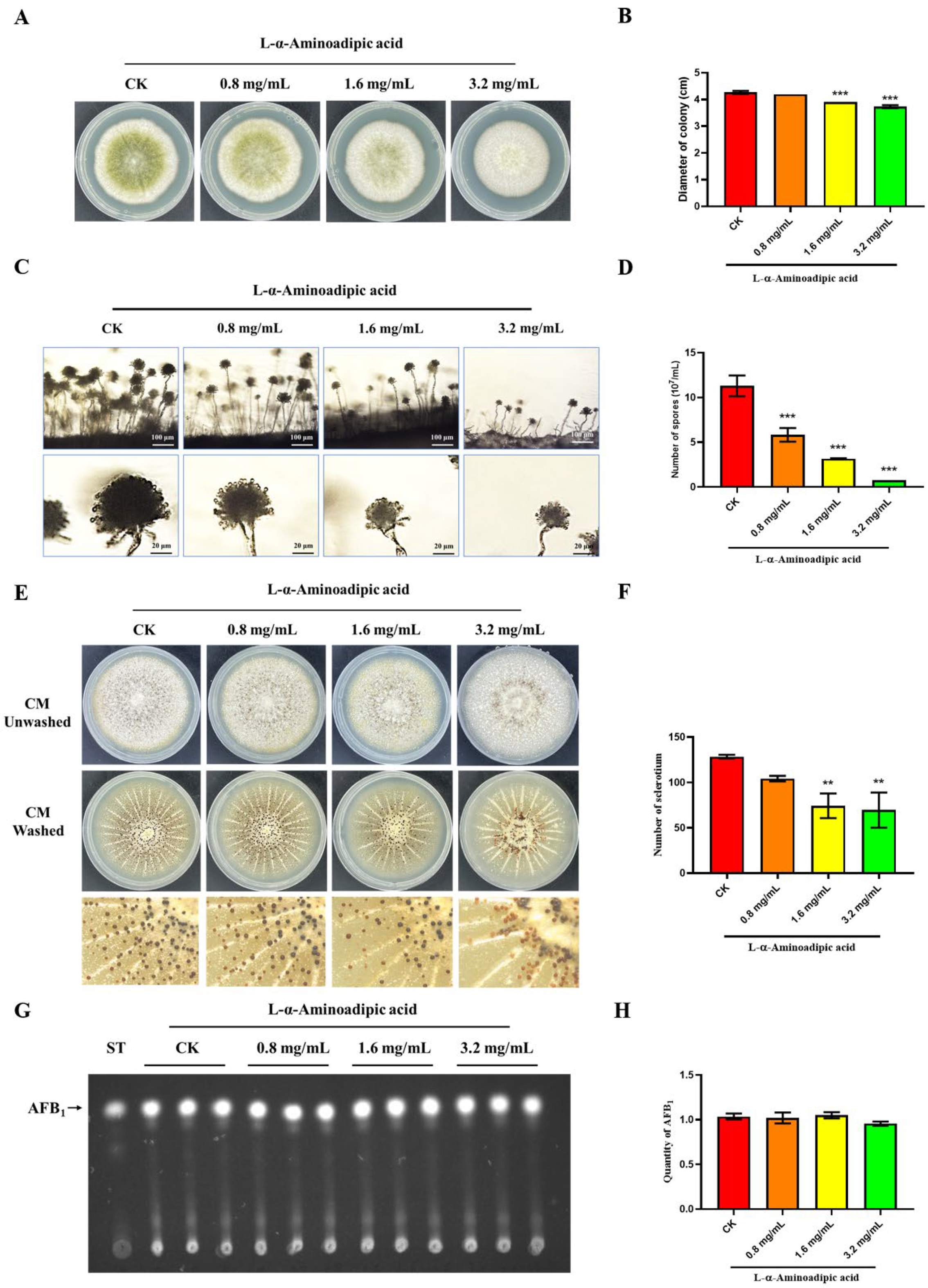
2.9. L-α-Aminoadipic Acid Is a Potential Inhibitor for rAflGsA and A. flavus
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strains and Culture Conditions
5.2. Sequence Analysis
5.3. Construction and Identification of Mutant Strain
5.4. Analysis of the Growth, Conidial Production, and Sclerotia Formation of A. flavus
5.5. Detection of ROS in A. flavus
5.6. Quantitative Real-Time PCR
5.7. Purification of Recombinant rAflGsA Protein and Determination of Enzyme Activity
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bennett, J.W.; Klich, M.A. Aspergillus: Biology and industrial applications. Biotechnology 1992, 23, 1. [Google Scholar]
- Samson, R.A. Current taxonomic schemes of the genus Aspergillus and its teleomorphs. Biotechnology 1992, 23, 355–390. [Google Scholar]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Manavathu, E.K.; Chandrasekar, P.H. Aspergillus flavus: An emerging non-fumigatus Aspergillus species of significance. Mycoses 2009, 52, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Kousha, M.; Tadi, R.; Soubani, A.O. Pulmonary aspergillosis: A clinical review. Eur. Respir. Rev. Off. J. Eur. Respir. Soc. 2011, 20, 156–174. [Google Scholar] [CrossRef]
- Mehl, H.L.; Jaime, R.; Callicott, K.A.; Probst, C.; Garber, N.P.; Ortega-Beltran, A.; Grubisha, L.C.; Cotty, P.J. Aspergillus flavus diversity on crops and in the environment can be exploited to reduce aflatoxin exposure and improve health. Ann. N. Y. Acad. Sci. 2012, 1273, 7–17. [Google Scholar] [CrossRef]
- Lim, C.W.; Yoshinari, T.; Layne, J.; Chan, S.H. Multi-mycotoxin screening reveals separate occurrence of aflatoxins and ochratoxin a in Asian rice. J. Agric. Food Chem. 2015, 63, 3104–3113. [Google Scholar] [CrossRef]
- Leung, M.C.; Díaz-Llano, G.; Smith, T.K. Mycotoxins in pet food: A review on worldwide prevalence and preventative strategies. J. Agric. Food Chem. 2006, 54, 9623–9635. [Google Scholar] [CrossRef]
- Choi, K.C.; Chung, W.T.; Kwon, J.K.; Yu, J.Y.; Jang, Y.S.; Park, S.M.; Lee, S.Y.; Lee, J.C. Inhibitory effects of quercetin on aflatoxin B1-induced hepatic damage in mice. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2010, 48, 2747–2753. [Google Scholar] [CrossRef]
- Lam, H.M.; Coschigano, K.T.; Oliveira, I.C.; Melo-Oliveira, R.; Coruzzi, G.M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef]
- Eisenberg, D.; Gill, H.S.; Pfluegl, G.M.; Rotstein, S.H. Structure-function relationships of glutamine synthetases. Biochim. Biophys. Acta 2000, 1477, 122–145. [Google Scholar] [CrossRef]
- Christa, L.; Simon, M.T.; Flinois, J.P.; Gebhardt, R.; Brechot, C.; Lasserre, C. Overexpression of glutamine synthetase in human primary liver cancer. Gastroenterology 1994, 106, 1312–1320. [Google Scholar] [CrossRef]
- Caizzi, R.; Bozzetti, M.P.; Caggese, C.; Ritossa, F. Homologous nuclear genes encode cytoplasmic and mitochondrial glutamine synthetase in Drosophila melanogaster. J. Mol. Biol. 1990, 212, 17–26. [Google Scholar] [CrossRef]
- Avisar, N.; Shiftan, L.; Ben-Dror, I.; Havazelet, N.; Vardimon, L. A silencer element in the regulatory region of glutamine synthetase controls cell type-specific repression of gene induction by glucocorticoids. J. Biol. Chem. 1999, 274, 11399–11407. [Google Scholar] [CrossRef]
- Teichert, S.; Schönig, B.; Richter, S.; Tudzynski, B. Deletion of the Gibberella fujikuroi glutamine synthetase gene has significant impact on transcriptional control of primary and secondary metabolism. Mol. Microbiol. 2004, 53, 1661–1675. [Google Scholar] [CrossRef]
- Kichey, T.; Le Gouis, J.; Sangwan, B.; Hirel, B.; Dubois, F. Changes in the cellular and subcellular localization of glutamine synthetase and glutamate dehydrogenase during flag leaf senescence in wheat (Triticum aestivum L.). Plant Cell Physiol. 2005, 46, 964–974. [Google Scholar] [CrossRef]
- Takano, H.K.; Beffa, R.; Preston, C.; Westra, P.; Dayan, F.E. A novel insight into the mode of action of glufosinate: How reactive oxygen species are formed. Photosynth. Res. 2020, 144, 361–372. [Google Scholar] [CrossRef]
- Fu, S.; Li, Z.; Xiao, L.; Hu, W.; Zhang, L.; Xie, B.; Zhou, Q.; He, J.; Qiu, Y.; Wen, M.; et al. Glutamine Synthetase Promotes Radiation Resistance via Facilitating Nucleotide Metabolism and Subsequent DNA Damage Repair. Cell Rep. 2019, 28, 1136–1143.e1134. [Google Scholar] [CrossRef]
- Kim, C.; Kültz, D. An osmolality/salinity-responsive enhancer 1 (OSRE1) in intron 1 promotes salinity induction of tilapia glutamine synthetase. Sci. Rep. 2020, 10, 12103. [Google Scholar] [CrossRef]
- Sasaki, Y.; Kojima, A.; Shibata, Y.; Mitsuzawa, H. Filamentous invasive growth of mutants of the genes encoding ammonia-metabolizing enzymes in the fission yeast Schizosaccharomyces pombe. PLoS ONE 2017, 12, e0186028. [Google Scholar] [CrossRef]
- Margelis, S.; D’Souza, C.; Small, A.J.; Hynes, M.J.; Adams, T.H.; Davis, M.A. Role of glutamine synthetase in nitrogen metabolite repression in Aspergillus nidulans. J. Bacteriol. 2001, 183, 5826–5833. [Google Scholar] [CrossRef] [PubMed]
- Takano, H.K.; Dayan, F.E. Biochemical Basis for the Time-of-Day Effect on Glufosinate Efficacy against Amaranthus palmeri. Plants 2021, 10, 2021. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Herrero, V.; Payá, G.; Bautista, V.; Vegara, A.; Cortés-Molina, M.; Camacho, M.; Esclapez, J.; Bonete, M.J. Essentiality of the glnA gene in Haloferax mediterranei: Gene conversion and transcriptional analysis. Extrem. Life Extrem. Cond. 2020, 24, 433–446. [Google Scholar] [CrossRef]
- Aron, O.; Wang, M.; Lin, L.; Batool, W.; Lin, B.; Shabbir, A.; Wang, Z.; Tang, W. MoGLN2 Is Important for Vegetative Growth, Conidiogenesis, Maintenance of Cell Wall Integrity and Pathogenesis of Magnaporthe oryzae. J. Fungi 2021, 7, 463. [Google Scholar] [CrossRef] [PubMed]
- Almassy, R.J.; Janson, C.A.; Hamlin, R.; Xuong, N.H.; Eisenberg, D. Novel subunit—Subunit interactions in the structure of glutamine synthetase. Nature 1986, 323, 304–309. [Google Scholar] [CrossRef]
- Valentine, R.C.; Shapiro, B.M.; Stadtman, E.R. Regulation of glutamine synthetase. XII. Electron microscopy of the enzyme from Escherichia coli. Biochemistry 1968, 7, 2143–2152. [Google Scholar] [CrossRef]
- van Rooyen, J.M.; Abratt, V.R.; Belrhali, H.; Sewell, T. Crystal structure of Type III glutamine synthetase: Surprising reversal of the inter-ring interface. Structure 2011, 19, 471–483. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, W.; Yu, S.; Ni, K.; She, G.; Ye, X.; Xing, Q.; Zhao, J.; Huang, C. Assembly status transition offers an avenue for activity modulation of a supramolecular enzyme. eLife 2021, 10, e72535. [Google Scholar] [CrossRef]
- Woolfolk, C.A.; Stadtman, E.R. Regulation of glutamine synthetase: III. Cumulative feedback inhibition of glutamine synthetase from Escherichia coli. Arch. Biochem. Biophys. 1967, 118, 736–755. [Google Scholar] [CrossRef]
- Lan, H.; Wu, L.; Sun, R.; Keller, N.P.; Yang, K.; Ye, L.; He, S.; Zhang, F.; Wang, S. The HosA histone deacetylase regulates aflatoxin biosynthesis through direct regulation of aflatoxin cluster genes. Mol. Plant-Microbe Interact. 2019, 32, 1210–1228. [Google Scholar] [CrossRef]
- McBean, G.J. Inhibition of the glutamate transporter and glial enzymes in rat striatum by the gliotoxin, alpha aminoadipate. Br. J. Pharmacol. 1994, 113, 536–540. [Google Scholar] [CrossRef]
- Bao, A.; Zhao, Z.; Ding, G.; Shi, L.; Xu, F.; Cai, H. The Stable Level of Glutamine synthetase 2 plays an important role in Rice growth and in Carbon-Nitrogen metabolic balance. Int. J. Mol. Sci. 2015, 16, 12713–12736. [Google Scholar] [CrossRef]
- Shatters, R.G.; Liu, Y.; Kahn, M.L. Isolation and characterization of a novel glutamine synthetase from Rhizobium meliloti. J. Biol. Chem. 1993, 268, 469–475. [Google Scholar] [CrossRef]
- Aguirre, J.; Ríos-Momberg, M.; Hewitt, D.; Hansberg, W. Reactive oxygen species and development in microbial eukaryotes. Trends Microbiol. 2005, 13, 111–118. [Google Scholar] [CrossRef]
- Gessler, N.N.; Aver’yanov, A.A.; Belozerskaya, T.A. Reactive oxygen species in regulation of fungal development. Biochem. Biokhimiia 2007, 72, 1091–1109. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Robles-Rengel, R.; Florencio, F.J.; Muro-Pastor, M.I. Redox interference in nitrogen status via oxidative stress is mediated by 2-oxoglutarate in cyanobacteria. New Phytol. 2019, 224, 216–228. [Google Scholar] [CrossRef]
- Montanini, B.; Betti, M.; Márquez, A.J.; Balestrini, R.; Bonfante, P.; Ottonello, S. Distinctive properties and expression profiles of glutamine synthetase from a plant symbiotic fungus. Biochem. J. 2003, 373, 357–368. [Google Scholar] [CrossRef]
- Sakakibara, H.; Shimizu, H.; Hase, T.; Yamazaki, Y.; Takao, T.; Shimonishi, Y.; Sugiyama, T. Molecular identification and characterization of cytosolic isoforms of glutamine synthetase in maize roots. J. Biol. Chem. 1996, 271, 29561–29568. [Google Scholar] [CrossRef]
- Gong, C.; You, X.; Zhang, S.; Xue, D. Functional analysis of a glutamine biosynthesis protein from a psychrotrophic Bacterium, Cryobacterium soli GCJ02. Indian J. Microbiol. 2020, 60, 153–159. [Google Scholar] [CrossRef]
- Zhao, W.; Yang, J.; Tian, Y.; Fu, X.; Zhu, B.; Xue, Y.; Gao, J.; Han, H.J.; Peng, R.; Yao, Q.H. Expression, purification, and characterization of recombinant mangrove glutamine synthetase. Mol. Biol. Rep. 2014, 41, 7575–7583. [Google Scholar] [CrossRef] [PubMed]
- Huck, S.; Grass, F.; Hörtnagl, H. The glutamate analogue alpha-aminoadipic acid is taken up by astrocytes before exerting its gliotoxic effect in vitro. J. Neurosci. Off. J. Soc. Neurosci. 1984, 4, 2650–2657. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Q.; Ungerstedt, U.; Schwarcz, R. L-alpha-aminoadipic acid as a regulator of kynurenic acid production in the hippocampus: A microdialysis study in freely moving rats. Eur. J. Pharmacol. 1995, 281, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, S.; Nie, X.; Yang, K.; Xu, P.; Wang, X.; Liu, M.; Yang, Y.; Chen, Z.; Wang, S. Molecular and structural basis of nucleoside diphosphate kinase-mediated regulation of spore and sclerotia development in the fungus Aspergillus flavus. J. Biol. Chem. 2019, 294, 12415–12431. [Google Scholar] [CrossRef]
- Zadra, I.; Abt, B.; Parson, W.; Haas, H. xylP promoter-based expression system and its use for antisense downregulation of the Penicillium chrysogenum nitrogen regulator NRE. Appl. Environ. Microbiol. 2000, 66, 4810–4816. [Google Scholar] [CrossRef]
- Chang, P.K.; Scharfenstein, L.L.; Wei, Q.; Bhatnagar, D. Development and refinement of a high-efficiency gene-targeting system for Aspergillus flavus. J. Microbiol. Methods 2010, 81, 240–246. [Google Scholar] [CrossRef]
- Zhang, F.; Geng, L.; Deng, J.; Huang, L.; Zhong, H.; Xin, S.; Fasoyin, O.E.; Wang, S. The MAP kinase AflSlt2 modulates aflatoxin biosynthesis and peanut infection in the fungus Aspergillus flavus. Int. J. Food Microbiol. 2020, 322, 108576. [Google Scholar] [CrossRef]
- Yang, K.; Liu, Y.; Wang, S.; Wu, L.; Xie, R.; Lan, H.; Fasoyin, O.E.; Wang, Y.; Wang, S. Cyclase-associated protein Cap with multiple domains contributes to mycotoxin biosynthesis and fungal virulence in Aspergillus flavus. J. Agric. Food Chem. 2019, 67, 4200–4213. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Lin, R.; Tumukunde, E.; Zeng, W.; Bao, Q.; Wang, S.; Wang, Y. Glutamine Synthetase Contributes to the Regulation of Growth, Conidiation, Sclerotia Development, and Resistance to Oxidative Stress in the Fungus Aspergillus flavus. Toxins 2022, 14, 822. https://doi.org/10.3390/toxins14120822
Wang S, Lin R, Tumukunde E, Zeng W, Bao Q, Wang S, Wang Y. Glutamine Synthetase Contributes to the Regulation of Growth, Conidiation, Sclerotia Development, and Resistance to Oxidative Stress in the Fungus Aspergillus flavus. Toxins. 2022; 14(12):822. https://doi.org/10.3390/toxins14120822
Chicago/Turabian StyleWang, Sen, Ranxun Lin, Elisabeth Tumukunde, Wanlin Zeng, Qian Bao, Shihua Wang, and Yu Wang. 2022. "Glutamine Synthetase Contributes to the Regulation of Growth, Conidiation, Sclerotia Development, and Resistance to Oxidative Stress in the Fungus Aspergillus flavus" Toxins 14, no. 12: 822. https://doi.org/10.3390/toxins14120822
APA StyleWang, S., Lin, R., Tumukunde, E., Zeng, W., Bao, Q., Wang, S., & Wang, Y. (2022). Glutamine Synthetase Contributes to the Regulation of Growth, Conidiation, Sclerotia Development, and Resistance to Oxidative Stress in the Fungus Aspergillus flavus. Toxins, 14(12), 822. https://doi.org/10.3390/toxins14120822