The Potential of Alternaria Toxins Production by A. alternata in Processing Tomatoes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
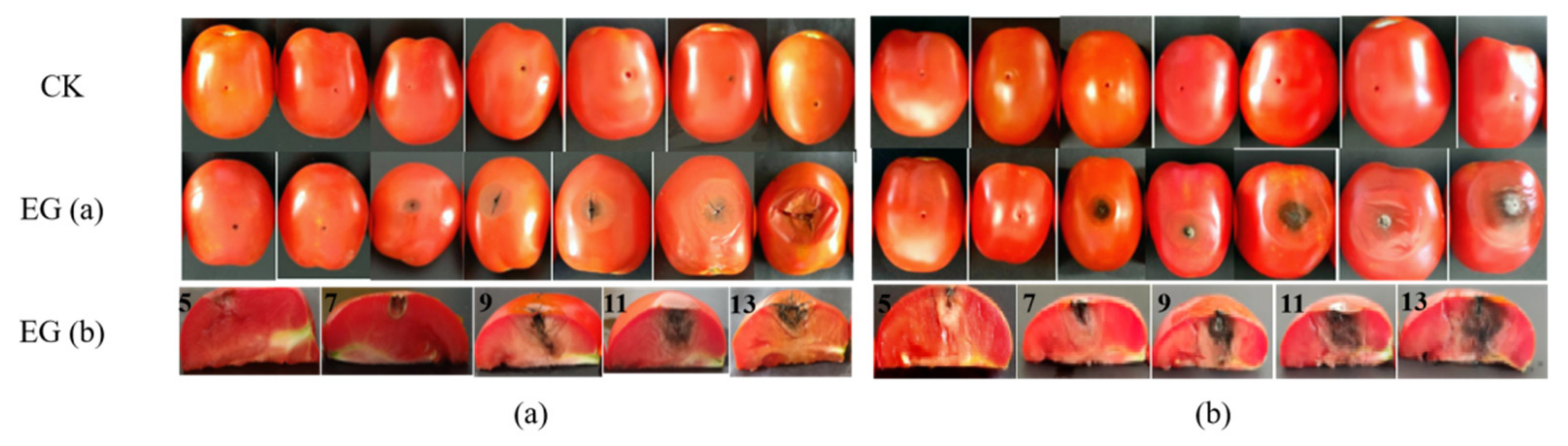
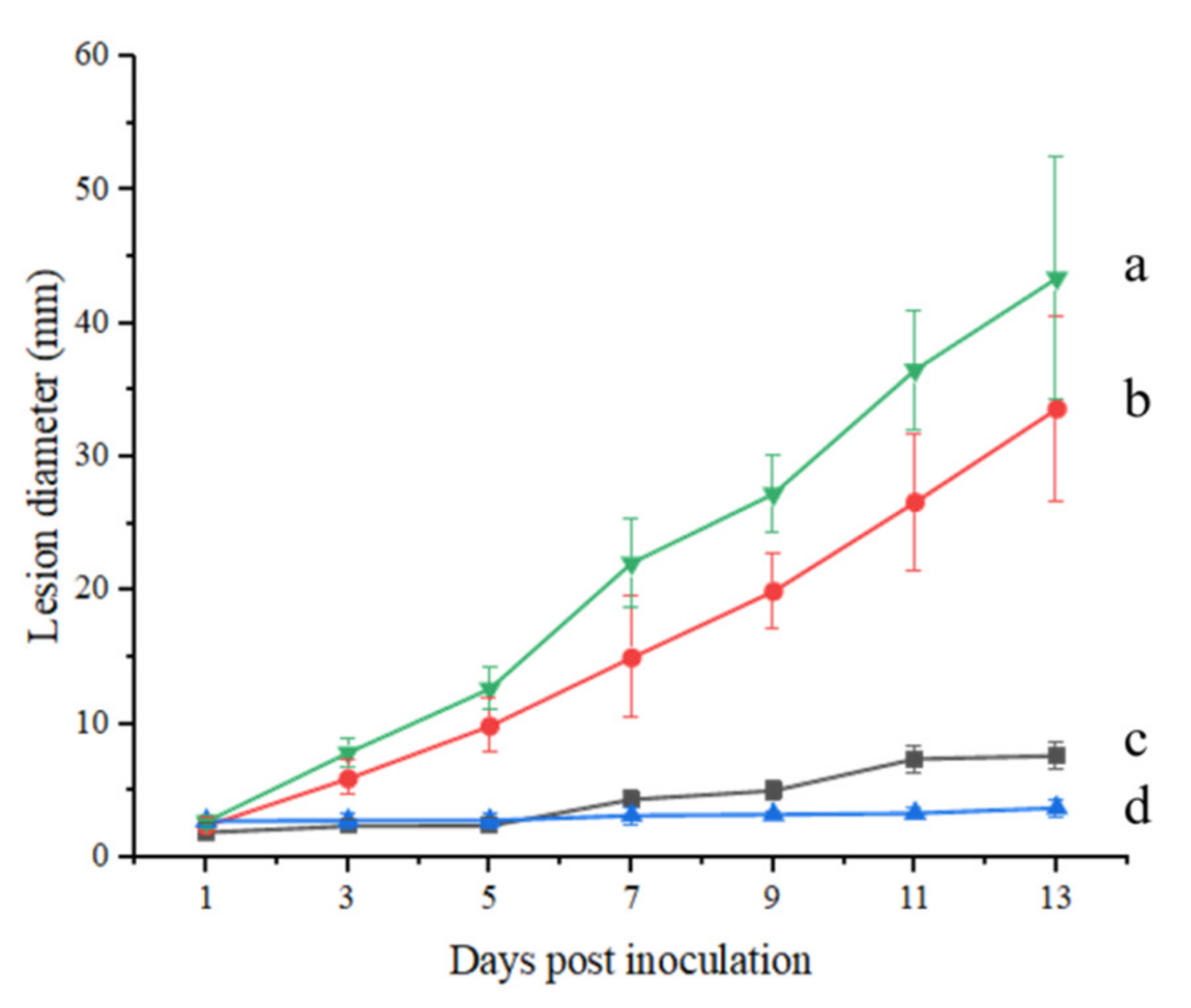
2.1. The Lesion Diameter Affected by A. alternata
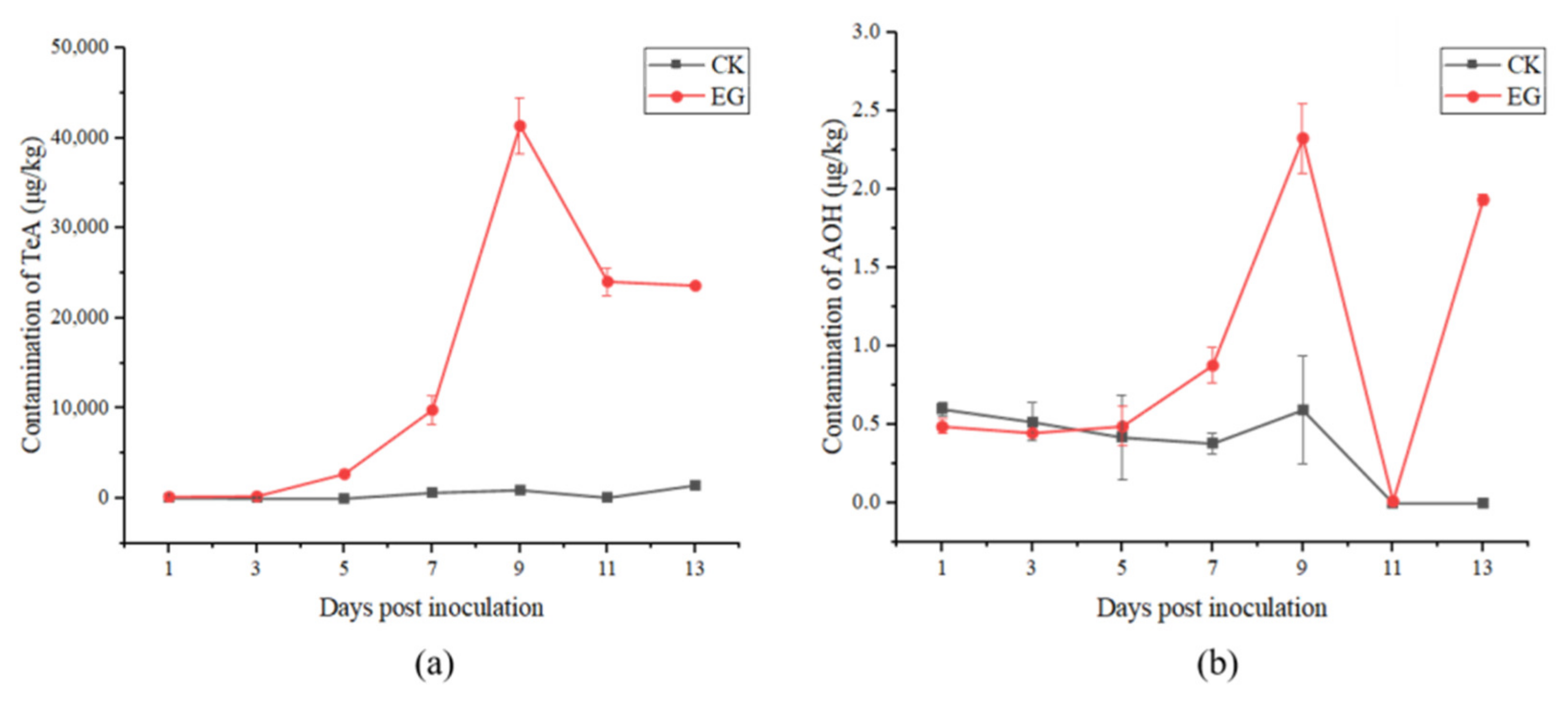
2.2. Alternaria Toxins Produced in Processing Tomatoes in the Field Experiment
2.2.1. Analysis of Alternaria Toxins
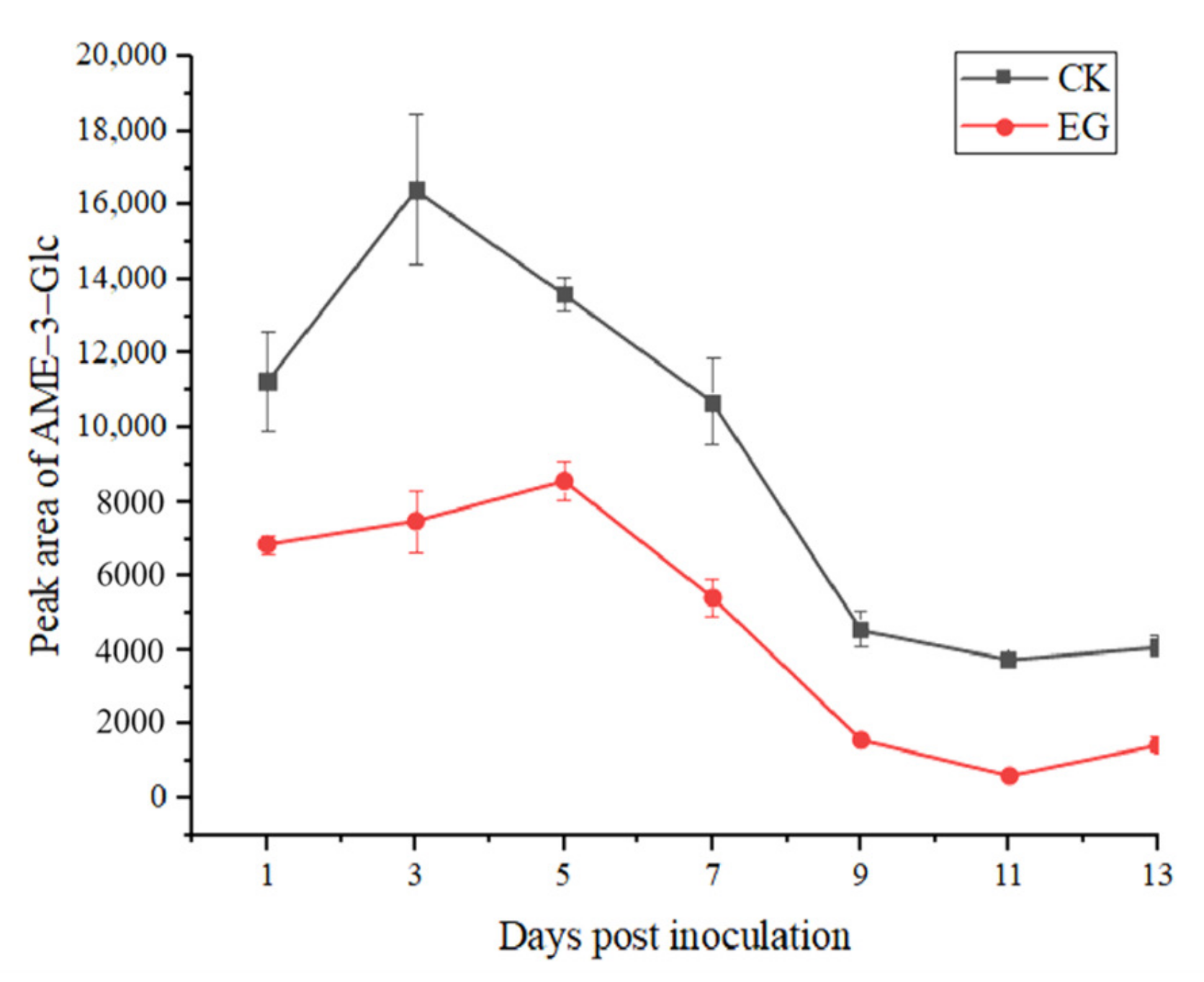
2.2.2. Analysis of Conjugated Alternaria Toxins
2.3. Alternaria Toxins Produced in Processing Tomato in In Vivo Experiment
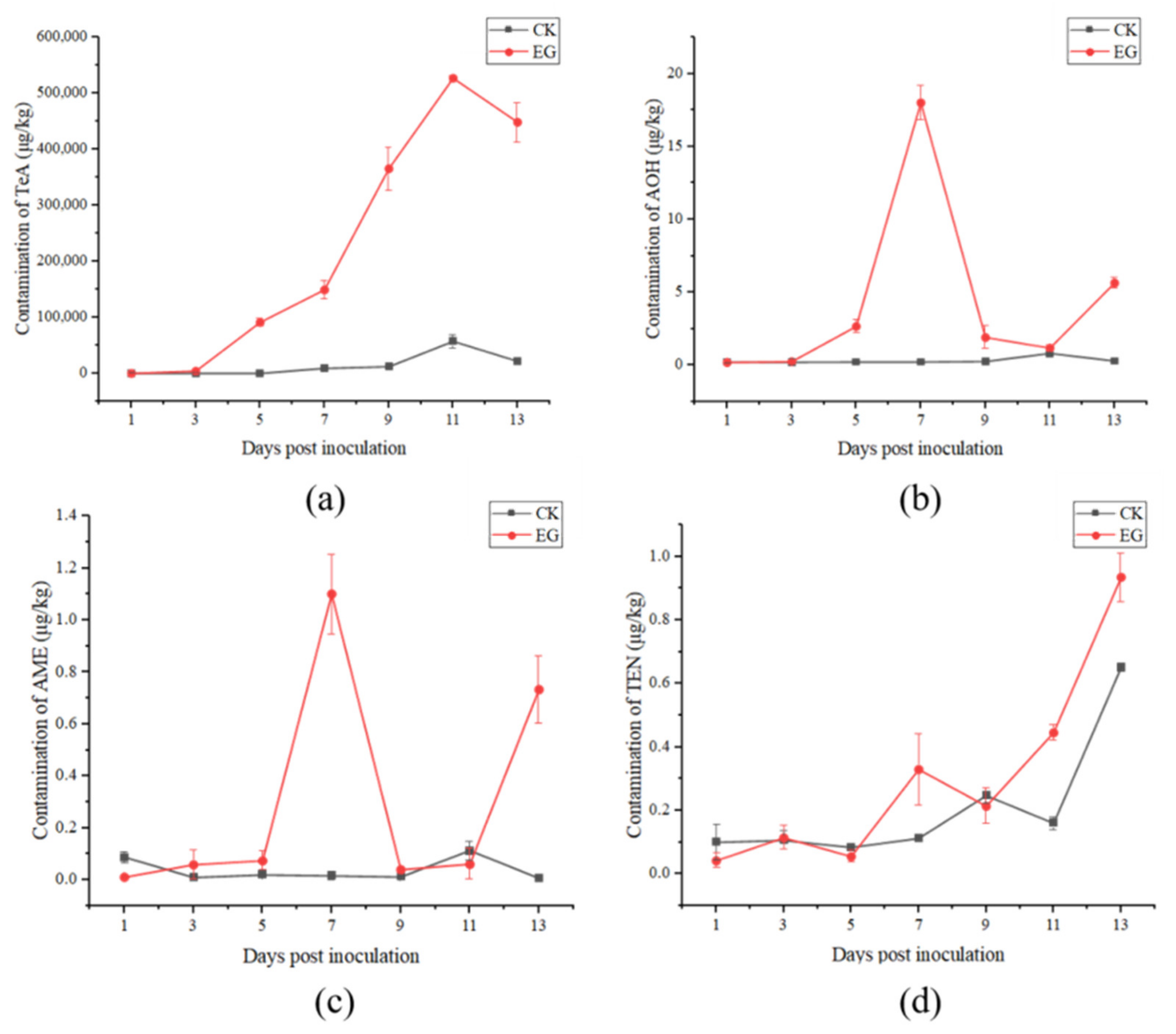
2.3.1. Analysis of Alternaria Toxins
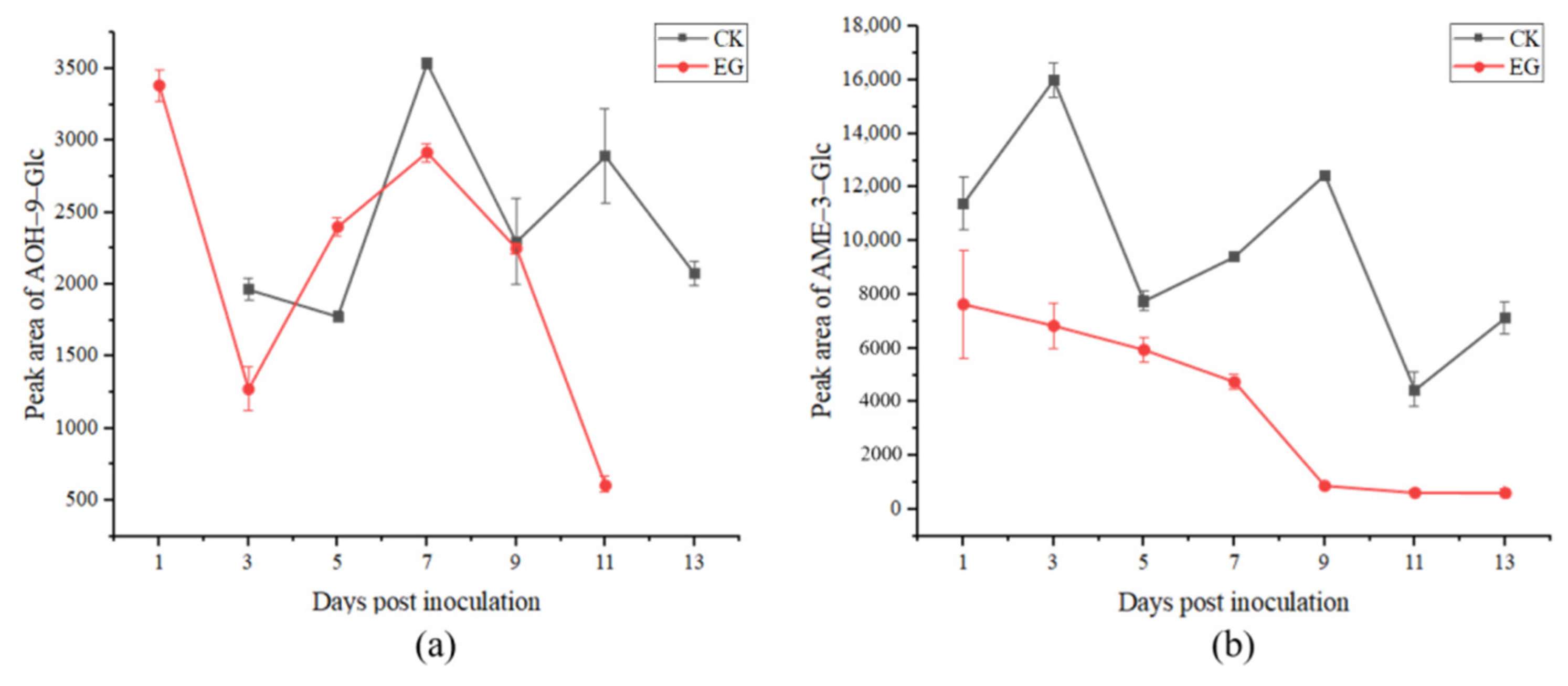
2.3.2. Analysis of Conjugated Alternaria Toxins
3. Discussion
3.1. Virulence of A. alternata in the Inoculation of Tomatoes
3.2. Varying Potential of Alternaria Toxins Production between Field and In Vivo Experiments
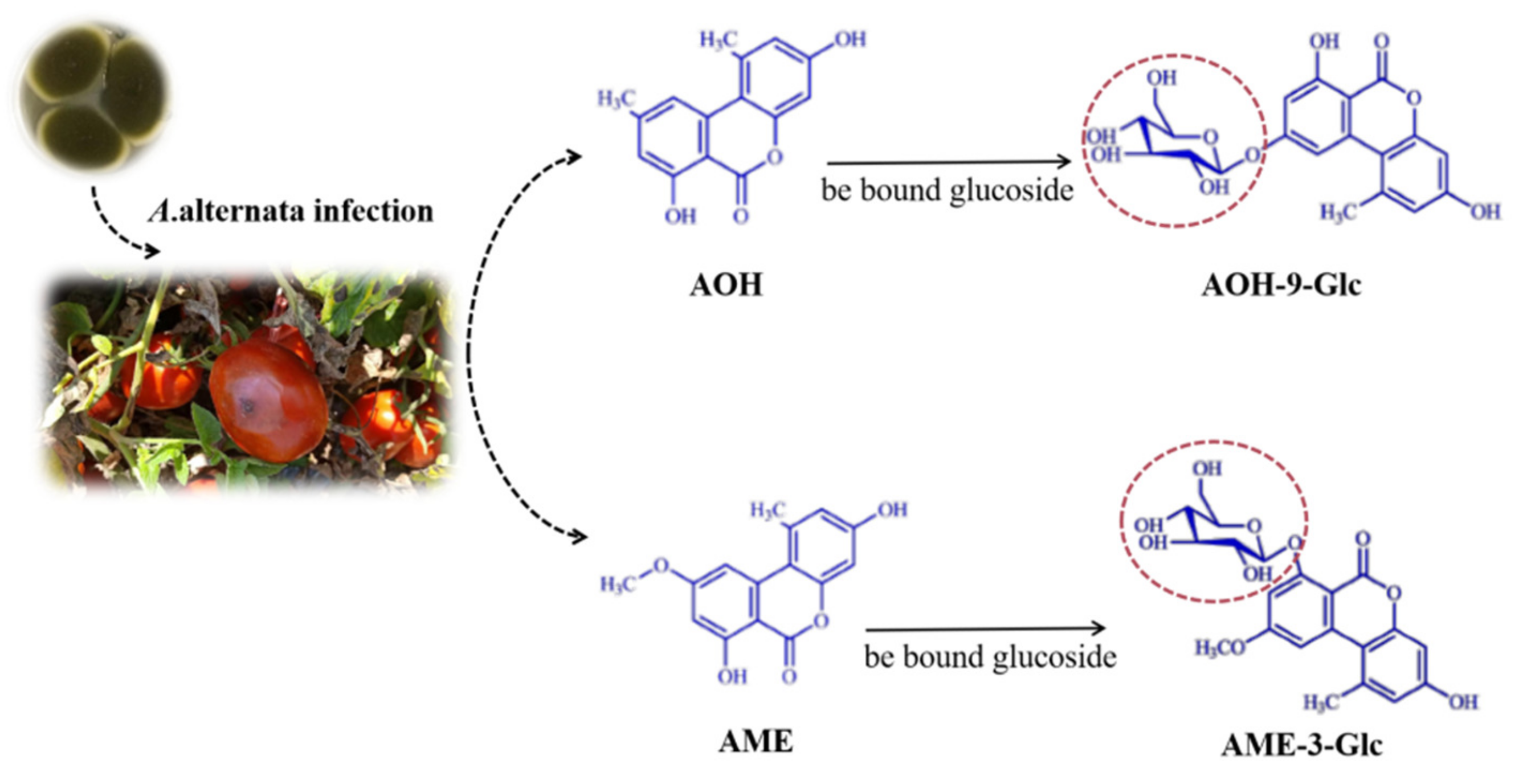
3.3. The Production of Conjugated Mycotoxins
4. Conclusions
5. Material and Methods
5.1. Chemical and Reagents
5.2. Instruments
5.3. Tomato, A. alternata and Spore Suspension
5.4. Field Experiment
5.5. In Vivo Experiment
5.6. Lesion Diameter Measurement
5.7. Extraction of Alternaria Toxins
5.8. Alternaria Toxins Detection
5.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prusky, D.; Eshel, D.; Kobiler, I.; Yakoby, N.; Moualem, D.B.; Ackerman, M.; Zuthji, Y.; Arie, R.B. Postharvest chlorine treatments for the control of the persimmon black spot disease caused by Alternaria alternata. Postharvest Biol. Technol. 2001, 22, 271–277. [Google Scholar] [CrossRef]
- Smith, J.E.; Mengesha, B.; Tang, H.; Mengiste, T.; Bluhm, B.H. Resistance to Botrytis cinerea in Solanum lycopersicoides involves widespread transcriptional reprogramming. BMC Genom. 2014, 15, 334. [Google Scholar] [CrossRef] [PubMed]
- Derbalah, A.; Shenashen, M.; Hamza, A.; Mohamed, A.; Safty, S.E. Antifungal activity of fabricated mesoporous silica nanoparticles against early blight of tomato. Egypt. J. Basic Appl. Sci. 2018, 5, 145–150. [Google Scholar] [CrossRef]
- Pearson, R.C. Factors affecting the occurrence and severity of blackmold of ripe tomato fruit caused by Alternaria alternata [Fungus diseases]. Phytopathology 1975, 65, 1352. [Google Scholar] [CrossRef]
- Troncoso, R.R.; Tiznado, H.M.E. Chapter 5—Alternaria alternata (Black Rot, Black Spot). Postharvest Decay. 2014, 147–187. [Google Scholar] [CrossRef]
- Prusky, D.; Alkan, N.; Mengiste, T.; Fluhr, R. Quiescent and necrotrophic lifestyle choice during postharvest disease development. Annu. Rev. Phytopathol. 2013, 51, 155–176. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef]
- Ozcelik, S.; Ozcelik, N.; Beuchat, L.R. Toxin production by Alternaria alternata in tomatoes and apples stored under various conditions and quantitation of the toxins by high-performance liquid chromatography. Int. J. Food Microbiol. 1990, 11, 187–194. [Google Scholar] [CrossRef]
- Mukesh, M.; Andleeb, Z.; Dubey, M.K.; Mohd, A.; Gupta, V.K.; Upadhyay, R.S. Comparative evaluation of biochemical changes in tomato (Lycopersicon esculentum mill.) infected by Alternaria alternata and its toxic metabolites (TeA, AOH, and AME). Front. Plant Sci. 2016, 7, 1408. [Google Scholar]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; Vries, M.D.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef]
- Patriarca, A.; Azcarate, M.P.; Terminiello, L.; Pinto, V.F. Mycotoxin production by Alternaria strains isolated from Argentinean wheat. Int. J. Food Microbiol. 2007, 119, 219–222. [Google Scholar] [CrossRef]
- Dall, A.C.; Cirlini, M.; Falavigna, C. Chapter three-mycotoxins from Alternaria: Toxicological implications. Adv. Mol. Toxicol. 2014, 8, 107–121. [Google Scholar]
- Chain, E. Scientific opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 2407. [Google Scholar]
- Michele, S. Recent advances on Alternaria mycotoxins. Curr. Opin. Food Sci. 2017, 17, 57–61. [Google Scholar]
- Rychlik, M.; Humpf, H.U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef]
- Pero, R.W.; Posner, H.; Blois, M.; Harvan, D.; Spalding, J.W. Toxicity of metabolites produced by the “Alternaria”. Environ. Health Persp. 1973, 4, 87–94. [Google Scholar] [CrossRef]
- Kang, Y.; Feng, H.W.; Zhang, J.X.; Chen, S.G.; Valverde, B.E.; Qiang, S. TeA is a key virulence factor for Alternaria alternata (Fr.) Keissler infection of its host. Plant Physiol. Biochem. 2017, 115, 73–82. [Google Scholar] [CrossRef]
- Vejdovszky, K.; Warth, B.; Sulyok, M.; Marko, D. Non-synergistic cytotoxic effects of Fusarium and Alternaria toxin combinations in Caco-2 cells. Toxicol. Lett. 2016, 241, 1–8. [Google Scholar] [CrossRef]
- Bensassi, F.; Gallerne, C.; Sharaf, E.D.O.; Hajlaoui, M.R.; Bacha, H.; Lemaire, C. Cell death induced by the Alternaria mycotoxin alternariol. Toxicol. In Vitro 2012, 26, 915–923. [Google Scholar] [CrossRef]
- Pfeiffer, E.; Eschbach, S.; Metzler, M. Alternaria toxins: DNA strand-breaking activity in mammalian cellsin vitro. Mycotoxin Res. 2007, 23, 152–157. [Google Scholar] [CrossRef]
- Commission Recommendation (EU) 2022/553 of 5 April 2022 on Monitoring the Presence of Alternaria Toxins in Food. 2022. Available online: https://op.europa.eu/en/publication-detail/-/publication/1bd43c13-b544-11ec-b6f4-01aa75ed71a1/language-en (accessed on 20 November 2022).
- Motta, S.D.; Soares, L.M.V. Survey of Brazilian tomato products for alternariol, alternariol monomethyl ether, tenuazonic acid and cyclopiazonic acid. Food Addit. Contam. 2001, 18, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, Y.; Curtui, V.; Dietrich, R.; Gross, M.; Latif, H.; Märtlbauer, E.; Usleber, E. Widespread occurrence of low levels of alternariol in apple and tomato products, as determined by comparative immunochemical assessment using monoclonal and polyclonal antibodies. J. Agric. Food Chem. 2011, 59, 6360–6368. [Google Scholar] [CrossRef]
- Terminiello, L.; Patriarca, A.; Pose, G.; Pinto, V.F. Occurrence of alternariol, alternariol monomethyl ether and tenuazonic acid in Argentinean tomato puree. Mycotoxin Res. 2006, 22, 236–240. [Google Scholar] [CrossRef]
- Noser, J.; Schneider, P.; Rother, M.; Schmutz, H. Determination of six Alternaria toxins with UPLC-MS/MS and their occurrence in tomatoes and tomato products from the Swiss market. Mycotoxin Res. 2011, 27, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Puntscher, H.; Cobankovic, L.; Marko, D.; Warth, B. Quantitation of free and modified Alternaria mycotoxins in European food products by LC-MS/MS. Food Control 2019, 102, 157–165. [Google Scholar] [CrossRef]
- Puntscher, H.; Kütt, M.L.; Marko, D.; Warth, B.; Skrinjar, P.; Mikula, H.; Frhlich, J.; Podlech, J. Tracking emerging mycotoxins in food: Development of an LC-MS/MS method for free and modified Alternaria toxins. Anal. Bioanal. Chem. 2018, 410, 4481–4494. [Google Scholar] [CrossRef]
- Kang, Z.; Buchenauer, H. Ultrastructural and cytochemical studies on the infection of wheat spikes by Fusarium culmorum as well as on degradation of cell wall components and localization of mycotoxins in the host tissue. Mycotoxin Res. 2000, 16, 1–5. [Google Scholar] [CrossRef]
- Wang, K.L.; Zheng, X.F.; Zhang, X.Y.; Zhao, L.N.; Yang, Q.Y.; Boateng, N.A.S.; Ahima, J.; Liu, J.; Zhang, H.Y. Comparative transcriptomic analysis of the interaction between Penicillium expansum and apple fruit (Malus pumila mill.) during early stages of infection. Microorganisms 2019, 7, 495. [Google Scholar] [CrossRef]
- Xie, H.L.; Wang, X.P.; Zhang, L.X.; Wang, T.; Zhang, W.; Jiang, J.; Chang, P.K.; Chen, Z.Y.; Bhatnagar, D.; Zhang, Q. Monitoring metabolite production of aflatoxin biosynthesis by orbitrap fusion mass spectrometry and a D-optimal mixture design method. Anal. Chem. 2018, 90, 14331–14338. [Google Scholar] [CrossRef]
- Rizwana, H.; Bokahri, N.A.; Alsahli, S.A.; Showiman, A.S.A.; Alzahrani, R.M.; Aldehaish, H.A. Postharvest disease management of Alternaria spots on tomato fruit by Annona muricata fruit extracts. Saudi J. Biol. Sci. 2021, 28, 2236–3344. [Google Scholar] [CrossRef]
- Miller, B.L.; Huffaker, R.C. Differential induction of endoproteinases during senescence of attached and detached barley leaves. Plant Physiol. 1985, 78, 442–446. [Google Scholar] [CrossRef]
- Stinson, E.E.; Osman, S.F.; Heisler, E.G.; Siciliano, J.; Bills, D.D. Mycotoxin production in whole tomatoes, apples, oranges, and lemons. J. Agric. Food Chem. 1981, 29, 790–792. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Gallone, T.; Garganese, F.; Caruso, A.G.; Amenduni, M.; Ippolito, A. Contamination of fresh and dried tomato by Alternaria toxins in southern Italy. Food Addit. Contam. 2019, 36, 789–799. [Google Scholar] [CrossRef]
- Patricia, L.; Dini, V.; Hans, M.; Martien, S.; Joyce, D.S.; Erika, P.; Monique, D.N. Alternaria toxins and conjugates in selected foods in the Netherlands. Food Control 2016, 69, 153–159. [Google Scholar]
- Walravens, J.; Mikula, H.; Rychlik, M.; Asam, S.; Devos, T.; Ediage, E.N.; Mavungu, J.D.D.; Jacxsens, L.; Landschoot, A.V.; Vanhaecke, L.; et al. Validated UPLC-MS/MS methods to quantitate free and conjugated Alternaria toxins in commercially available tomato products, fruit and vegetable juices in belgium. J. Agric. Food Chem. 2016, 64, 5101–5109. [Google Scholar] [CrossRef]
- Sebastian, T.S.; Beate, N.K.; Erika, P.; Rolf, G.; Manfred, M.; Mirko, B.; Sabine, E.K. Sulfoglucosides as novel modified forms of the mycotoxins alternariol and alternariol monomethyl ether. J. Agric. Food Chem. 2016, 64, 8892–8901. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, Q.; Fan, Y.; Jia, Q.; Duan, S.; Liu, F.; Jia, B.; Wang, G.; Guo, W.; Wang, C. The Potential of Alternaria Toxins Production by A. alternata in Processing Tomatoes. Toxins 2022, 14, 827. https://doi.org/10.3390/toxins14120827
Qin Q, Fan Y, Jia Q, Duan S, Liu F, Jia B, Wang G, Guo W, Wang C. The Potential of Alternaria Toxins Production by A. alternata in Processing Tomatoes. Toxins. 2022; 14(12):827. https://doi.org/10.3390/toxins14120827
Chicago/Turabian StyleQin, Qiaomei, Yingying Fan, Qinlan Jia, Shuaishuai Duan, Fengjuan Liu, Binxin Jia, Guangquan Wang, Wanhui Guo, and Cheng Wang. 2022. "The Potential of Alternaria Toxins Production by A. alternata in Processing Tomatoes" Toxins 14, no. 12: 827. https://doi.org/10.3390/toxins14120827
APA StyleQin, Q., Fan, Y., Jia, Q., Duan, S., Liu, F., Jia, B., Wang, G., Guo, W., & Wang, C. (2022). The Potential of Alternaria Toxins Production by A. alternata in Processing Tomatoes. Toxins, 14(12), 827. https://doi.org/10.3390/toxins14120827