The Middle Eastern Cousin: Comparative Venomics of Daboia palaestinae and Daboia russelii

 ,
,  ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
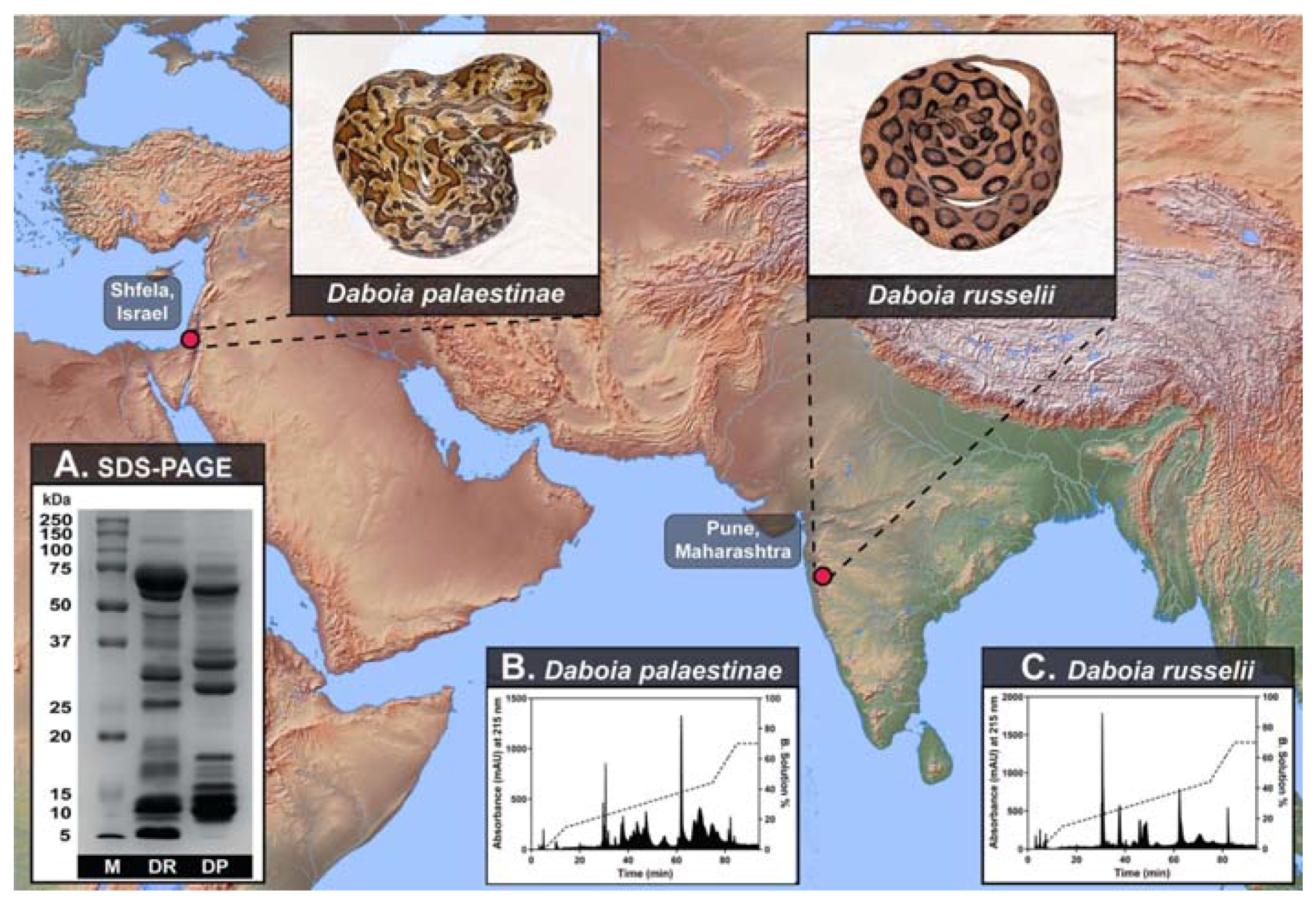
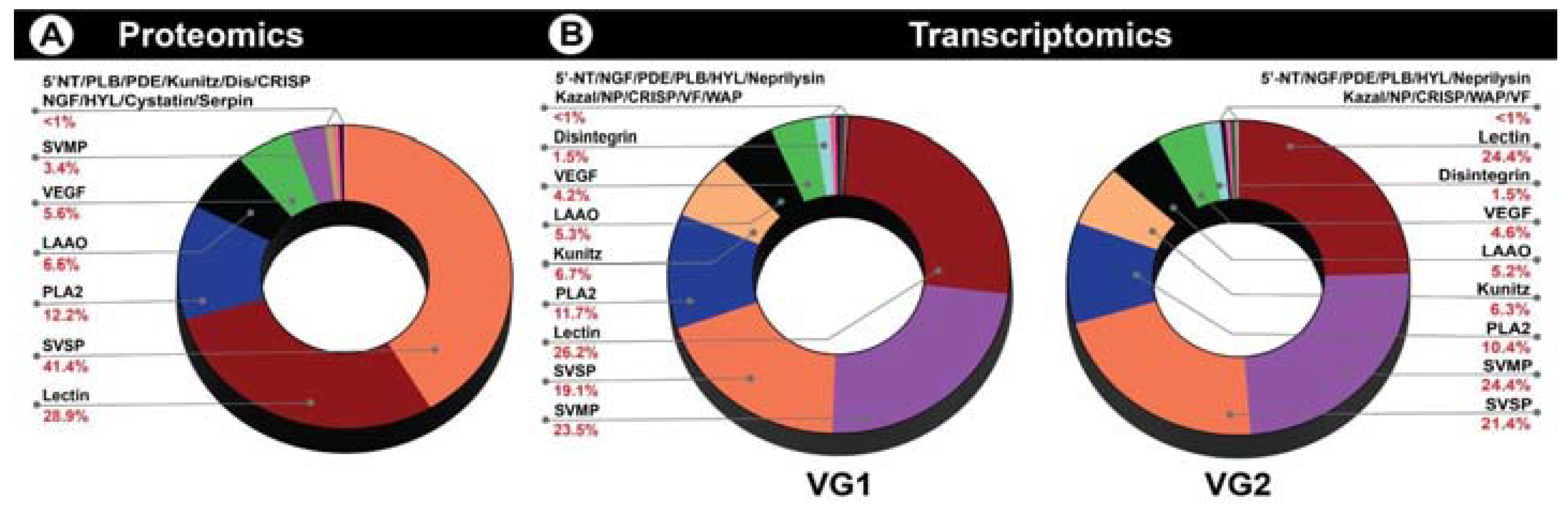
2.1. Venom Proteomics
2.2. Venom Gland Transcriptomics
2.3. Venom Biochemistry
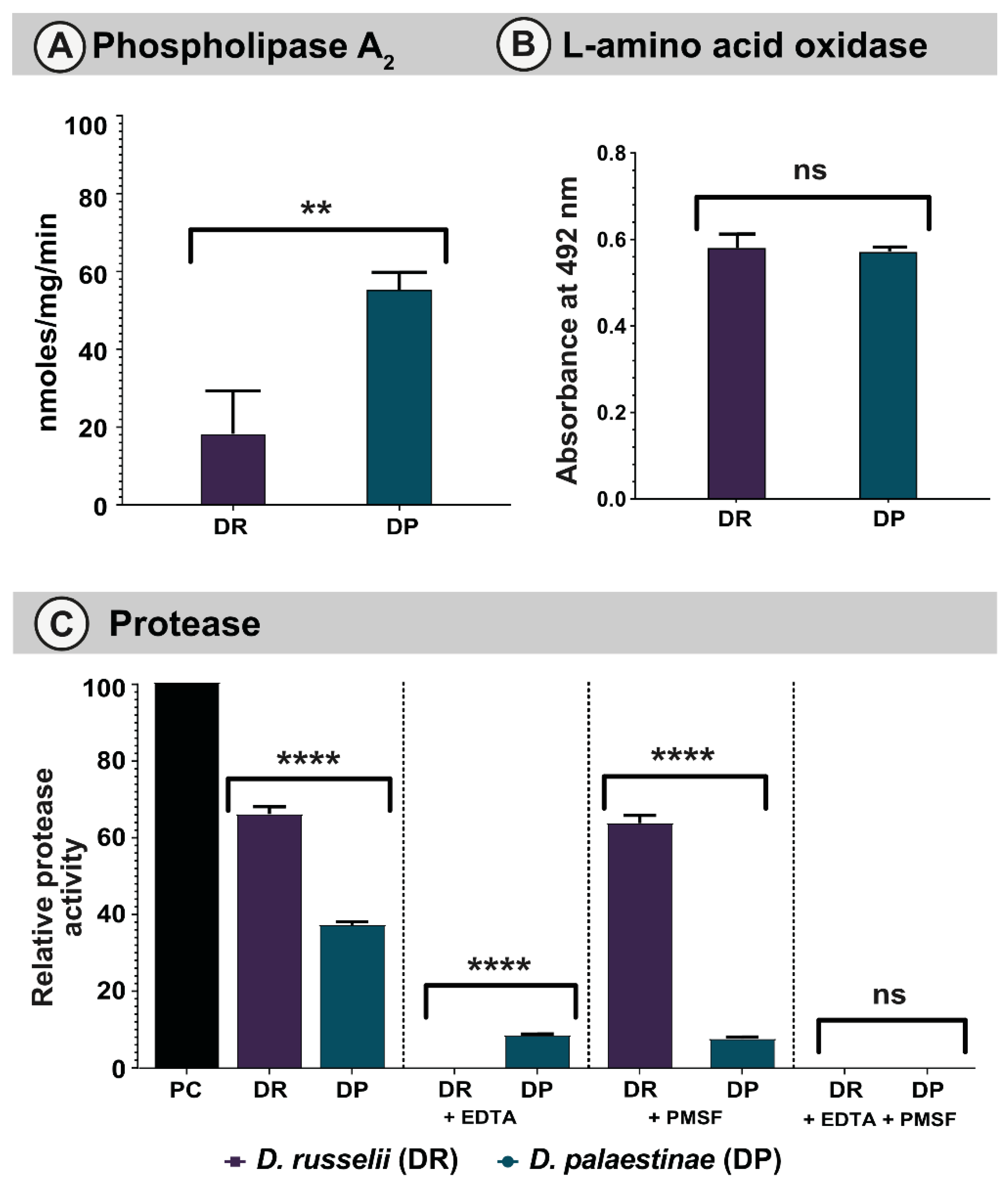
2.3.1. PLA2 assay
2.3.2. LAAO Assay
2.3.3. Snake Venom Protease Assay
2.3.4. DNase Assay
2.3.5. Fibrinogenolytic Assay
2.4. Preclinical Assessments
2.4.1. Venom Toxicity
2.4.2. Venom-Induced Morbidity
2.4.3. Venom Induced Nephrotoxicity
3. Discussion
3.1. Disparate Venoms of D. russelii and D. palaestinae
3.2. Venom Production in D. palaestinae Is Not Partitioned between the Venom Glands
3.3. The Role of Compositional and Activity Differences in Determining the Clinical Relevance of Daboia Species
4. Conclusions
5. Materials and Methods
5.1. Sampling Permits, and the Collection of Venom and Venom Gland
5.2. Ethical Statements
5.3. Proteomic Analyses
5.3.1. Protein Estimation, One-Dimensional Gel Electrophoresis and In-Gel Digestion
5.3.2. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
5.3.3. Reversed-Phase High-Performance Liquid Chromatography (RP-HPLC)
5.4. Transcriptomics Analyses
5.4.1. RNA Isolation and Sequencing
5.4.2. Quality Assessment, De Novo Assembly and Annotation
5.4.3. Quantification and Differential Expression Analyses
5.5. Biochemical Characterisation
5.5.1. Colourimetric Phospholipase A2 (PLA2) Assay
5.5.2. L-amino Acid Oxidase (LAAO) Assay
5.5.3. Snake Venom Protease Assay
5.5.4. DNase Assay
5.5.5. Fibrinogenolytic Assay
5.6. Preclinical Assessments
5.6.1. The Median lethal Dose (LD50)
5.6.2. The Minimum Haemorrhagic Dose (MHD)
5.6.3. The Minimum Necrotic Dose (MND)
5.7. Nephrotoxic Potentials of Daboia Venoms
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Warrell, D.A. Snake venoms in science and clinical medicine 1. Russell’s viper: Biology, venom and treatment of bites. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 732–740. [Google Scholar] [CrossRef]
- Chaisakul, J.; Alsolaiss, J.; Charoenpitakchai, M.; Wiwatwarayos, K.; Sookprasert, N.; Harrison, R.A.; Chaiyabutr, N.; Chanhome, L.; Tan, C.H.; Casewell, N.R. Evaluation of the geographical utility of Eastern Russell’s viper (Daboia siamensis) antivenom from Thailand and an assessment of its protective effects against venom-induced nephrotoxicity. PLoS Negl. Trop. Dis. 2019, 13, e0007338. [Google Scholar] [CrossRef] [PubMed]
- Faisal, T.; Tan, K.Y.; Sim, S.M.; Quraishi, N.; Tan, N.H.; Tan, C.H. Proteomics, functional characterization and antivenom neutralization of the venom of Pakistani Russell’s viper (Daboia russelii) from the wild. J. Proteom. 2018, 183, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Senji Laxme, R.R.; Khochare, S.; De Souza, H.F.; Ahuja, B.; Suranse, V.; Martin, G.; Whitaker, R.; Sunagar, K. Beyond the ‘big four’: Venom profiling of the medically important yet neglected Indian snakes reveals disturbing antivenom deficiencies. PLoS Negl. Trop. Dis. 2019, 13, e0007899. [Google Scholar] [CrossRef]
- Mukherjee, A.K. The pro-coagulant fibrinogenolytic serine protease isoenzymes purified from Daboia russelii russelii venom coagulate the blood through factor V activation: Role of glycosylation on enzymatic activity. PLoS ONE 2014, 9, e86823. [Google Scholar] [CrossRef]
- Pla, D.; Sanz, L.; Quesada-Bernat, S.; Villalta, M.; Baal, J.; Chowdhury, M.A.W.; León, G.; Gutiérrez, J.M.; Kuch, U.; Calvete, J.J. Phylovenomics of Daboia russelii across the Indian subcontinent. Bioactivities and comparative in vivo neutralization and in vitro third-generation antivenomics of antivenoms against venoms from India, Bangladesh and Sri Lanka. J. Proteom. 2019, 207, 103443. [Google Scholar] [CrossRef] [PubMed]
- Abu Baker, M.A.; Al-Saraireh, M.; Amr, Z.; Amr, S.S.; Warrell, D.A. Snakebites in Jordan: A clinical and epidemiological study. Toxicon 2022, 208, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Agajany, N.; Kozer, E.; Agajany, N.; Trotzky, D.; Kishk, I.A.; Youngster, I. Is severity of Daboia (Vipera) palaestinae snakebites influenced by season of exposure? Toxicon 2021, 206, 51–54. [Google Scholar] [CrossRef]
- El Zahran, T.; Kazzi, Z.; Chehadeh, A.A.-H.; Sadek, R.; El Sayed, M.J. Snakebites in Lebanon: A descriptive study of snakebite victims treated at a tertiary care center in Beirut, Lebanon. J. Emergencies Trauma Shock. 2018, 11, 119. [Google Scholar]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Yee, K.T.; Pitts, M.; Tongyoo, P.; Rojnuckarin, P.; Wilkinson, M.C. Snake venom metalloproteinases and their peptide inhibitors from Myanmar Russell’s viper venom. Toxins 2017, 9, 15. [Google Scholar] [CrossRef]
- Higuchi, S.; Murayama, N.; Saguchi, K.; Ohi, H.; Fujita, Y.; Camargo, A.C.; Ogawa, T.; Deshimaru, M.; Ohno, M. Bradykinin-potentiating peptides and C-type natriuretic peptides from snake venom. Immunopharmacology 1999, 44, 129–135. [Google Scholar] [CrossRef]
- Leonardi, A.; Sajevic, T.; Pungerčar, J.; Križaj, I. Comprehensive study of the proteome and transcriptome of the venom of the most venomous european viper: Discovery of a new subclass of ancestral snake venom metalloproteinase precursor-derived proteins. J. Proteome Res. 2019, 18, 2287–2309. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.R.; Burin, S.M.; Menaldo, D.L.; de Castro, F.A.; Sampaio, S.V. Snake venom L-amino acid oxidases: An overview on their antitumor effects. J. Venom. Anim. Toxins Incl. Trop. Dis. 2014, 20, 23. [Google Scholar] [CrossRef] [PubMed]
- Paloschi, M.V.; Pontes, A.S.; Soares, A.M.; Zuliani, J.P. An update on potential molecular mechanisms underlying the actions of snake venom L-amino acid oxidases (LAAOs). Current medicinal chemistry 2018, 25, 2520–2530. [Google Scholar] [CrossRef]
- Ribeiro, P.H.; Zuliani, J.P.; Fernandes, C.F.; Calderon, L.A.; Stábeli, R.G.; Nomizo, A.; Soares, A.M. Mechanism of the cytotoxic effect of l-amino acid oxidase isolated from Bothrops alternatus snake venom. Int. J. Biol. Macromol. 2016, 92, 329–337. [Google Scholar] [CrossRef]
- Slagboom, J.; Kool, J.; Harrison, R.A.; Casewell, N.R. Haemotoxic snake venoms: Their functional activity, impact on snakebite victims and pharmaceutical promise. Br. J. Haematol. 2017, 177, 947–959. [Google Scholar] [CrossRef]
- Asega, A.F.; Menezes, M.C.; Trevisan-Silva, D.; Cajado-Carvalho, D.; Bertholim, L.; Oliveira, A.K.; Zelanis, A.; Serrano, S.M.T. Cleavage of proteoglycans, plasma proteins and the platelet-derived growth factor receptor in the hemorrhagic process induced by snake venom metalloproteinases. Sci. Rep. 2020, 10, 12912. [Google Scholar] [CrossRef]
- Lu, Q.; Clemetson, J.; Clemetson, K.J. Snake venoms and hemostasis. J. Thromb. Haemost. 2005, 3, 1791–1799. [Google Scholar] [CrossRef]
- Katkar, G.D.; Sundaram, M.S.; NaveenKumar, S.K.; Swethakumar, B.; Sharma, R.D.; Paul, M.; Vishalakshi, G.J.; Devaraja, S.; Girish, K.S.; Kemparaju, K. NETosis and lack of DNase activity are key factors in Echis carinatus venom-induced tissue destruction. Nat. Commun. 2016, 7, 11361. [Google Scholar] [CrossRef] [PubMed]
- Senji Laxme, R.; Khochare, S.; Attarde, S.; Suranse, V.; Iyer, A.; Casewell, N.R.; Whitaker, R.; Martin, G.; Sunagar, K. Biogeographic venom variation in Russell’s viper (Daboia russelii) and the preclinical inefficacy of antivenom therapy in snakebite hotspots. PLoS Negl. Trop. Dis. 2021, 15, e0009247. [Google Scholar] [CrossRef] [PubMed]
- Kattula, S.; Byrnes, J.R.; Wolberg, A.S. Fibrinogen and fibrin in hemostasis and thrombosis. Arterioscler. Thromb. Vasc. Biol. 2017, 37, e13–e21. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Morita, T. Snake venom components affecting blood coagulation and the vascular system: Structural similarities and marked diversity. Curr. Pharm. Des. 2007, 13, 2872–2886. [Google Scholar] [CrossRef] [PubMed]
- Prasad, N.B.; Uma, B.; Bhatt, S.K.; Gowda, V.T. Comparative characterisation of Russell’s viper (Daboia/Vipera russelli) venoms from different regions of the Indian peninsula. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1999, 1428, 121–136. [Google Scholar] [CrossRef]
- Oukkache, N.; Lalaoui, M.; Ghalim, N. General characterization of venom from the Moroccan snakes Macrovipera mauritanica and Cerastes cerastes. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 411–420. [Google Scholar] [CrossRef]
- Alirol, E.; Sharma, S.K.; Bawaskar, H.S.; Kuch, U.; Chappuis, F. Snake bite in South Asia: A review. PLoS neglected tropical diseases 2010, 4, e603. [Google Scholar] [CrossRef]
- Kularatne, S. Epidemiology and clinical picture of the Russell’s viper (Daboia russelii russelii) bite in Anuradhapura, Sri Lanka: A prospective study of 336 patients. Southeast Asian J. Trop. Med. Public Health 2003, 34, 855–862. [Google Scholar]
- Vikrant, S.; Jaryal, A.; Parashar, A. Clinicopathological spectrum of snake bite-induced acute kidney injury from India. World J. Nephrol. 2017, 6, 150. [Google Scholar] [CrossRef]
- Ariga, K.; Dutta, T.K.; Haridasan, S.; Puthenpurackal, P.S.P.; Harichandrakumar, K.; Parameswaran, S. Chronic Kidney Disease after Snake Envenomation Induced Acute Kidney Injury. Saudi J. Kidney Dis. Transplant. 2021, 32, 146. [Google Scholar]
- Tirosh-Levy, S.; Solomovich-Manor, R.; Comte, J.; Nissan, I.; Sutton, G.A.; Gabay, A.; Gazit, E.; Steinman, A. Daboia (Vipera) palaestinae Envenomation in 123 Horses: Treatment and Efficacy of Antivenom Administration. Toxins 2019, 11, 168. [Google Scholar] [CrossRef] [PubMed]
- Damm, M.; Hempel, B.-F.; Süssmuth, R.D. Old World Vipers—A Review about Snake Venom Proteomics of Viperinae and Their Variations. Toxins 2021, 13, 427. [Google Scholar] [CrossRef]
- Makran, B.; Fahmi, L.; Pla, D.; Sanz, L.; Oukkache, N.; Lkhider, M.; Ghalim, N.; Calvete, J.J. Snake venomics of Macrovipera mauritanica from Morocco, and assessment of the para-specific immunoreactivity of an experimental monospecific and a commercial antivenoms. J. Proteom. 2012, 75, 2431–2441. [Google Scholar] [CrossRef]
- Risch, M.; Georgieva, D.; von Bergen, M.; Jehmlich, N.; Genov, N.; Arni, R.K.; Betzel, C. Snake venomics of the Siamese Russell’s viper (Daboia russelli siamensis)—Relation to pharmacological activities. J. Proteom. 2009, 72, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Momic, T.; Arlinghaus, F.T.; Arien-Zakay, H.; Katzhendler, J.; Eble, J.A.; Marcinkiewicz, C.; Lazarovici, P. Pharmacological aspects of Vipera xantina palestinae venom. Toxins 2011, 3, 1420–1432. [Google Scholar] [CrossRef]
- Durban, J.; Pérez, A.; Sanz, L.; Gómez, A.; Bonilla, F.; Rodríguez, S.; Chacón, D.; Sasa, M.; Angulo, Y.; Gutiérrez, J.M.; et al. Integrated “omics” profiling indicates that miRNAs are modulators of the ontogenetic venom composition shift in the Central American rattlesnake, Crotalus simus simus. BMC Genom. 2013, 14, 234. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Kalita, B.; Mackessy, S.P. A proteomic analysis of Pakistan Daboia russelii russelii venom and assessment of potency of Indian polyvalent and monovalent antivenom. J. Proteom. 2016, 144, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Khochare, S.; Senji Laxme, R.; Attarde, S.; Dam, P.; Suranse, V.; Khaire, A.; Martin, G.; Captain, A. A wolf in another wolf’s clothing: Post-genomic regulation dictates venom profiles of medically-important cryptic kraits in India. Toxins 2021, 13, 69. [Google Scholar] [CrossRef]
- Kalita, B.; Mackessy, S.P.; Mukherjee, A.K. Proteomic analysis reveals geographic variation in venom composition of Russell’s Viper in the Indian subcontinent: Implications for clinical manifestations post-envenomation and antivenom treatment. Expert Rev. Proteom. 2018, 15, 837–849. [Google Scholar] [CrossRef] [PubMed]
- Kalita, B.; Patra, A.; Das, A.; Mukherjee, A.K. Proteomic analysis and immuno-profiling of eastern India Russell’s Viper (Daboia russelii) venom: Correlation between RVV composition and clinical manifestations post RV bite. J. Proteome Res. 2018, 17, 2819–2833. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Martínez, D.; Plenge-Tellechea, L.F.; Gatica-Colima, A.; Cruz-Pérez, M.S.; Aguilar-Yáñez, J.M.; Licona-Cassani, C. Functional mining of the Crotalus Spp. venom protease repertoire reveals potential for chronic wound therapeutics. Molecules 2020, 25, 3401. [Google Scholar] [CrossRef]
- Yamashita, K.M.; Alves, A.F.; Barbaro, K.C.; Santoro, M.L. Bothrops jararaca venom metalloproteinases are essential for coagulopathy and increase plasma tissue factor levels during envenomation. PLoS Negl. Trop. Dis. 2014, 8, e2814. [Google Scholar] [CrossRef]
- Sharma, M.; Gogoi, N.; Dhananjaya, B.; Menon, J.C.; Doley, R. Geographical variation of Indian Russell’s viper venom and neutralization of its coagulopathy by polyvalent antivenom. Toxin Rev. 2014, 33, 7–15. [Google Scholar] [CrossRef]
- Sunagar, K.; Columbus-Shenkar, Y.Y.; Fridrich, A.; Gutkovich, N.; Aharoni, R.; Moran, Y. Cell type-specific expression profiling unravels the development and evolution of stinging cells in sea anemone. BMC Biol. 2018, 16, 108. [Google Scholar] [CrossRef]
- Columbus-Shenkar, Y.Y.; Sachkova, M.Y.; Macrander, J.; Fridrich, A.; Modepalli, V.; Reitzel, A.M.; Sunagar, K.; Moran, Y. Dynamics of venom composition across a complex life cycle. eLife 2018, 7, e35014. [Google Scholar] [CrossRef]
- Dutertre, S.; Jin, A.-H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation-and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.A.; Mayhew, M.L.; Jin, J.; Herzig, V.; Undheim, E.A.; Sombke, A.; Fry, B.G.; Meritt, D.J.; King, G.F. The assassin bug Pristhesancus plagipennis produces two distinct venoms in separate gland lumens. Nat. Commun. 2018, 9, 755. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Bonilla, F.; Granados-Martínez, S.; Sanz, L.; Lomonte, B.; Sasa, M. Venomics of the Duvernoy’s gland secretion of the false coral snake Rhinobothryum bovallii (Andersson, 1916) and assessment of venom lethality towards synapsid and diapsid animal models. J. Proteom. 2020, 225, 103882. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Zdenek, C.N.; Nouwens, A.; Sweeney, C.; Dunstan, N.; Fry, B.G. A symmetry or asymmetry: Functional and compositional comparison of venom from the left and right glands of the Indochinese spitting cobra (Naja siamensis). Toxicon: X 2020, 7, 100050. [Google Scholar] [CrossRef]
- Suraweera, W.; Warrell, D.; Whitaker, R.; Menon, G.; Rodrigues, R.; Fu, S.H.; Begum, R.; Sati, P.; Piyasena, K. Trends in snakebite deaths in India from 2000 to 2019 in a nationally representative mortality study. eLife 2020, 9, e54076. [Google Scholar] [CrossRef]
- Abd Rabou, A. On the occurrence and health risks of the venomous Palestine Viper (Vipera palaestinae Werner, 1938) in the Gaza Strip-Palestine. Biomed. J. Sci. Tech. Res. Mini Rev. 2019, 18, 13934–13937. [Google Scholar] [CrossRef]
- Kochva, E. A quantitative study of venom secretion by Vipera palaestinae. Am. J. Trop. Med. Hyg. 1960, 9, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Bentur, Y.; Cahana, A. Unusual local complications of Vipera palaestinae bite. Toxicon 2003, 41, 633–635. [Google Scholar] [CrossRef]
- Pivko-Levy, D.; Munchnak, I.; Rimon, A.; Balla, U.; Scolnik, D.; Hoyte, C.; Voliovitch, Y.; Glatstein, M. Evaluation of antivenom therapy for Vipera palaestinae bites in children: Experience of two large, tertiary care pediatric hospitals. Clin. Toxicol. 2017, 55, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, R.B.; Gawarammana, I.B.; De Silva, D.; Dangolla, A.; Mallawa, C.; Premarathna, A.; Silva, I.D. Clinico-epidemiology and management of Russell’s viper (Daboia russelii) envenoming in dogs in Sri Lanka. Toxicol. Rep. 2019, 6, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Warrell, D.A. Clinical toxicology of snakebite in Asia. In Handbook of: Clinical Toxicology of Animal Venoms and Poisons; CRC Press: Boca Raton, FL, USA, 2017; pp. 493–594. [Google Scholar]
- Warrell, D.A.; Gutiérrez, J.M.; Calvete, J.J.; Williams, D. New approaches & technologies of venomics to meet the challenge of human envenoming by snakebites in India. Indian J. Med. Res. 2013, 138, 38–59. [Google Scholar]
- Harrison, R.A.; Hargreaves, A.; Wagstaff, S.C.; Faragher, B.; Lalloo, D.G. Snake envenoming: A disease of poverty. PLoS Negl. Trop. Dis. 2009, 3, e569. [Google Scholar] [CrossRef]
- United Nations, Department of Economics and Social Affairs, Population Division. World Urbanization Prospects: The 2018 Revision (ST/ESA/SER/420); United Nations: New York, NY, USA, 2018. [Google Scholar]
- Leary, S.L.; Underwood, W.; Anthony, R.; Cartner, S.; Corey, D.; Grandin, T.; Greenacre, C.; Gwaltney-Brant, S.; McCrackin, M.A.; Meyer, R.; et al. AVMA Guidelines for the Euthanasia of Animals: 2013 Edition; American Veterinary Medical Association: Schaumburg, IL, USA, 2013. [Google Scholar]
- WHO. World Health Organisation Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; World Health Organisation: Geneva, Switzerland, 2018. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Smith, B. SDS Polyacrylamide Gel Electrophoresis of Proteins; Springer: Berlin/Heidelberg, Germany, 1984; pp. 41–55. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Tan, N.H.; Wong, K.Y.; Tan, C.H. Venomics of Naja sputatrix, the Javan spitting cobra: A short neurotoxin-driven venom needing improved antivenom neutralization. J. Proteom. 2017, 157, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. Babraham Bioinformatics-FastQC a Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc.2010 (accessed on 22 October 2022).
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nature biotechnology 2011, 29, 644. [Google Scholar] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC bioinformatics 2011, 12, 323. [Google Scholar] [CrossRef]
- Freitas-de-Sousa, L.A.; Nachtigall, P.G.; Portes-Junior, J.A.; Holding, M.L.; Nystrom, G.S.; Ellsworth, S.A.; Guimarães, N.C.; Tioyama, E.; Ortiz, F.; Silva, B.R.; et al. Size matters: An evaluation of the molecular basis of ontogenetic modifications in the composition of Bothrops jararacussu snake venom. Toxins 2020, 12, 791. [Google Scholar] [CrossRef]
- Tasoulis, T.; Lee, M.S.; Ziajko, M.; Dunstan, N.; Sumner, J.; Isbister, G.K. Activity of two key toxin groups in Australian elapid venoms show a strong correlation to phylogeny but not to diet. BMC Evol. Biol. 2020, 20, 9. [Google Scholar] [CrossRef]
- Chowdhury, M.; Miyoshi, S.; Shinoda, S. Purification and characterization of a protease produced by Vibrio mimicus. Infect. Immun. 1990, 58, 4159–4162. [Google Scholar]
- Gerceker, D.; Karasartova, D.; Elyürek, E.; Barkar, S.; KIyan, M.; Özsan, T.M.; Calgin, M.K.; Sahin, F. A new, simple, rapid test for detection of DNase activity of microorganisms: DNase Tube test. J. Gen. Appl. Microbiol. 2009, 55, 291–294. [Google Scholar] [CrossRef]
- Teng, C.-M.; Ouyang, C.; Lin, S.-C. Species difference in the fibrinogenolytic effects of α-and β-fibrinogenases from Trimeresurus mucrosquamatus snake venom. Toxicon 1985, 23, 777–782. [Google Scholar] [CrossRef]
- op den Brouw, B.; Ghezellou, P.; Casewell, N.R.; Ali, S.A.; Fathinia, B.; Fry, B.G.; Bos, M.H.A.; Ikonomopoulou, M.P. Pharmacological characterisation of Pseudocerastes and Eristicophis viper venoms reveal anticancer (Melanoma) properties and a potentially novel mode of fibrinogenolysis. Int. J. Mol. Sci. 2021, 22, 6896. [Google Scholar] [CrossRef] [PubMed]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: London, UK, 1971. [Google Scholar]
- Gutiérrez, J.M.; Solano, G.; Pla, D.; Herrera, M.; Segura, Á.; Vargas, M.; Villalta, M.; Sánchez, A.; Sanz, L.; Lomonte, B.; et al. Preclinical evaluation of the efficacy of antivenoms for snakebite envenoming: State-of-the-art and challenges ahead. Toxins 2017, 9, 163. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Kondo, S.; Ikezawa, H.; Murata, R.; Ohsaka, A. Studies on the quantitative method for determination of hemorrhagic activity of Habu snake venom. Jpn. J. Med. Sci. Biol. 1960, 13, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Hedrich, H. The Laboratory Mouse; Academic Press: Cambridge, MA, UK, 2004. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senji Laxme, R.R.; Khochare, S.; Attarde, S.; Kaur, N.; Jaikumar, P.; Shaikh, N.Y.; Aharoni, R.; Primor, N.; Hawlena, D.; Moran, Y.; et al. The Middle Eastern Cousin: Comparative Venomics of Daboia palaestinae and Daboia russelii. Toxins 2022, 14, 725. https://doi.org/10.3390/toxins14110725
Senji Laxme RR, Khochare S, Attarde S, Kaur N, Jaikumar P, Shaikh NY, Aharoni R, Primor N, Hawlena D, Moran Y, et al. The Middle Eastern Cousin: Comparative Venomics of Daboia palaestinae and Daboia russelii. Toxins. 2022; 14(11):725. https://doi.org/10.3390/toxins14110725
Chicago/Turabian StyleSenji Laxme, R. R., Suyog Khochare, Saurabh Attarde, Navneet Kaur, Priyanka Jaikumar, Naeem Yusuf Shaikh, Reuven Aharoni, Naftali Primor, Dror Hawlena, Yehu Moran, and et al. 2022. "The Middle Eastern Cousin: Comparative Venomics of Daboia palaestinae and Daboia russelii" Toxins 14, no. 11: 725. https://doi.org/10.3390/toxins14110725
APA StyleSenji Laxme, R. R., Khochare, S., Attarde, S., Kaur, N., Jaikumar, P., Shaikh, N. Y., Aharoni, R., Primor, N., Hawlena, D., Moran, Y., & Sunagar, K. (2022). The Middle Eastern Cousin: Comparative Venomics of Daboia palaestinae and Daboia russelii. Toxins, 14(11), 725. https://doi.org/10.3390/toxins14110725