The Impact of Cyanobacteria Blooms on the Aquatic Environment and Human Health

 ,
,
Abstract
1. Introduction
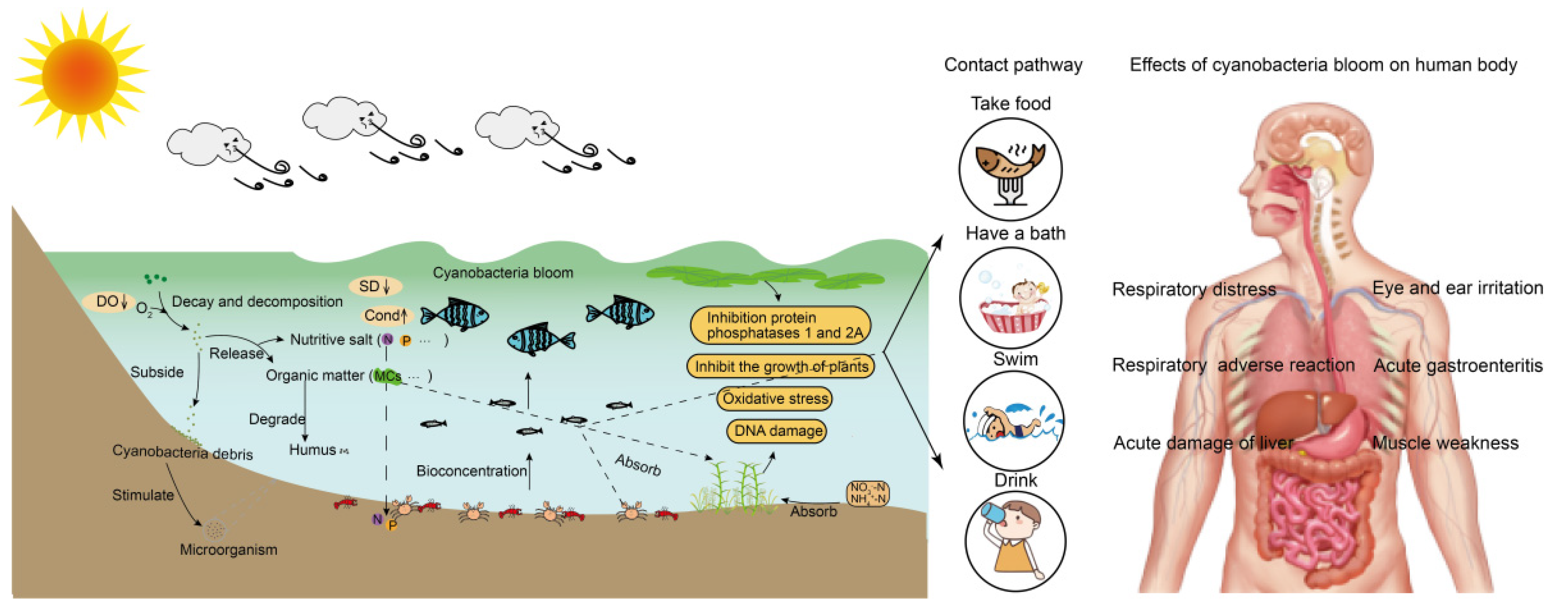
2. The Pollution of Cyanobacteria Blooms to Water
2.1. Impacts of Cyanobacteria Blooms on Aquatic Fauna
2.2. Impacts of Cyanobacteria Blooms on Aquatic Flora
2.3. Impact of Cyanobacteria Blooms on Microorganisms in the Aquatic Environment
3. Impacts of Cyanobacteria Blooms on Human Health
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohr, K.I.; Brinkmann, N.; Friedl, T. Cyanobacteria. In Encyclopedia of Geobiology; Springer: Dordrecht, The Netherlands, 2011; pp. 306–311. [Google Scholar]
- Stewart, I.; Carmichael, W.W.; Backer, L.C. Toxic Cyanobacteria. Water Sanit.-Relat. Dis. Environ. 2011, 8, 95–109. [Google Scholar] [CrossRef]
- Paerl, H.W. Nuisance phytoplankton blooms in coastal, estuarine, and inland waters. Limnol. Oceanogr. 1988, 33, 823–843. [Google Scholar] [CrossRef]
- Qian, K.; Chen, Y.; Song, X. Long-term development of phytoplankton dominant species related to eutrophicarion in Lake Taihu. Ecol. Sci. 2008, 27, 65–70. [Google Scholar]
- Bai, M.; Zhou, S.; Zhao, M. Cyanobacterial bloom control in Taihu basin: Analysis of cost-risk analysis framework based on cooperative game. J. Clean. Prod. 2018, 195, 318–327. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, M.; Liu, L.; Liu, X.; Chen, H.; Yang, J. The antibiotic resistome of free-living and particle-attached bacteria under a reservoir cyanobacterial bloom. Environ. Int. 2018, 117, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Aubriot, L.; Bonilla, S. Regulation of phosphate uptake reveals cyanobacterial bloom resilience to shifting N: P ratios. Freshwater Biol. 2018, 63, 318–329. [Google Scholar] [CrossRef]
- Huang, I.S.; Zimba, P.V. Cyanobacterial bioactive metabolites—A review of their chemistry and biology. Harmful Algae 2019, 86, 101608. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Lin, G.; Gao, G.; Qin, B.; Shen, J. Bacterial and archaeal assemblages in sediments of a large shallow freshwater lake, lake taihu, as revealed by denaturing gradient gel electrophoresis. J. Appl. Microbiol. 2010, 106, 1022–1032. [Google Scholar] [CrossRef]
- Gallitelli, M.; Ungaro, N.; Addante, L.M.; Procacci, V.; Silveri, N.G.; Sabbà, C. Respiratory Illness as a Reaction to Tropical Algal Blooms Occurring in a Temperate Climate. Jama 2005, 293, 2599–2600. [Google Scholar]
- Zhao, X.; Zhu, G.; Xu, L.; Lv, X. Characteristics of natural decomposition of cyanobacteri. Jiangsu J. Agric. Sci. 2013, 29, 312–318. [Google Scholar]
- Kong, F.; Ma, R.; Gao, J.; Wu, X. The theory and practice of prevention, forecast and warning on cyanobacteria bloom in Lake Taihu. J. Lake Sci. 2009, 21, 314–328. [Google Scholar]
- Liu, G.; Zhong, J.; He, J.; Zhang, L.; Fan, C. Effects of Black Spots of Dead-Cyanobacterial Mats on Fe-S-P Cycling in Sediments of Zhushan Bay, Lake Taihu. J. Environ. Sci. 2009, 30, 2520–2526. [Google Scholar]
- Shang, L.; Ke, F.; Li, W.; Xu, X.; Song, Y.; Feng, M. Laboratory research on the contaminants release during the anaerobic decomposition of high-density cyanobacteria. J. Lake Sci. 2013, 25, 47–54. [Google Scholar]
- Li, K.; Guan, B.; Liu, Z. Experiments on decomposition rate and release forms of nitrogen and phosphorus from the decomposing cyanobacterial detritus. J. Lake Sci. 2011, 23, 919–925. [Google Scholar]
- Krivtsov, V.; Bellinger, E.G.; Sigee, D.C. Elemental composition of Microcystis aeruginosa under conditions of lake nutrient depletion. Aquat. Ecol. 2005, 39, 123–134. [Google Scholar] [CrossRef]
- Li, X.; Li, Z.; Wang, X.; Zhang, S.; Wang, H.; Li, R.; Wang, G.; Li, Q. Characteristics of Dissolved Organic Matter in Overlying Water During Algal Bloom Decay. Environ. Sci. 2021, 42, 3281–3290. [Google Scholar]
- Zhang, S.; Wang, W.; Zhang, K.; Xu, P.; Lu, Y. Phosphorus release from cyanobacterial blooms during their decline period in eutrophic Dianchi Lake, China. Environ. Sci. Pollut. Res. 2018, 25, 13579–13588. [Google Scholar] [CrossRef]
- Zhu, G.; Qin, B.; Zhang, Y.; Li, Y.; Zhu, M.; Xu, H.; Zhang, B. Fluctuation of phosphorus concentration in Lake Taihu in the past 70 years and future control strategy. J. Lake Sci. 2021, 33, 957–973. [Google Scholar]
- Wang, J.; Li, J.; Jiang, D.; Wu, H.; Wang, S.; Lin, A.J. The potential impact of phosphorus concentration in typical lakes in China on water body indicators and cyanobacteria bloom trends based on meta-analysis. J. Beijing Univ. Chem. Technol. 2021, 48, 59–67. [Google Scholar]
- Li, K. The fate of nutrients released during the decomposing of cyanobacterial detritus in submerged macrophytes zones. Huazhong Agric. Univ. 2011.
- Li, D. Ecological threshold for prevention and control of cyanobacterial blooms in Baiyangdian Lake. Chin. Res. Acad. Environ. Sci. 2021. [Google Scholar] [CrossRef]
- Xia, H.; Song, T.; Wang, L.; Jiang, L.; Zhou, Q.; Wang, W.; Liu, L.; Yang, P.; Zhang, X. Effects of dietary toxic cyanobacteria and ammonia exposure on immune function of blunt snout bream (Megalabrama amblycephala). Fish Shellfish Immunol. 2018, 78, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Ferrão-Filho, A.D.S.; Kozlowsky-Suzuki, B. Cyanotoxins: Bioaccumulation and effects on aquatic animals. Mar. Drugs 2011, 9, 2729–2772. [Google Scholar] [CrossRef]
- Yang, J.; Hu, L.; Zhou, W.; Chen, J.; Shi, Z. Bioaccumulation of microcystin and antioxidative response in Carassius auratus L. Ecol. Environ. Sci. 2009, 18, 2044–2050. [Google Scholar]
- Zhang, J.; Wang, Z.; Song, Z.; Xie, Z.; Li, L.; Song, L. Bioaccumulation of microcystins in two freshwater gastropods from a cyanobacteria-bloom plateau lake, Lake Dianchi. Environ. Pollut. 2012, 164, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.J.; Dietrich, D.R. Pathological and biochemical characterization of microcystin-induced hepatopancreas and kidney damage in carp (Cyprinus carpio). Toxicol. Appl. Pharmacol. 2000, 164, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Wang, L.; Yang, H.; Wei, W.; Zhang, Y. Low-dose microcystins MC-LR induced hepatopancreas injury and apoptosis in Macrobrachium rosenbergii. Asian J. Ecotox. 2020, 2, 171–179. [Google Scholar]
- Andersen, R.J.; Luu, H.A.; Chen, D.Z.X.; Holmes, C.F.B.; Kent, M.L.; Blanc, M.L.; Taylor, F.J.R.; Williams, D.E. Chemical and biological evidence links microcystins to salmon ‘netpen liver disease’. Toxicon 1993, 31, 1315. [Google Scholar] [CrossRef]
- Xie, L.; Yokoyama, A.; Nakamura, K.; Park, H. Accumulation of microcystins in various organs of the freshwater snail sinotaia histrica and three fishes in a temperate lake, the eutrophic lake suwa, japan. Toxicon 2007, 49, 646–652. [Google Scholar] [CrossRef]
- Yang, W.; Xiang, F.; Liang, L.; Zhou, Y. Toxicity of Ammonia and Its Effects on Oxidative Stress Mechanisms of Juvenile Crucian Carp (Carassius auratus). J. Freshw. Ecol. 2010, 25, 297–302. [Google Scholar] [CrossRef]
- Benli, A.Ç.K.; Özkul, A. Acute toxicity and histopathological effects of sublethal fenitrothion on Nile tilapia, Oreochromis niloticus. Pestic. Biochem. Phys. 2010, 97, 32–35. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, S.; Ge, X.; Xia, S.; Zhu, J.; Miao, L.; Lin, Y.; Liang, H.; Pen, W.; Su, Y.; et al. Acute effects of ammonia exposure on the plasma and haematological parameters and histological structure of the juvenile blunt snout bream, Megalobrama amblycephala, and post-exposure recovery. Aquac. Res. 2017, 49, 1008–1019. [Google Scholar] [CrossRef]
- Li, D.; Li, G.; Chen, W.; Liu, Y. Interactions between a cyanobacterial bloom (Microcystis) and the submerged aquatic plant Ceratophyllum oryzetorum. Kom. Chin. J. Oceanol. Limn. 2009, 27, 38–42. [Google Scholar] [CrossRef]
- Shang, Y.; Guan, B.; Zheng, J.; Kang, Y. Effect of Cyanobacteria Accumulation on Water Environment and Submerged Plant Growth. Chin. Agric. Sci. Bull. 2015, 31, 195–198. [Google Scholar]
- Chen, K.; Li, W.; Wu, Q.; Qiang, S. Impacts of Cycanobacteria on the Growth of Submerged Macrophytes, Dianchi Lake. J. Lake Sci. 2003, 15, 364–368. [Google Scholar]
- Jiang, M.; Zhou, Y.; Wang, N.; Xu, L.; Zheng, Z.; Zhang, J. Allelopathic effects of harmful algal extracts and exudates on biofilms on leaves of Vallisneria natans. Sci. Total. Environ. 2018, 655, 823–830. [Google Scholar] [CrossRef]
- Saqrane, S.; Oudra, B. Cyanobacterial toxins: A short review on phytotoxic effect in an aquatic environment. Afr. J. Environ. Sci. Technol. 2011, 5, 1146–1151. [Google Scholar] [CrossRef]
- Ha, M.H.; Contardo-Jara, V.; Pflugmacher, S. Uptake of the cyanobacterial neurotoxin, anatoxin-a, and alterations in oxidative stress in the submerged aquatic plant Ceratophyllum demersum. Ecotox. Environ. Safe 2014, 101, 205–212. [Google Scholar] [CrossRef]
- Romanowska-Duda, Z.; Tarczyńska, M. The influence of microcystin-LR and hepatotoxic cyanobacterial extract on the water plant Spirodela oligorrhiza. Environ. Toxicol. 2010, 17, 434–440. [Google Scholar] [CrossRef]
- Gehringer, M.M.; Kewada, V.; Coates, N.; Downing, T.G. The use of Lepidium sativum in a plant bioassay system for the detection of microcystin-LR. Toxicon 2003, 41, 871–876. [Google Scholar] [CrossRef]
- Chen, J.; Song, L.; Dai, J.; Gan, N.; Liu, Z. Effects of microcystins on the growth and the activity of superoxide dismutase and peroxidase of rape (Brassica napus L.) and rice (Oryza sativa L.). Toxicon 2004, 43, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Máthé, C.; Beyer, D.; Erdődi, F.; Serfőző, Z.; Székvölgyi, L.; Vasas, G.; M-Hamvas, M.; Jámbrik, K.; Gonda, S.; Kiss, A.; et al. Microcystin-LR induces abnormal root development by altering microtubule organization in tissue-cultured common reed (Phragmites australis) plantlets. Aquat. Toxicol. 2009, 92, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Jámbrik, K.; Máthé, C.; Vasas, G.; Beyer, D.; Molnár, E.; Borbély, G.; M-Hamvas, M. Microcystin-LR induces chromatin alterations and modulates neutral single-strand-preferring nuclease activity in Phragmites australis. J. Plant Physiol. 2011, 168, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ye, J.; Zhang, H.; Jiang, X.; Zhang, Y.; Liu, Z. Freshwater toxic cyanobacteria induced DNA damage in apple (Malus pumila), rape (Brassica napus) and rice (Oryza sativa). J. Hazard. Mater. 2011, 190, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, C.C.; Azevedo, J.; Osório, H.; Vasconcelos, V.; Campos, A. Early physiological and biochemical responses of rice seedlings to low concentration of microcystin-LR. Ecotoxicology 2014, 23, 107–121. [Google Scholar] [CrossRef]
- Jampeetong, A.; Brix, H. Nitrogen nutrition of Salvinia natans: Effects of inorganic nitrogen form on growth, morphology, nitrate reductase activity and uptake kinetics of ammonium and nitrate. Aquat. Bot. 2009, 90, 67–73. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, S.; Wang, P.; Hou, J.; Li, W.; Zhang, W. Metabolic adaptations to ammonia-induced oxidative stress in leaves of the submerged macrophyte Vallisneria natans (Lour.) Hara. Aquat. Toxicol. 2008, 87, 88–98. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, C.; Jia, H.; Wei, L.; Yin, D. Effects of different nitrogen forms on growth and physiological indexes of Vallisneria natans. J. Plant Resour. Environ. 2006, 15, 48–51. [Google Scholar]
- Ma, Q.; Wang, Y.; Li, C.; Fang, P.; Zhou, J.; Fang, W. Influence of cyanobacterial bloom dominated by Planktothrix sp. and Cylindrospermopsis raciborskii on microflora structure of intestine, gill and culture enviroment of cultured Eriocheir sinensis. Mar. Fish 2021, 43, 595–606. [Google Scholar]
- Xue, Y.; Jiang, C.; Geng, J.; Xie, W.; Zhang, H.; Chen, X. Profiles of Bacterioplankton Based on qPCR and 16S rDNA High-throughput Sequencing during a Heavy Cyanobacterial Bloom in Zhushan Bay, Taihu Lake. Environ. Monit. Forewarn. 2017, 9, 19–23. [Google Scholar]
- Shao, K.; Gao, G.; Wang, Y.; Tang, X.; Qin, B. Vertical diversity of sediment bacterial communities in two different trophic states of the eutrophic Lake Taihu, China. J. Environ. Sci. China 2013, 25, 1186–1194. [Google Scholar] [CrossRef]
- Zheng, G.; Huang, H.; Tu, Z.; Zhang, L.; Jin, L.; Bai, Y.; Sun, R.; Zhang, Z. An Analysis of Correlation Between Bacterial Diversity and Water Environment in Poyang Lake. Acta Agric. Univ. Jiangxiensis 2017, 39, 549–558. [Google Scholar]
- Zhang, W.; Gu, P.; Zhu, W.; Wang, N.; Jiang, M.; He, J. Phenotype changes of cyanobacterial and microbial distribution characteristics of surface sediments in different periods of cyanobacterial blooms in taihu lake. Aquat. Ecol. 2020, 54, 591–607. [Google Scholar] [CrossRef]
- Huang, R.; Shen, F.; Luo, J.; Wang, S.; Tang, Q.; Xu, M.; Wu, Y.; Zhao, D. Effects of Withering of Cyanobacteria Bloom on Abundance and Community Composition of Ammonia-Oxidizing Bacteria in Surface Lake Sediments. J. Ecol. Rural Environ. 2015, 31, 334–339. [Google Scholar]
- Sun, X. The Influence of Zoobenthos on Community Structure and Function of Nitrogen Transformation Microbe in Sediment from Eutrophic Lakes. Ph.D. Thesis, Nanjing University, Nanjing, China, 2015. [Google Scholar]
- Peng, Y.; Lu, J.; Chen, H.; Xiao, L. Dynamic Changes of Nitrogen-Transforming and Phosphorus-Accumulating Bacteria Along with the Formation of Cyanobacterial Blooms. Environ. Sci. 2018, 39, 4938–4945. [Google Scholar]
- Jochimsen, E.M.; Carmichael, W.W.; An, J.; Cardo, D.M.; Cookson, S.T.; Holmes, C.E. Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. N. Engl. J. Med. 1998, 338, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Zhu, G.; Gao, G.; Zhang, Y.; Li, W.; Paerl, H.W.; Carmichael, W.W. A drinking water crisis in Lake Taihu, China: Linkage to climatic variability and lake management. Environ. Manag. 2010, 45, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Levy, S. Microcystis Rising: Why Phosphorus Reduction Isn’t Enough to Stop CyanoHABs. Environ. Health Perspect. 2017, 125, A34. [Google Scholar]
- Watson, S.B. Aquatic taste and odor: A primary signal of drinking-water integrity. J. Toxicol. Environ. Health 2004, 67, 1779–1795. [Google Scholar] [CrossRef]
- Chen, Y.; Qin, B.; Teubner, K.; Dokulil, M.T. Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China. J. Plankton. Res. 2003, 25, 445–453. [Google Scholar] [CrossRef]
- Sivonen, K.; Niemelä, S.I.; Niemi, R.M.; Lepistö, L.; Luoma, T.H.; Räsänen, L.A. Toxic cyanobacteria (blue-green algae) in Finnish fresh and coastal waters. Hydrobiologia 1990, 190, 267–275. [Google Scholar] [CrossRef]
- Turner, A.D.; Dhanji-Rapkova, M.; O’Neill, A.; Coates, L.; Lewis, A.; Lewis, K. Analysis of microcystins in cyanobacterial blooms from freshwater bodies in England. Toxins 2018, 10, 39. [Google Scholar] [CrossRef] [PubMed]
- Suput, D. Effects of microcystins, cyanobacterial toxins, on mammalian cells and organs. Acta Chim. Slov. 2011, 58, 708–716. [Google Scholar] [PubMed]
- Ohta, T.; Sueoka, E.; Lida, N.; Komori, A.; Suganuma, M.; Nishiwaki, R.; Tatematsu, M.; Kim, S.; Carmichael, W.W.; Fujiki, H. Nodularin, a potent inhibitor of protein phosphatases 1 and 2A, is a new environmental carcinogen in male F344 rat liver. Cancer Res. 1994, 54, 6402–6406. [Google Scholar] [PubMed]
- Terao, K.; Ohmori, S.; Igarashi, K.; Ohtani, I.; Watanabe, M.F.; Harada, K.I.; Ito, E.; Watanabe, M. Electron microscopic studies on experimental poisoning in mice induced by cylindrospermopsin isolated from blue-green alga Umezakia natans. Toxicon 1994, 32, 833–843. [Google Scholar] [CrossRef]
- Llewellyn, L.E. Saxitoxin, a toxic marine natural product that targets a multitude of receptors. Nat. Prod. Rep. 2006, 23, 200–222. [Google Scholar] [CrossRef]
- Lilleheil, G.; Andersen, R.A.; Skulberg, O.M.; Alexander, J. Effects of a homoanatoxin-a-containing extract from oscillato ria formosa (cyanophyceaebacteria) on neuromuscular transmission. Toxicon 1997, 35, 1275–1289. [Google Scholar] [CrossRef]
- Hyde, E.G.; Carmichael, W.W. a naturally occurring organophosphate, is an irreversible active site-directed inhibitor of acetylcholinesterase (EC 3.1. 1.7). J. Biochem. Toxicol. 1991, 6, 195–201. [Google Scholar] [CrossRef]
- Okle, O.; Stemmer, K.; Deschl, U.; Dietrich, D.R. L-BMAA induced ER stress and enhanced caspase 12 cleavage in human neuroblastoma SH-SY5Y cells at low nonexcitotoxic concentrations. Toxicol. Sci. 2013, 131, 217–224. [Google Scholar] [CrossRef]
- Meng, G.; Sun, Y.; Fu, W.; Guo, Z.; Xu, K. Microcystin-LR induces cytoskeleton system reorganization through hyperphosphorylation of tau and hsp27 via pp2a inhibition and subsequent activation of the p38 mapk signaling pathway in neuroendocrine (pc12) cells. Toxicology 2011, 290, 219–230. [Google Scholar] [CrossRef]
- Vesterkvist, P.S.M.; Meriluoto, J.A.O. Interaction between microcystins of different hydrophobicities and lipid monolayers. Toxicon 2003, 41, 349–355. [Google Scholar] [CrossRef]
- Ernst, B. Investigations on the Impact of Toxic Cyanobacteria on Fish as Exemplified by the Coregonids in Lake Ammersee. Ph.D. Thesis, University of Konstanz, Konstanz, Germany, 2008. [Google Scholar]
- Fischer, W.; Altheimer, S.; Cattori, V.; Meier, P.J.; Dietrich, D.R.; Hagenbuch, B. Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin—Science Direct. Toxicol. App. Pharm. 2005, 203, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Nishiwaki-Matsushima, R.; Ohta, T.; Nishiwaki, S.; Suganuma, M.; Kohyama, K.; Ishikawa, T.; Carmichael, W.W.; Fujiki, H. Liver tumor promotion by the cyanobacterial cyclic peptide toxin microcystin-LR. J. Cancer Res. Clin. 1992, 118, 420–424. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Odor Type | Compounds | |
|---|---|---|
| Taste odor | Sour | Citric acid, acetic acid |
| Sweet | Sucrose, glucose | |
| Bitter | Caffeine, quinine hydrochloride | |
| Salty | Sodium chloride | |
| Mouth/nose sensations | Spicy, greasy, spicy Metallic | Aluminium sulfate; menthol; methanol |
| Earthy/musty | Geosmin; 2-MIB; IPMP etc. | |
| Fragrant | 4-Nonylphenol; Decanal | |
| Grassy/woody | cis-3-Hexen-1-ol; cis-3-Hexenylacetic acid; β-Cyclocitric acid | |
| Fishy | 2-trans-4,7-cis-Decatrienal; 2,4-cis-Heptadienal | |
| Swamp | Dimethyl sulfide compounds; Isopropyl mercaptan | |
| Chemical | MTBE; 2-EDD etc. | |
| Chlorinenous | Free chlorine; Monochloramine; Dichloramine | |
| Medicinal | Bromophenol; Chlorophenols; Iodoform | |
| Toxins | Toxic Effect on Human Health |
|---|---|
 Microcystins | A potent hepatotoxin and tumor promoter, which inhibited protein phosphatase at the molecular level, resulting in hyperphosphorylation of critical regulatory proteins in the signal transduction process of cytoskeletal tissues, resulting in oxidative stress in liver, kidney, brain, and reproductive organs [65]. |
 Nodularin | It inhibits the activity of PP-1 and PP-2A, and has tumor-promoting activity, which is considered carcinogenic [66]. |
 Cylindrospermopsin | Also known as hepatotoxin, it is also harmful to other organs such as the thymus, kidney, and heart [67]. |
 Saxitoxins | A potent neurotoxin, which can be used as an antagonist of voltage-gated sodium channels to motor nerves, causes conduction defects and leads to respiratory paralysis [68]. |
 Anatoxin-a | An effective depolarizing neuromuscular blocker, these toxins irreversibly bind to the alkali acetylcholine receptor on the motor nerve endplate, and continuously stimulate muscle cells, thus leading to muscle paralysis [69]. |
 Anatoxin-a(s) | A cholinesterase inhibitor that leads to conduction disturbance and asphyxia death [70]. Anatoxin-a(s) poisoning symptoms include muscle weakness, convulsion, respiratory distress and death due to respiratory failure. |
 β-N-methylamino-L-alanine, (BMAA) | A developmental neurotoxin, maybe a factor in the increased incidence of Amyotrophic lateral sclerosis (ALS) and Parkinson’s dementia complex (PDC) in Guam [71]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Liu, J.; Xiao, Y.; Zhang, Y.; Yu, Y.; Zheng, Z.; Liu, Y.; Li, Q. The Impact of Cyanobacteria Blooms on the Aquatic Environment and Human Health. Toxins 2022, 14, 658. https://doi.org/10.3390/toxins14100658
Zhang W, Liu J, Xiao Y, Zhang Y, Yu Y, Zheng Z, Liu Y, Li Q. The Impact of Cyanobacteria Blooms on the Aquatic Environment and Human Health. Toxins. 2022; 14(10):658. https://doi.org/10.3390/toxins14100658
Chicago/Turabian StyleZhang, Weizhen, Jing Liu, Yunxing Xiao, Yumiao Zhang, Yangjinzhi Yu, Zheng Zheng, Yafeng Liu, and Qi Li. 2022. "The Impact of Cyanobacteria Blooms on the Aquatic Environment and Human Health" Toxins 14, no. 10: 658. https://doi.org/10.3390/toxins14100658
APA StyleZhang, W., Liu, J., Xiao, Y., Zhang, Y., Yu, Y., Zheng, Z., Liu, Y., & Li, Q. (2022). The Impact of Cyanobacteria Blooms on the Aquatic Environment and Human Health. Toxins, 14(10), 658. https://doi.org/10.3390/toxins14100658