Folding Control in the Path of Type 5 Secretion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
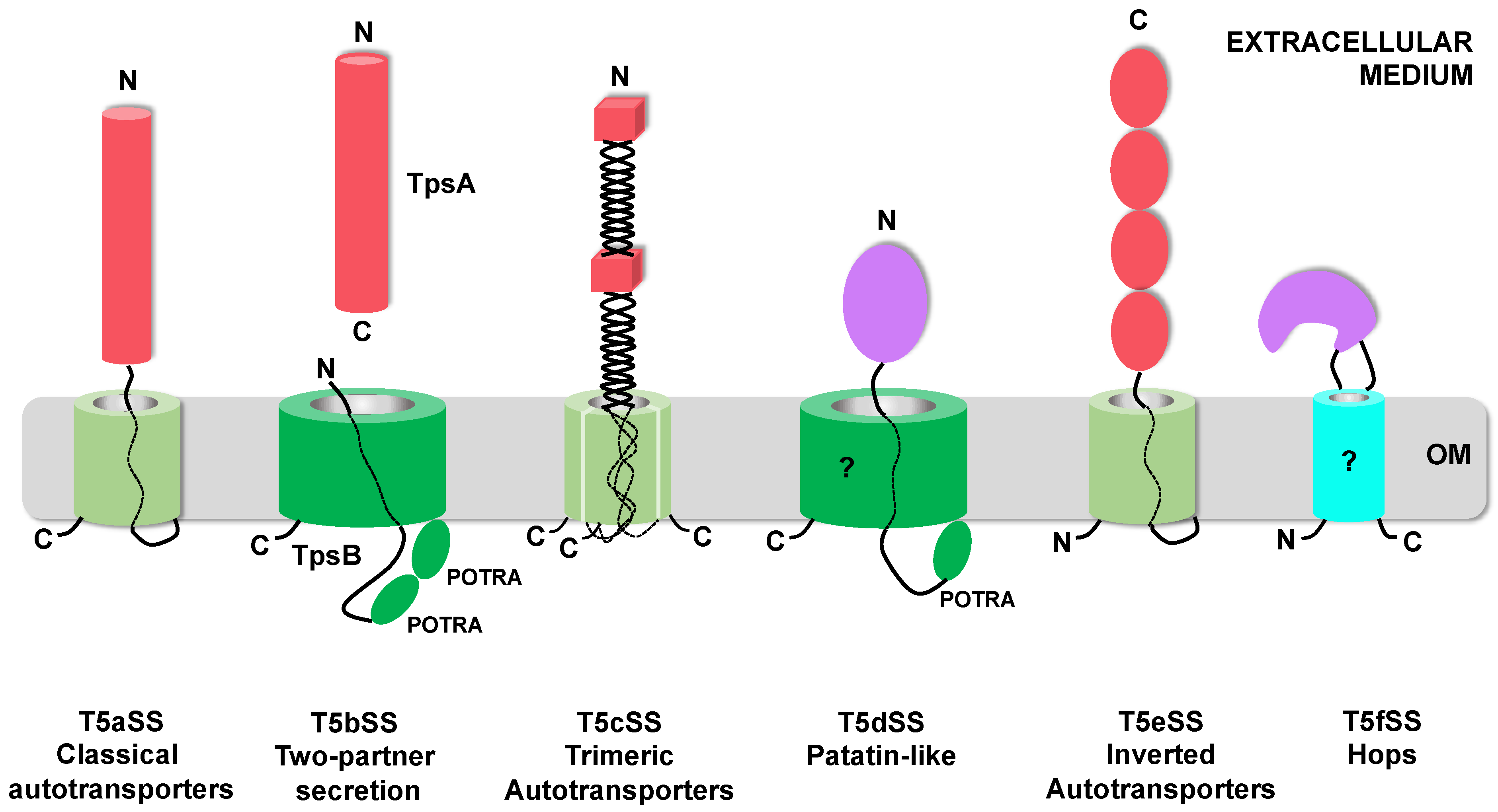
1. Introduction

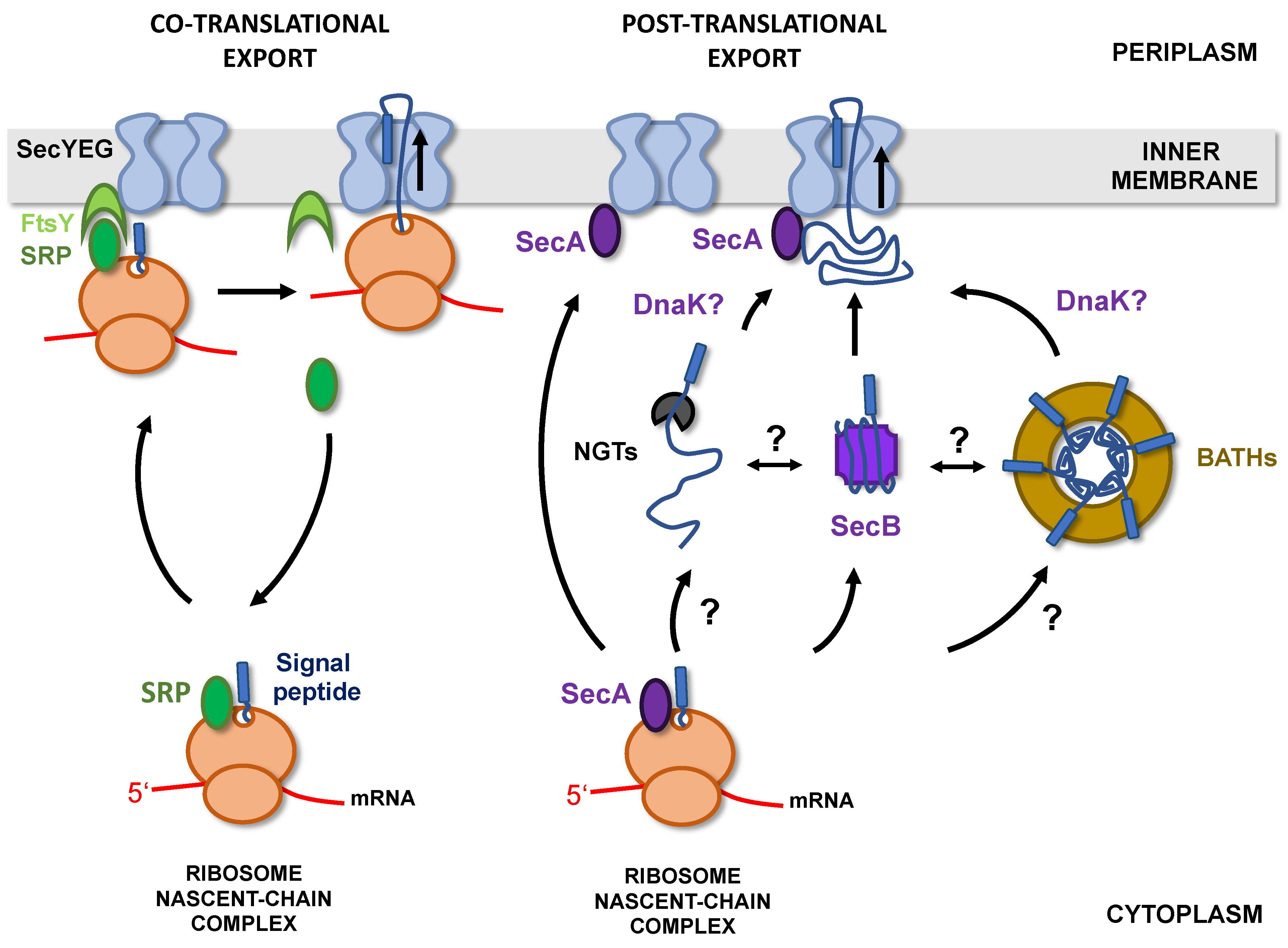
2. Staying Unfolded in the Cytoplasm
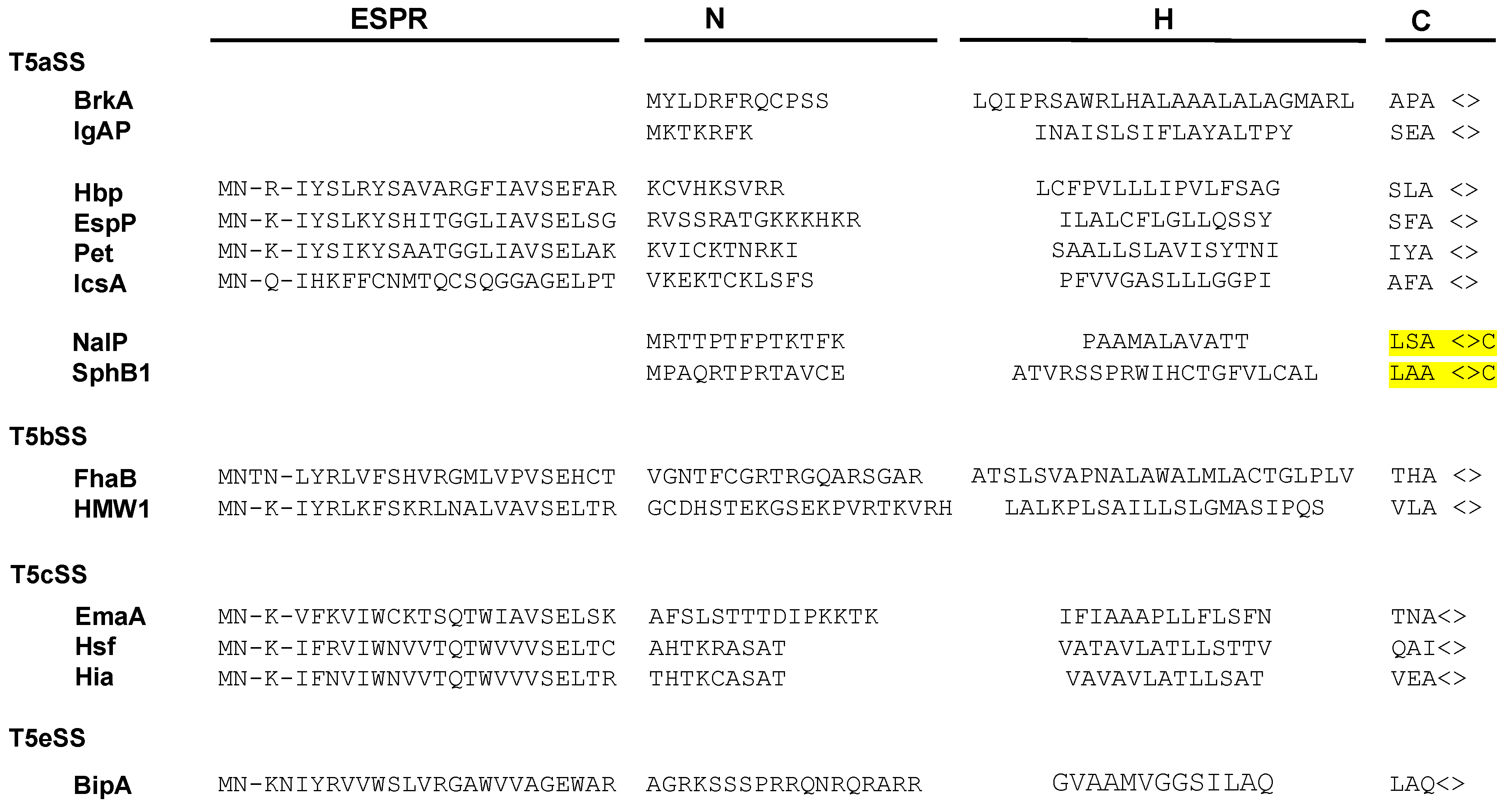
2.1. Targeting Pathways to the Inner Membrane
2.2. Cytoplasmic Glycosylation and Folding
3. Periplasmic Transit
3.1. Transient Anchoring to the Inner Membrane
3.2. Role of Periplasmic Factors in Maintaining Secretion-Competent Conformation
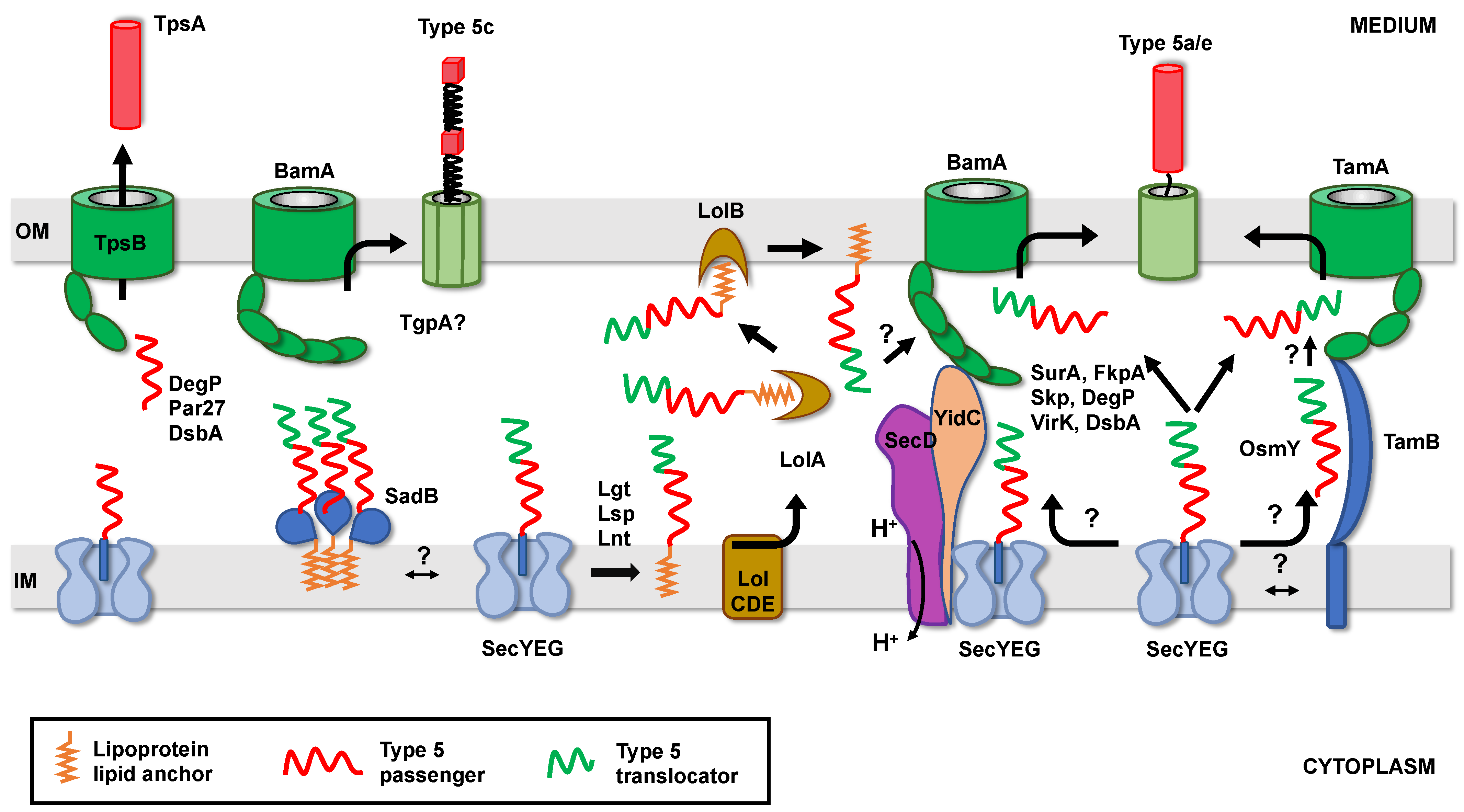
4. Translocator Domain Insertion and Folding in the Outer Membrane
4.1. Role of the BAM Complex
4.2. Role of the TAM Complex
5. Passenger Domain Secretion and Folding in the Extracellular Space
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhoite, S.; van Gerven, N.; Chapman, M.R.; Remaut, H. Curli Biogenesis: Bacterial Amyloid Assembly by the Type VIII Secretion Pathway. EcoSal Plus. 2019, 8. [Google Scholar] [CrossRef]
- Cherrak, Y.; Flaugnatti, N.; Durand, E.; Journet, L.; Cascales, E. Structure and Activity of the Type VI Secretion System. EcoSal Plus. 2019, 7, 329–342. [Google Scholar]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, Structure, Function and Regulation of Type III Secretion Systems. Nat. Rev. Microbiol. 2017, 15, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Gorasia, D.G.; Veith, P.D.; Reynolds, E.C. The Type IX Secretion System: Advances in Structure, Function and Organisation. Microorganisms 2020, 8, 1173. [Google Scholar] [CrossRef]
- Grossman, A.S.; Mauer, T.J.; Forest, K.T.; Goodrich-Blair, H. Description of a Widespread Bacterial Secretion System with Chemically Diverse Substrates. bioRxiv 2020. [Google Scholar] [CrossRef]
- Kanonenberg, K.; Spitz, O.; Erenburg, I.N.; Beer, T.; Schmitt, L. Type I Secretion System-It Takes Three and a Substrate. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef] [PubMed]
- Korotkov, K.V.; Sandkvist, M. Architecture, Function, and Substrates of the Type II Secretion System. EcoSal Plus. 2019, 8, 227–244. [Google Scholar]
- Palmer, T.; Finney, A.; Saha, C.K.; Atkinson, G.C.; Sargent, F. A Holin/Peptidoglycan Hydrolase-dependent Protein Secretion System. Mol. Microbiol. 2021, 115, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Grohmann, E.; Christie, P.J.; Waksman, G.; Backert, S. Type IV Secretion in Gram-negative and Gram-positive Bacteria. Mol. Microbiol. 2018, 107, 455–471. [Google Scholar] [CrossRef]
- Tran, H.R.; Grebenc, D.W.; Klein, T.A.; Whitney, J.C. Bacterial Type VII Secretion: An Important Player in Host-microbe and Microbe-microbe Interactions. Mol. Microbiol. 2021, 115, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Abby, S.S.; Cury, J.; Guglielmini, J.; Néron, B.; Touchon, M.; Rocha, E.P. Identification of Protein Secretion Systems in Bacterial Genomes. Sci. Rep. 2016, 6, 23080. [Google Scholar] [CrossRef]
- Drobnak, I.; Braselmann, E.; Chaney, J.L.; Leyton, D.L.; Bernstein, H.D.; Lithgow, T.; Luirink, J.; Nataro, J.P.; Clark, P.L. Of Linkers and Autochaperones: An Unambiguous Nomenclature to Identify Common and Uncommon Themes for Autotransporter Secretion. Mol. Microbiol. 2015, 95, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Doyle, M.T.; Bernstein, H.D. BamA Forms a Translocation Channel for Polypeptide Export Across the Bacterial Outer Membrane. Mol. Cell 2021. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, D.; Rydmark, M.O.; Pang, Y.T.; Lundquist, K.P.; Linke, D.; Gumbart, J.C. BamA is Required for Autotransporter Secretion. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129581. [Google Scholar] [CrossRef]
- Meuskens, I.; Saragliadis, A.; Leo, J.C.; Linke, D. Type V Secretion Systems: An Overview of Passenger Domain Functions. Front. Microbiol. 2019, 10, 1163. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Charles, I.G.; Fairweather, N.F.; Isaacs, N.W. Structure of Bordetella pertussis Virulence Factor P.69 Pertactin. Nature 1996, 381, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Oomen, C.J.; van Ulsen, P.; van Gelder, P.; Feijen, M.; Tommassen, J.; Gros, P. Structure of the Translocator Domain of a Bacterial Autotransporter. EMBO J. 2004, 23, 1257–1266. [Google Scholar] [CrossRef]
- Clantin, B.; Hodak, H.; Willery, E.; Locht, C.; Jacob-Dubuisson, F.; Villeret, V. The Crystal Structure of Filamentous Hemagglutinin Secretion Domain and Its Implications for the Two-partner Secretion Pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 6194–6199. [Google Scholar] [CrossRef]
- Clantin, B.; Delattre, A.S.; Rucktooa, P.; Saint, N.; Méli, A.C.; Locht, C.; Jacob-Dubuisson, F.; Villeret, V. Structure of the Membrane Protein FhaC: A Member of the Omp85-TpsB Transporter Superfamily. Science 2007, 317, 957–961. [Google Scholar] [CrossRef]
- Nummelin, H.; Merckel, M.C.; Leo, J.C.; Lankinen, H.; Skurnik, M.; Goldman, A. The Yersinia Adhesin YadA Collagen-binding Domain Structure is a Novel Left-handed Parallel Beta-roll. EMBO J. 2004, 23, 701–711. [Google Scholar] [CrossRef]
- Meng, G.; Surana, N.K.; St Geme, J.W., III; Waksman, G. Structure of the Outer Membrane Translocator Domain of the Haemophilus influenzae Hia Trimeric Autotransporter. EMBO J. 2006, 25, 2297–2304. [Google Scholar] [CrossRef] [PubMed]
- Da Mata Madeira, P.V.; Zouhir, S.; Basso, P.; Neves, D.; Laubier, A.; Salacha, R.; Bleves, S.; Faudry, E.; Contreras-Martel, C.; Dessen, A. Structural Basis of Lipid Targeting and Destruction by the Type V Secretion System of Pseudomonas aeruginosa. J. Mol. Biol. 2016, 428, 1790–1803. [Google Scholar] [CrossRef]
- Hamburger, Z.A.; Brown, M.S.; Isberg, R.R.; Bjorkman, P.J. Crystal Structure of Invasin: A Bacterial Integrin-binding Protein. Science 1999, 286, 291–295. [Google Scholar] [CrossRef]
- Fairman, J.W.; Dautin, N.; Wojtowicz, D.; Wei, L.; Noinaj, N.; Barnard, T.J.; Przytycka, T.; Cherezov, V.; Buchanan, S.K. Crystal Structure of the Transmembrane Domain from the Bacterial Adhesins Intimin and Invasin. Structure 2012, 20, 1233–1243. [Google Scholar] [CrossRef]
- Hage, N.; Howard, T.; Phillips, C.; Brassington, C.; Overman, R.; Debreczeni, J.; Gellert, P.; Stolnik, S.; Winkler, G.S.; Falcone, F.H. Structural Basis of Lewis(b) Antigen Binding by the Helicobacter pylori Adhesin BabA. Sci. Adv. 2015, 1, e1500315. [Google Scholar] [CrossRef]
- Leyton, D.L.; Rossiter, A.E.; Henderson, I.R. From Self-sufficiency to Dependence: Mechanisms and Factors Important for Autotransporter Biogenesis. Nat. Rev. Microbiol. 2012, 10, 213–225. [Google Scholar] [CrossRef]
- Madasu, Y.; Suarez, C.; Kast, D.J.; Kovar, D.R.; Dominguez, R. Rickettsia Sca2 has Evolved Formin-like Activity through a Different Molecular Mechanism. Proc. Natl. Acad. Sci. USA 2013, 110, E2677–E2686. [Google Scholar] [CrossRef]
- Leo, J.C.; Oberhettinger, P.; Schütz, M.; Linke, D. The Inverse Autotransporter Family: Intimin, Invasin and Related Proteins. Int. J. Med. Microbiol. 2015, 305, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Kiessling, A.R.; Malik, A.; Goldman, A. Recent Advances in the Understanding of Trimeric Autotransporter Adhesins. Med. Microbiol. Immunol. 2020, 209, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Guérin, J.; Bigot, S.; Schneider, R.; Buchanan, S.K.; Jacob-Dubuisson, F. Two-partner Secretion: Combining Efficiency and Simplicity in the Secretion of Large Proteins for Bacteria-host and Bacteria-bacteria Interactions. Front. Cell. Infect. Microbiol. 2017, 7, 148. [Google Scholar] [CrossRef]
- Salacha, R.; Kovacić, F.; Brochier-Armanet, C.; Wilhelm, S.; Tommassen, J.; Filloux, A.; Voulhoux, R.; Bleves, S. The Pseudomonas aeruginosa Patatin-like Protein PlpD is the Archetype of a Novel Type V Secretion System. Environ. Microbiol. 2010, 12, 1498–1512. [Google Scholar] [PubMed]
- Coppens, F.; Castaldo, G.; Debraekeleer, A.; Subedi, S.; Moonens, K.; Lo, A.; Remaut, H. Hop-family Helicobacter Outer Membrane Adhesins Form a Novel Class of Type 5-like Secretion Proteins with an Interrupted Beta-barrel Domain. Mol. Microbiol. 2018, 110, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Voulhoux, R.; Bos, M.P.; Geurtsen, J.; Mols, M.; Tommassen, J. Role of a Highly Conserved Bacterial Protein in Outer Membrane Protein Assembly. Science 2003, 299, 262–265. [Google Scholar] [CrossRef]
- Ricci, D.P.; Silhavy, T.J. Outer Membrane Protein Insertion by the β-barrel Assembly Machine. EcoSal Plus. 2019, 8, 91–101. [Google Scholar]
- Von Heijne, G. Signal Sequences. The Limits of Variation. J. Mol. Biol. 1985, 184, 99–105. [Google Scholar] [CrossRef]
- Smets, D.; Loos, M.S.; Karamanou, S.; Economou, A. Protein Transport across the Bacterial Plasma Membrane by the Sec Pathway. Protein J. 2019, 38, 262–273. [Google Scholar] [CrossRef]
- Steinberg, R.; Knüpffer, L.; Origi, A.; Asti, R.; Koch, H.G. Co-translational Protein Targeting in Bacteria. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Loos, M.S.; Ramakrishnan, R.; Vranken, W.; Tsirigotaki, A.; Tsare, E.P.; Zorzini, V.; Geyter, J.; Yuan, B.; Tsamardinos, I.; Klappa, M.; et al. Structural Basis of the Subcellular Topology Landscape of Escherichia coli. Front. Microbiol. 2019, 10, 1670. [Google Scholar] [CrossRef] [PubMed]
- Castanié-Cornet, M.P.; Bruel, N.; Genevaux, P. Chaperone Networking Facilitates Protein Targeting to the Bacterial Cytoplasmic Membrane. Biochim. Biophys. Acta 2014, 1843, 1442–1456. [Google Scholar] [CrossRef]
- Desvaux, M.; Cooper, L.M.; Filenko, N.A.; Scott-Tucker, A.; Turner, S.M.; Cole, J.A.; Henderson, I.R. The Unusual Extended Signal Peptide Region of the Type V Secretion System is Phylogenetically Restricted. FEMS Microbiol. Lett. 2006, 264, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Szabady, R.L.; Peterson, J.H.; Skillman, K.M.; Bernstein, H.D. An Unusual Signal Peptide Facilitates Late Steps in the Biogenesis of a Bacterial Autotransporter. Proc. Natl. Acad. Sci. USA 2005, 102, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Ruiz, T.; Mintz, K.P. The Extended Signal Peptide of the Trimeric Autotransporter EmaA of Aggregatibacter actinomycetemcomitans Modulates Secretion. J. Bacteriol. 2011, 193, 6983–6994. [Google Scholar] [CrossRef]
- Jong, W.S.; Luirink, J. The Conserved Extension of the Hbp Autotransporter Signal Peptide Does Not Determine Targeting Pathway Specificity. Biochem. Biophys. Res. Commun. 2008, 368, 522–527. [Google Scholar] [CrossRef]
- Leyton, D.L.; de Luna, M.D.; Sevastsyanovich, Y.R.; Tveen Jensen, K.; Browning, D.F.; Scott-Tucker, A.; Henderson, I.R. The Unusual Extended Signal Peptide Region is not Required for Secretion and Function of an Escherichia coli Autotransporter. FEMS Microbiol. Lett. 2010, 311, 133–139. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Peterson, J.H.; Woolhead, C.A.; Bernstein, H.D. Basic Amino Acids in a Distinct Subset of Signal Peptides Promote Interaction with the Signal Recognition Particle. J. Biol. Chem. 2003, 278, 46155–46162. [Google Scholar] [CrossRef] [PubMed]
- Pina-Pedrero, S.; Olvera, À.; Bensaid, A. The Extended Leader Peptide of Haemophilus parasuis Trimeric Autotransporters Conditions Their Protein Expression in Escherichia coli. Protein Expr. Purif. 2017, 133, 15–24. [Google Scholar] [CrossRef]
- Brandon, L.D.; Goehring, N.; Janakiraman, A.; Yan, A.W.; Wu, T.; Beckwith, J.; Goldberg, M.B. IcsA, a Polarly Localized Autotransporter with an Atypical Signal Peptide, Uses the Sec Apparatus for Secretion, although the Sec Apparatus is Circumferentially Distributed. Mol. Microbiol. 2003, 50, 45–60. [Google Scholar] [CrossRef]
- Desvaux, M.; Scott-Tucker, A.; Turner, S.M.; Cooper, L.M.; Huber, D.; Nataro, J.P.; Henderson, I.R. A Conserved Extended Signal Peptide Region Directs Posttranslational Protein Translocation via a Novel Mechanism. Microbiology 2007, 153, 59–70. [Google Scholar] [CrossRef]
- Peterson, J.H.; Szabady, R.L.; Bernstein, H.D. An Unusual Signal Peptide Extension Inhibits the Binding of Bacterial Presecretory Proteins to the Signal Recognition Particle, Trigger Factor, and the SecYEG Complex. J. Biol. Chem. 2006, 281, 9038–9048. [Google Scholar] [CrossRef]
- Ariosa, A.; Lee, J.H.; Wang, S.; Saraogi, I.; Shan, S.O. Regulation by a Chaperone Improves Substrate Selectivity during Cotranslational Protein Targeting. Proc. Natl. Acad. Sci. USA 2015, 112, E3169–E3178. [Google Scholar] [CrossRef]
- Zhang, X.; Rashid, R.; Wang, K.; Shan, S.O. Sequential Checkpoints Govern Substrate Selection during Cotranslational Protein Targeting. Science 2010, 328, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, N.; Moser, M.; Koch, H.G.; Schimz, K.L.; Willery, E.; Locht, C.; Jacob-Dubuisson, F.; Müller, M. Membrane Targeting of a Bacterial Virulence Factor Harbouring an Extended Signal Peptide. J. Mol. Microbiol. Biotechnol. 2004, 8, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Sijbrandi, R.; Urbanus, M.L.; ten Hagen-Jongman, C.M.; Bernstein, H.D.; Oudega, B.; Otto, B.R.; Luirink, J. Signal Recognition Particle (SRP)-mediated Targeting and Sec-dependent Translocation of an Extracellular Escherichia coli protein. J. Biol. Chem. 2003, 278, 4654–4659. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Ruiz, T.; Mintz, K.P. Characterization of the Secretion Pathway of the Collagen Adhesin EmaA of Aggregatibacter actinomycetemcomitans. Mol. Oral Microbiol. 2012, 27, 382–396. [Google Scholar] [CrossRef]
- Janakiraman, A.; Fixen, K.R.; Gray, A.N.; Niki, H.; Goldberg, M.B. A Genome-scale Proteomic Screen Identifies a Role for DnaK in Chaperoning of Polar Autotransporters in Shigella. J. Bacteriol. 2009, 191, 6300–6311. [Google Scholar] [CrossRef] [PubMed]
- Calloni, G.; Chen, T.; Schermann, S.M.; Chang, H.C.; Genevaux, P.; Agostini, F.; Tartaglia, G.G.; Hayer-Hartl, M.; Hartl, F.U. DnaK Functions as a Central Hub in the E. coli Chaperone Network. Cell Rep. 2012, 1, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Huber, D.; Boyd, D.; Xia, Y.; Olma, M.H.; Gerstein, M.; Beckwith, J. Use of Thioredoxin as a Reporter to Identify a Subset of Escherichia coli Signal Sequences That Promote Signal Recognition Particle-dependent Translocation. J. Bacteriol. 2005, 187, 2983–2991. [Google Scholar] [CrossRef]
- Braun, V.; Hantke, K. Lipoproteins: Structure, Function, Biosynthesis. Subcell Biochem. 2019, 92, 39–77. [Google Scholar]
- Watanabe, T.; Hayashi, S.; Wu, H.C. Synthesis and Export of the Outer Membrane Lipoprotein in Escherichia coli Mutants Defective in Generalized Protein Export. J. Bacteriol. 1988, 170, 4001–4007. [Google Scholar] [CrossRef]
- Fröderberg, L.; Houben, E.N.; Baars, L.; Luirink, J.; de Gier, J.W. Targeting and Translocation of Two Lipoproteins in Escherichia coli via the SRP/Sec/YidC Pathway. J. Biol. Chem. 2004, 279, 31026–31032. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, D.; Kuhn, A. YidC-mediated Membrane Insertion. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Junker, M.; Schuster, C.C.; McDonnell, A.V.; Sorg, K.A.; Finn, M.C.; Berger, B.; Clark, P.L. Pertactin Beta-helix Folding Mechanism Suggests Common Themes for the Secretion and Folding of Autotransporter Proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 4918–4923. [Google Scholar] [CrossRef]
- Knudsen, S.K.; Stensballe, A.; Franzmann, M.; Westergaard, U.B.; Otzen, D.E. Effect of Glycosylation on the Extracellular Domain of the Ag43 Bacterial Autotransporter: Enhanced Stability and Reduced Cellular Aggregation. Biochem. J. 2008, 412, 563–577. [Google Scholar] [CrossRef][Green Version]
- Bowman, M.A.; Riback, J.A.; Rodriguez, A.; Guo, H.; Li, J.; Sosnick, T.R.; Clark, P.L. Properties of Protein Unfolded States Suggest Broad Selection for Expanded Conformational Ensembles. Proc. Natl. Acad. Sci. USA 2020, 117, 23356–23364. [Google Scholar] [CrossRef]
- Lindenthal, C.; Elsinghorst, E.A. Identification of a Glycoprotein Produced by Enterotoxigenic Escherichia coli. Infect. Immun. 1999, 67, 4084–4091. [Google Scholar] [CrossRef] [PubMed]
- Benz, I.; Schmidt, M.A. Glycosylation with Heptose Residues Mediated by the aah Gene Product is Essential for Adherence of the AIDA-I Adhesin. Mol. Microbiol. 2001, 40, 1403–1413. [Google Scholar] [CrossRef]
- Sherlock, O.; Dobrindt, U.; Jensen, J.B.; Munk Vejborg, R.; Klemm, P. Glycosylation of the Self-recognizing Escherichia coli Ag43 Autotransporter Protein. J. Bacteriol. 2006, 188, 1798–1807. [Google Scholar] [CrossRef]
- Lu, Q.; Yao, Q.; Xu, Y.; Li, L.; Li, S.; Liu, Y.; Gao, W.; Niu, M.; Sharon, M.; Ben-Nissan, G.; et al. An Iron-containing Dodecameric Heptosyltransferase Family Modifies Bacterial Autotransporters in Pathogenesis. Cell Host Microbe 2014, 16, 351–363. [Google Scholar] [CrossRef]
- Fleckenstein, J.M.; Roy, K.; Fischer, J.F.; Burkitt, M. Identification of a Two-partner Secretion Locus of Enterotoxigenic Escherichia coli. Infect. Immun. 2006, 74, 2245–2258. [Google Scholar] [CrossRef]
- Grass, S.; Buscher, A.Z.; Swords, W.E.; Apicella, M.A.; Barenkamp, S.J.; Ozchlewski, N.; St Geme, J.W., III. The Haemophilus influenzae HMW1 Adhesin is Glycosylated in a Process That Requires HMW1C and Phosphoglucomutase, an Enzyme Involved in Lipooligosaccharide Biosynthesis. Mol. Microbiol. 2003, 48, 737–751. [Google Scholar] [CrossRef]
- Rempe, K.A.; Spruce, L.A.; Porsch, E.A.; Seeholzer, S.H.; Nørskov-Lauritsen, N.; St. Geme, J.W., 3rd. Unconventional N-linked Glycosylation Promotes Trimeric Autotransporter Function in Kingella kingae and Aggregatibacter aphrophilus. mBio 2015, 6, e01206-15. [Google Scholar] [CrossRef]
- Tang, G.; Mintz, K.P. Glycosylation of the Collagen Adhesin EmaA of Aggregatibacter actinomycetemcomitans is Dependent upon the Lipopolysaccharide Biosynthetic Pathway. J. Bacteriol. 2010, 192, 1395–1404. [Google Scholar] [CrossRef]
- Choi, K.J.; Grass, S.; Paek, S.; St Geme, J.W., 3rd; Yeo, H.J. The Actinobacillus pleuropneumoniae HMW1C-like Glycosyltransferase Mediates N-linked Glycosylation of the Haemophilus influenzae HMW1 Adhesin. PLoS ONE 2010, 5, e15888. [Google Scholar] [CrossRef]
- Naegeli, A.; Neupert, C.; Fan, Y.Y.; Lin, C.W.; Poljak, K.; Papini, A.M.; Schwarz, F.; Aebi, M. Molecular Analysis of an Alternative N-glycosylation Machinery by Functional Transfer from Actinobacillus pleuropneumoniae to Escherichia coli. J. Biol. Chem. 2014, 289, 2170–2179. [Google Scholar] [CrossRef] [PubMed]
- Bhakta, S.; Akbar, S.; Sengupta, J. Cryo-EM structures Reveal Relocalization of MetAP in the Presence of Other Protein Biogenesis Factors at the Ribosomal Tunnel Exit. J. Mol. Biol. 2019, 431, 1426–1439. [Google Scholar] [CrossRef]
- Sandikci, A.; Gloge, F.; Martinez, M.; Mayer, M.P.; Wade, R.; Bukau, B.; Kramer, G. Dynamic Enzyme Docking to the Ribosome Coordinates N-terminal Processing with Polypeptide Folding. Nat. Struct. Mol. Biol. 2013, 20, 843–850. [Google Scholar] [CrossRef]
- Song, Q.; Wu, Z.; Fan, Y.; Song, W.; Zhang, P.; Wang, L.; Wang, F.; Xu, Y.; Wang, P.G.; Cheng, J. Production of Homogeneous Glycoprotein with Multisite Modifications by an Engineered N-glycosyltransferase Mutant. J. Biol. Chem. 2017, 292, 8856–8863. [Google Scholar] [CrossRef]
- Yakovlieva, L.; Ramírez-Palacios, C.; Marrink, S.J.; Walvoort, M.T.C. Semiprocessive Hyperglycosylation of Adhesin by Bacterial Protein N-glycosyltransferases. ACS Chem. Biol. 2021, 16, 165–175. [Google Scholar] [CrossRef]
- Charbonneau, M.E.; Girard, V.; Nikolakakis, A.; Campos, M.; Berthiaume, F.; Dumas, F.; Lépine, F.; Mourez, M. O-linked Glycosylation Ensures the Normal Conformation of the Autotransporter Adhesin Involved in Diffuse Adherence. J. Bacteriol. 2007, 189, 8880–8889. [Google Scholar] [CrossRef]
- Yao, Q.; Lu, Q.; Wan, X.; Song, F.; Xu, Y.; Hu, M.; Zamyatina, A.; Liu, X.; Huang, N.; Zhu, P.; et al. A Structural Mechanism for Bacterial Autotransporter Glycosylation by a Dodecameric Heptosyltransferase Family. eLife 2014, 13, 3. [Google Scholar]
- Moormann, C.; Benz, I.; Schmidt, M.A. Functional Substitution of the TibC Protein of Enterotoxigenic Escherichia coli Strains for the Autotransporter Adhesin Heptosyltransferase of the AIDA System. Infect. Immun. 2002, 70, 2264–2270. [Google Scholar] [CrossRef] [PubMed]
- Charbonneau, M.E.; Côté, J.P.; Haurat, M.F.; Reiz, B.; Crépin, S.; Berthiaume, F.; Dozois, C.M.; Feldman, M.F.; Mourez, M. A Structural Motif is the Recognition Site for a New Family of Bacterial Protein O-glycosyltransferases. Mol. Microbiol. 2012, 83, 894–907. [Google Scholar] [CrossRef]
- McCann, J.R.; St. Geme, J.W., III. The HMW1C-like Glycosyltransferases—An Enzyme Family with a Sweet Tooth for Simple Sugars. PLoS Pathog. 2014, 10, e1003977. [Google Scholar] [CrossRef] [PubMed]
- Charbonneau, M.E.; Mourez, M. The Escherichia coli AIDA-I Autotransporter Undergoes Cytoplasmic Glycosylation Independently of Export. Res. Microbiol. 2008, 159, 537–544. [Google Scholar] [CrossRef]
- Lundquist, K.; Billings, E.; Bi, M.; Wellnitz, J.; Noinaj, N. The Assembly of β-barrel Membrane Proteins by BAM and SAM. Mol. Microbiol. 2020. [Google Scholar] [CrossRef]
- Tomasek, D.; Kahne, D. The Assembly of β-barrel Outer Membrane Proteins. Curr. Opin. Microbiol. 2021, 60, 16–23. [Google Scholar] [CrossRef]
- Stubenrauch, C.J.; Lithgow, T. The TAM: A Translocation and Assembly Module of the β-Barrel Assembly Machinery in Bacterial Outer Membranes. EcoSal Plus. 2019, 8, 103–111. [Google Scholar]
- Stockbauer, K.E.; Fuchslocher, B.; Miller, J.F.; Cotter, P.A. Identification and Characterization of BipA, a Bordetella Bvg-intermediate Phase Protein. Mol. Microbiol. 2001, 39, 65–78. [Google Scholar] [CrossRef]
- Jong, W.S.; ten Hagen-Jongman, C.M.; Ruijter, E.; Orru, R.V.; Genevaux, P.; Luirink, J. YidC is Involved in the Biogenesis of the Secreted Autotransporter Hemoglobin Protease. J. Biol. Chem. 2010, 285, 39682–39690. [Google Scholar] [CrossRef] [PubMed]
- Fürst, M.; Zhou, Y.; Merfort, J.; Müller, M. Involvement of PpiD in Sec-dependent Protein Translocation. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 273–280. [Google Scholar] [CrossRef]
- Jauss, B.; Petriman, N.A.; Drepper, F.; Franz, L.; Sachelaru, I.; Welte, T.; Steinberg, R.; Warscheid, B.; Koch, H.G. Noncompetitive Binding of PpiD and YidC to the SecYEG Translocon Expands the Global View on the SecYEG Interactome in Escherichia coli. J. Biol. Chem. 2019, 294, 19167–19183. [Google Scholar] [CrossRef]
- Braselmann, E.; Chaney, J.L.; Champion, M.M.; Clark, P.L. DegP Chaperone Suppresses Toxic Inner Membrane Translocation Intermediates. PLoS ONE 2016, 11, e0162922. [Google Scholar] [CrossRef]
- Grin, I.; Hartmann, M.D.; Sauer, G.; Hernandez Alvarez, B.; Schütz, M.; Wagner, S.; Madlung, J.; Macek, B.; Felipe-Lopez, A.; Hensel, M.; et al. Trimeric Lipoprotein Assists in Trimeric Autotransporter Biogenesis in Enterobacteria. J. Biol. Chem. 2014, 289, 7388–7398. [Google Scholar] [CrossRef] [PubMed]
- Josts, I.; Stubenrauch, C.J.; Vadlamani, G.; Mosbahi, K.; Walker, D.; Lithgow, T.; Grinter, R. The Structure of a Conserved Domain of TamB Reveals a Hydrophobic β Taco Fold. Structure 2017, 25, 1898–1906. [Google Scholar] [CrossRef] [PubMed]
- Selkrig, J.; Mosbahi, K.; Webb, C.T.; Belousoff, M.J.; Perry, A.J.; Well, T.J.; Morris, F.; Leyton, D.L.; Totsika, M.; Phan, M.D.; et al. Discovery of an Archetypal Protein Transport System in Bacterial Outer Membranes. Nat. Struct. Mol. Biol. 2012, 9, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.H.; Leyton, D.L.; Shiota, T.; Belousoff, M.J.; Noinaj, N.; Lu, J.; Holt, S.A.; Tan, K.; Selkrig, J.; Webb, C.T.; et al. Reconstitution of a Nanomachine Driving the Assembly of Proteins into Bacterial Outer Membranes. Nat. Commun. 2014, 5, 5078. [Google Scholar] [CrossRef]
- Babu, M.; Bundalovic-Torma, C.; Calmettes, C.; Phanse, S.; Zhang, Q.; Jiang, Y.; Minic, Z.; Kim, S.; Mehla, J.; Gagarinova, A.; et al. Global Landscape of Cell Envelope Protein Complexes in Escherichia coli. Nat. Biotechnol. 2018, 36, 103–112. [Google Scholar] [CrossRef]
- Alvira, S.; Watkins, D.W.; Troman, L.; Allen, W.J.; Lorriman, J.S.; Degliesposti, G.; Cohen, E.J.; Beeby, M.; Daum, B.; Gold, V.A.; et al. Inter-membrane Association of the Sec and BAM Translocons for Bacterial Outer-membrane Biogenesis. eLife 2020, 9, e60669. [Google Scholar] [CrossRef]
- Carlson, M.L.; Stacey, R.G.; Young, J.W.; Wason, I.S.; Zhao, Z.; Rattray, D.G.; Scott, N.; Kerr, C.H.; Babu, M.; Foster, L.J.; et al. Profiling the Escherichia coli Membrane Protein Interactome Captured in Peptidisc libraries. eLife 2019, 8, e46615. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, R.; Jin, F.; Liu, Y.; Yu, J.; Fu, X.; Chang, Z. A Supercomplex Spanning the Inner and Outer Membranes Mediates the Biogenesis of Beta-barrel Outer Membrane Proteins in Bacteria. J. Biol. Chem. 2016, 291, 16720–16729. [Google Scholar] [CrossRef]
- Sklar, J.G.; Wu, T.; Kahne, D.; Silhavy, T.J. Defining the Roles of the Periplasmic Chaperones SurA, Skp, and DegP in Escherichia coli. Genes Dev. 2007, 21, 2473–2484. [Google Scholar] [CrossRef]
- Purdy, G.E.; Fisher, C.R.; Payne, S.M. IcsA Surface Presentation in Shigella flexneri Requires the Periplasmic Chaperones DegP, Skp, and SurA. J. Bacteriol. 2007, 189, 5566–5573. [Google Scholar] [CrossRef]
- Ruiz-Perez, F.; Henderson, I.R.; Leyton, D.L.; Rossiter, A.E.; Zhang, Y.; Nataro, J.P. Roles of Periplasmic Chaperone Proteins in the Biogenesis of Serine Protease Autotransporters of Enterobacteriaceae. J. Bacteriol. 2009, 191, 6571–6583. [Google Scholar] [CrossRef]
- Sauri, A.; Soprova, Z.; Wickström, D.; de Gier, J.W.; Van der Schors, R.C.; Smit, A.B.; Jong, W.S.; Luirink, J. The Bam (Omp85) Complex is Involved in Secretion of the Autotransporter Haemoglobin Protease. Microbiology 2009, 155, 3982–3991. [Google Scholar] [CrossRef] [PubMed]
- Bodelón, G.; Marín, E.; Fernández, L.A. Role of Periplasmic Chaperones and BamA (YaeT/Omp85) in Folding and Secretion of Intimin from Enteropathogenic Escherichia coli Strains. J. Bacteriol. 2009, 191, 5169–5179. [Google Scholar] [CrossRef] [PubMed]
- Oberhettinger, P.; Schütz, M.; Leo, J.C.; Heinz, N.; Berger, J.; Autenrieth, I.B.; Linke, D. Intimin and Invasin Export Their C-terminus to the Bacterial Cell Surface Using an Inverse Mechanism Compared to Classical Autotransport. PLoS ONE 2012, 7, e47069. [Google Scholar] [CrossRef]
- Obi, I.R.; Francis, M.S. Demarcating SurA Activities Required for Outer Membrane Targeting of Yersinia pseudotuberculosis Adhesins. Infect. Immun. 2013, 81, 2296–2308. [Google Scholar] [CrossRef] [PubMed]
- Weirich, J.; Bräutigam, C.; Mühlenkamp, M.; Franz-Wachtel, M.; Macek, B.; Meuskens, I.; Skurnik, M.; Leskinen, K.; Bohn, E.; Autenrieth, I.; et al. Identifying Components Required for OMP Biogenesis as Novel Targets for Antiinfective Drugs. Virulence 2017, 8, 1170–1188. [Google Scholar] [CrossRef] [PubMed]
- Ieva, R.; Bernstein, H.D. Interaction of an Autotransporter Passenger Domain with BamA during Its Translocation Across the Bacterial Outer Membrane. Proc. Natl. Acad. Sci. USA 2009, 106, 19120–19125. [Google Scholar] [CrossRef] [PubMed]
- Ieva, R.; Tian, P.; Peterson, J.H.; Bernstein, H.D. Sequential and Spatially Restricted Interactions of Assembly Factors with an Autotransporter Beta Domain. Proc. Natl. Acad. Sci. USA 2011, 108, E383–E391. [Google Scholar] [CrossRef]
- Qu, J.; Mayer, C.; Behrens, S.; Holst, O.; Kleinschmidt, J.H. The Trimeric Periplasmic Chaperone Skp of Escherichia coli Forms 1:1 Complexes with Outer Membrane Proteins via Hydrophobic and Electrostatic Interactions. J. Mol. Biol. 2007, 374, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Pastrana, G.; Chavez-Dueñas, L.; Lanz-Mendoza, H.; Teter, K.; Navarro-García, F. VirK is a Periplasmic Protein Required for Efficient Secretion of Plasmid-encoded Toxin from Enteroaggregative Escherichia coli. Infect. Immun. 2012, 80, 2276–2285. [Google Scholar] [CrossRef] [PubMed]
- Jong, W.S.; ten Hagen-Jongman, C.M.; den Blaauwen, T.; Slotboom, D.J.; Tame, J.R.; Wickström, D.; de Gier, J.W.; Otto, B.R.; Luirink, J. Limited Tolerance Towards Folded Elements during Secretion of the Autotransporter Hbp. Mol. Microbiol. 2007, 63, 1524–1536. [Google Scholar] [CrossRef]
- Ruiz-Perez, F.; Henderson, I.R.; Nataro, J.P. Interaction of FkpA, a Peptidyl-prolyl Cis/Trans Isomerase with EspP Autotransporter Protein. Gut Microbes 2010, 1, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Hagan, C.L.; Kim, S.; Kahne, D. Reconstitution of Outer Membrane Protein Assembly from Purified Components. Science 2010, 328, 890–892. [Google Scholar] [CrossRef]
- Roman-Hernandez, G.; Peterson, J.H.; Bernstein, H.D. Reconstitution of Bacterial Autotransporter Assembly Using Purified Components. eLife 2014, 3, e04234. [Google Scholar] [CrossRef]
- Nakata, N.; Sasakawa, C.; Okada, N.; Tobe, T.; Fukuda, I.; Suzuki, T.; Komatsu, K.; Yoshikawa, M. Identification and Characterization of virK, a Virulence-associated Large Plasmid Gene Essential for Intercellular Spreading of Shigella flexneri. Mol. Microbiol. 1992, 6, 2387–2395. [Google Scholar] [CrossRef]
- Yan, Z.; Hussain, S.; Wang, X.; Bernstein, H.D.; Bardwell, J.C.A. Chaperone OsmY Facilitates the Biogenesis of a Major Family of Autotransporters. Mol. Microbiol. 2019, 112, 1373–1387. [Google Scholar] [CrossRef] [PubMed]
- Volokhina, E.B.; Grijpstra, J.; Stork, M.; Schilders, I.; Tommassen, J.; Bos, M.P. Role of the Periplasmic Chaperones Skp, SurA, and DegQ in Outer Membrane Protein Biogenesis in Neisseria meningitidis. J. Bacteriol. 2011, 193, 1612–1621. [Google Scholar] [CrossRef]
- Baud, C.; Hodak, H.; Willery, E.; Drobecq, H.; Locht, C.; Jamin, M.; Jacob-Dubuisson, F. Role of DegP for Two-partner Secretion in Bordetella. Mol. Microbiol. 2009, 74, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Choi, P.S.; Bernstein, H.D. Sequential Translocation of an Escherichia coli Two-partner Secretion Pathway Exoprotein Across the Inner and Outer Membranes. Mol. Microbiol. 2010, 75, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Schiebel, E.; Schwarz, H.; Braun, V. Subcellular Location and Unique Secretion of the Hemolysin of Serratia marcescens. J. Biol Chem. 1989, 264, 16311–16320. [Google Scholar] [CrossRef]
- St Geme, J.W., III; Grass, S. Secretion of the Haemophilus influenzae HMW1 and HMW2 Adhesins Involves a Periplasmic Intermediate and Requires the HMWB and HMWC Proteins. Mol. Microbiol. 1998, 27, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Van Ulsen, P.; Rutten, L.; Feller, M.; Tommassen, J.; van der Ende, A. Two-partner Secretion Systems of Neisseria meningitidis Associated with Invasive Clonal Complexes. Infect. Immun. 2008, 76, 4649–4658. [Google Scholar] [CrossRef] [PubMed]
- Hodak, H.; Wohlkönig, A.; Smet-Nocca, C.; Drobecq, H.; Wieruszeski, J.M.; Sénéchal, M.; Landrieu, I.; Locht, C.; Jamin, M.; Jacob-Dubuisson, F. The Peptidyl-prolyl Isomerase and Chaperone Par27 of Bordetella pertussis as the Prototype for a New Group of Parvulins. J. Mol. Biol. 2008, 376, 414–426. [Google Scholar] [CrossRef]
- Fan, E.; Fiedler, S.; Jacob-Dubuisson, F.; Müller, M. Two-partner Secretion of Gram-negative Bacteria: A Single β-barrel Protein Enables Transport Across the Outer Membrane. J. Biol. Chem. 2012, 287, 2591–2599. [Google Scholar] [CrossRef]
- Grosskinsky, U.; Schütz, M.; Fritz, M.; Schmid, Y.; Lamparter, M.C.; Szczesny, P.; Lupas, A.N.; Autenrieth, I.B.; Linke, D. A Conserved Glycine Residue of Trimeric Autotransporter Domains Plays a Key Role in Yersinia Adhesin A Autotransport. J. Bacteriol. 2007, 189, 9011–9019. [Google Scholar] [CrossRef]
- Lehr, U.; Schütz, M.; Oberhettinger, P.; Ruiz-Perez, F.; Donald, J.W.; Palmer, T.; Linke, D.; Henderson, I.R.; Autenrieth, I.B. C-terminal Amino Acid Residues of the Trimeric Autotransporter Adhesin YadA of Yersinia enterocolitica are Decisive for Its Recognition and Assembly by BamA. Mol. Microbiol. 2010, 78, 932–946. [Google Scholar] [CrossRef]
- Ulrich, T.; Oberhettinger, P.; Schütz, M.; Holzer, K.; Ramms, A.S.; Linke, D.; Autenrieth, I.B.; Rapaport, D. Evolutionary Conservation in Biogenesis of β-barrel Proteins Allows Mitochondria to Assemble a Functional Bacterial Trimeric Autotransporter Protein. J. Biol. Chem. 2014, 289, 29457–29470. [Google Scholar] [CrossRef]
- Ishikawa, M.; Yoshimoto, S.; Hayashi, A.; Kanie, J.; Hori, K. Discovery of a Novel Periplasmic Protein That Forms a Complex with a Trimeric Autotransporter Adhesin and Peptidoglycan. Mol. Microbiol. 2016, 101, 394–410. [Google Scholar] [CrossRef]
- Brandon, L.D.; Goldberg, M.B. Periplasmic Transit and Disulfide Bond Formation of the Autotransported Shigella Protein IcsA. J. Bacteriol. 2001, 183, 951–958. [Google Scholar] [CrossRef]
- Buscher, A.Z.; Grass, S.; Heuser, J.; Roth, R.; St Geme, J.W., 3rd. Surface Anchoring of a Bacterial Adhesin Secreted by the Two-partner Secretion Pathway. Mol. Microbiol. 2006, 61, 470–483. [Google Scholar] [CrossRef]
- Paes, W.; Dowle, A.; Coldwell, J.; Leech, A.; Ganderton, T.; Brzozowski, A. The Chlamydia trachomatis PmpD Adhesin Forms Higher Order Structures through Disulphide-mediated Covalent Interactions. PLoS ONE 2018, 13, e0198662. [Google Scholar] [CrossRef]
- Hooda, Y.; Moraes, T.F. Translocation of Lipoproteins to the Surface of Gram-negative Bacteria. Curr. Opin. Struct. Biol. 2018, 51, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Noinaj, N.; Kuszak, A.J.; Gumbart, J.C.; Lukacik, P.; Chang, H.; Easley, N.C.; Lithgow, T.; Buchanan, S.K. Structural Insight into the Biogenesis of β-barrel Membrane Proteins. Nature 2013, 501, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Bakelar, J.; Buchanan, S.K.; Noinaj, N. The Structure of the β-barrel Assembly Machinery Complex. Science 2016, 351, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Li, H.; Dong, H.; Zeng, Y.; Zhang, Z.; Paterson, N.G.; Stansfeld, P.J.; Wang, Z.; Zhang, Y.; Wang, W.; et al. Structural Basis of Outer Membrane Protein Insertion by the BAM Complex. Nature 2016, 531, 64–69. [Google Scholar] [CrossRef]
- Han, L.; Zheng, J.; Wang, Y.; Yang, X.; Liu, Y.; Sun, C.; Cao, B.; Zhou, H.; Ni, D.; Lou, J.; et al. Structure of the BAM Complex and Its Implications for Biogenesis of Outer-membrane Proteins. Nat. Struct Mol. Biol. 2016, 23, 192–196. [Google Scholar] [CrossRef]
- Iadanza, M.G.; Higgins, A.J.; Schiffrin, B.; Calabrese, A.N.; Brockwell, D.J.; Ashcroft, A.E.; Radford, S.E.; Ranson, N.A. Lateral Opening in the Intact β-barrel Assembly Machinery Captured by Cryo-EM. Nat. Commun. 2016, 7, 12865. [Google Scholar] [CrossRef]
- Jain, S.; Goldberg, M.B. Requirement for YaeT in the Outer Membrane Assembly of Autotransporter Proteins. J. Bacteriol. 2007, 189, 5393–5398. [Google Scholar] [CrossRef] [PubMed]
- Rossiter, A.E.; Leyton, D.L.; Tveen-Jensen, K.; Browning, D.F.; Sevastsyanovich, Y.; Knowles, T.J.; Nichols, K.B.; Cunningham, A.F.; Overduin, M.; Schembri, M.A.; et al. The Essential β-barrel Assembly Machinery Complex Components BamD and BamA are Required for Autotransporter Biogenesis. J. Bacteriol. 2011, 193, 4250–4253. [Google Scholar] [CrossRef]
- Rooke, J.L.; Icke, C.; Wells, T.J.; Rossiter, A.E.; Browning, D.F.; Morris, F.C.; Leo, J.C.; Schütz, M.S.; Autenrieth, I.B.; Cunningham, A.F.; et al. BamA and BamD Are Essential for the Secretion of Trimeric Autotransporter Adhesins. Front. Microbiol. 2021, 12, 628879. [Google Scholar] [CrossRef]
- Ranava, D.; Caumont-Sarcos, A.; Albenne, C.; Ieva, R. Bacterial Machineries for the Assembly of Membrane-embedded β-barrel Proteins. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef] [PubMed]
- Tomasek, D.; Rawson, S.; Lee, J.; Wzorek, J.S.; Harrison, S.C.; Li, Z.; Kahne, D. Structure of a Nascent Membrane Protein as it Folds on the BAM Complex. Nature 2020, 583, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Ieva, R.; Skillman, K.M.; Bernstein, H.D. Incorporation of a Polypeptide Segment into the Beta-domain Pore during the Assembly of a Bacterial Autotransporter. Mol. Microbiol. 2008, 67, 188–201. [Google Scholar] [CrossRef] [PubMed]
- Sikdar, R.; Peterson, J.H.; Anderson, D.E.; Bernstein, H.D. Folding of a Bacterial Integral Outer Membrane Protein is Initiated in the Periplasm. Nat. Commun. 2017, 8, 1309. [Google Scholar] [CrossRef]
- Doyle, M.T.; Bernstein, H.D. Bacterial Outer Membrane Proteins Assemble via Asymmetric Interactions with the BamA β-barrel. Nat. Commun. 2019, 10, 3358. [Google Scholar] [CrossRef] [PubMed]
- Bamert, R.S.; Lundquist, K.; Hwang, H.; Webb, C.T.; Shiota, T.; Stubenrauch, C.J.; Belousoff, M.J.; Goode, R.J.A.; Schittenhelm, R.B.; Zimmerman, R.; et al. Structural Basis for Substrate Selection by the Translocation and Assembly Module of the β-barrel Assembly Machinery. Mol. Microbiol. 2017, 106, 142–156. [Google Scholar] [CrossRef] [PubMed]
- Gruss, F.; Zähringer, F.; Jakob, R.P.; Burmann, B.M.; Hiller, S.; Maier, T. The Structural Basis of Autotransporter Translocation by TamA. Nat. Struct. Mol. Biol. 2013, 20, 1318–1320. [Google Scholar] [CrossRef]
- Heinz, E.; Stubenrauch, C.J.; Grinter, R.; Croft, N.P.; Purcell, A.W.; Strugnell, R.A.; Dougan, G.; Lithgow, T. Conserved Features in the Structure, Mechanism, and Biogenesis of the Inverse Autotransporter Protein Family. Genome Biol. Evol. 2016, 8, 1690–1705. [Google Scholar] [CrossRef] [PubMed]
- Saragliadis, A.; Trunk, T.; Leo, J.C. Producing Gene Deletions in Escherichia coli by P1 Transduction with Excisable Antibiotic Resistance Cassettes. J. Vis. Exp. 2018, 139, 58267. [Google Scholar] [CrossRef]
- Norell, D.; Heuck, A.; Tran-Thi, T.A.; Götzke, H.; Jacob-Dubuisson, F.; Clausen, T.; Daley, D.O.; Braun, V.; Müller, M.; Fan, E. Versatile in vitro System to Study Translocation and Functional Integration of Bacterial Outer Membrane Proteins. Nat. Commun. 2014, 5, 5396. [Google Scholar] [CrossRef]
- Bernstein, H.D. Type V Secretion in Gram-negative Bacteria. EcoSal Plus. 2019, 8, 307–318. [Google Scholar]
- Nash, Z.M.; Cotter, P.A. Bordetella Filamentous Hemagglutinin, a Model for the Two-partner Secretion Pathway. EcoSal Plus. 2019, 7, 319–328. [Google Scholar]
- Leo, J.C.; Oberhettinger, P.; Yoshimoto, S.; Udatha, D.B.; Morth, J.P.; Schütz, M.; Hori, K.; Linke, D. Secretion of the Intimin Passenger Domain Is Driven by Protein Folding. J. Biol. Chem. 2016, 291, 20096–20112. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Lopez, M.; Zorgani, M.A.; Kelley, L.A.; Bailly, X.; Kajava, A.V.; Henderson, I.R.; Polticelli, F.; Pizza, M.; Rosini, R.; Desvaux, M. Identification of the Autochaperone Domain in the Type Va Secretion System (T5aSS): Prevalent Feature of Autotransporters with a β-helical Passenger. Front. Microbiol. 2018, 8, 2607. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Johnson, M.D.; Zhang, J.; Lo, A.W.; Schembri, M.A.; Wijeyewickrema, L.C.; Pike, R.N.; Huysmans, G.H.M.; Henderson, I.R.; Leyton, D.L. Molecular Basis for the Folding of β-helical Autotransporter Passenger Domains. Nat. Commun. 2018, 9, 1395. [Google Scholar] [CrossRef]
- Baud, C.; Guérin, J.; Petit, E.; Lesne, E.; Dupré, E.; Locht, C.; Jacob-Dubuisson, F. Translocation Path of a Substrate Protein through Its Omp85 Transporter. Nat. Commun. 2014, 5, 5271. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dautin, N. Folding Control in the Path of Type 5 Secretion. Toxins 2021, 13, 341. https://doi.org/10.3390/toxins13050341
Dautin N. Folding Control in the Path of Type 5 Secretion. Toxins. 2021; 13(5):341. https://doi.org/10.3390/toxins13050341
Chicago/Turabian StyleDautin, Nathalie. 2021. "Folding Control in the Path of Type 5 Secretion" Toxins 13, no. 5: 341. https://doi.org/10.3390/toxins13050341
APA StyleDautin, N. (2021). Folding Control in the Path of Type 5 Secretion. Toxins, 13(5), 341. https://doi.org/10.3390/toxins13050341