Duvernoy’s Gland Transcriptomics of the Plains Black-Headed Snake, Tantilla nigriceps (Squamata, Colubridae): Unearthing the Venom of Small Rear-Fanged Snakes

 , , , and
, , , and
Abstract
1. Introduction
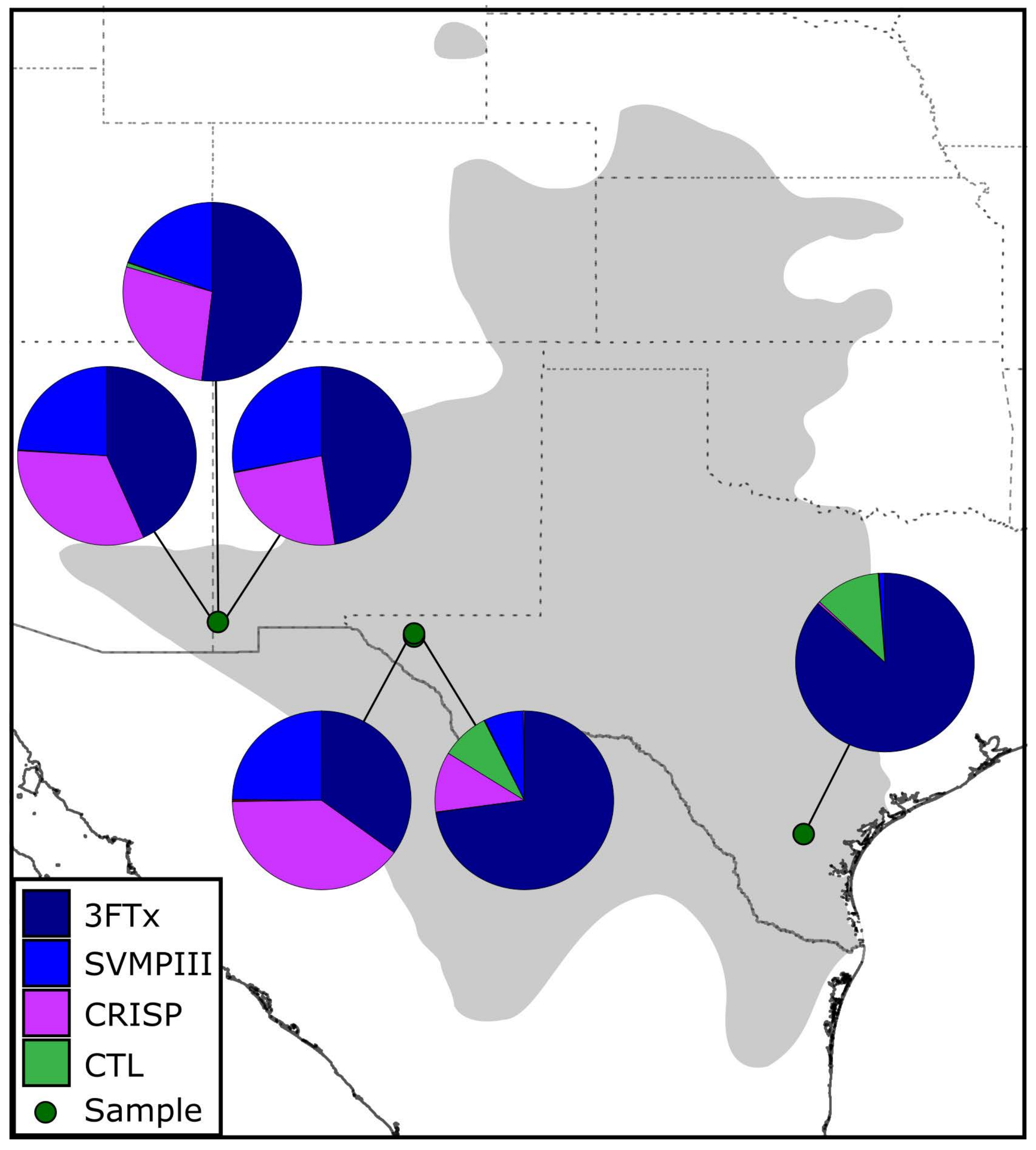
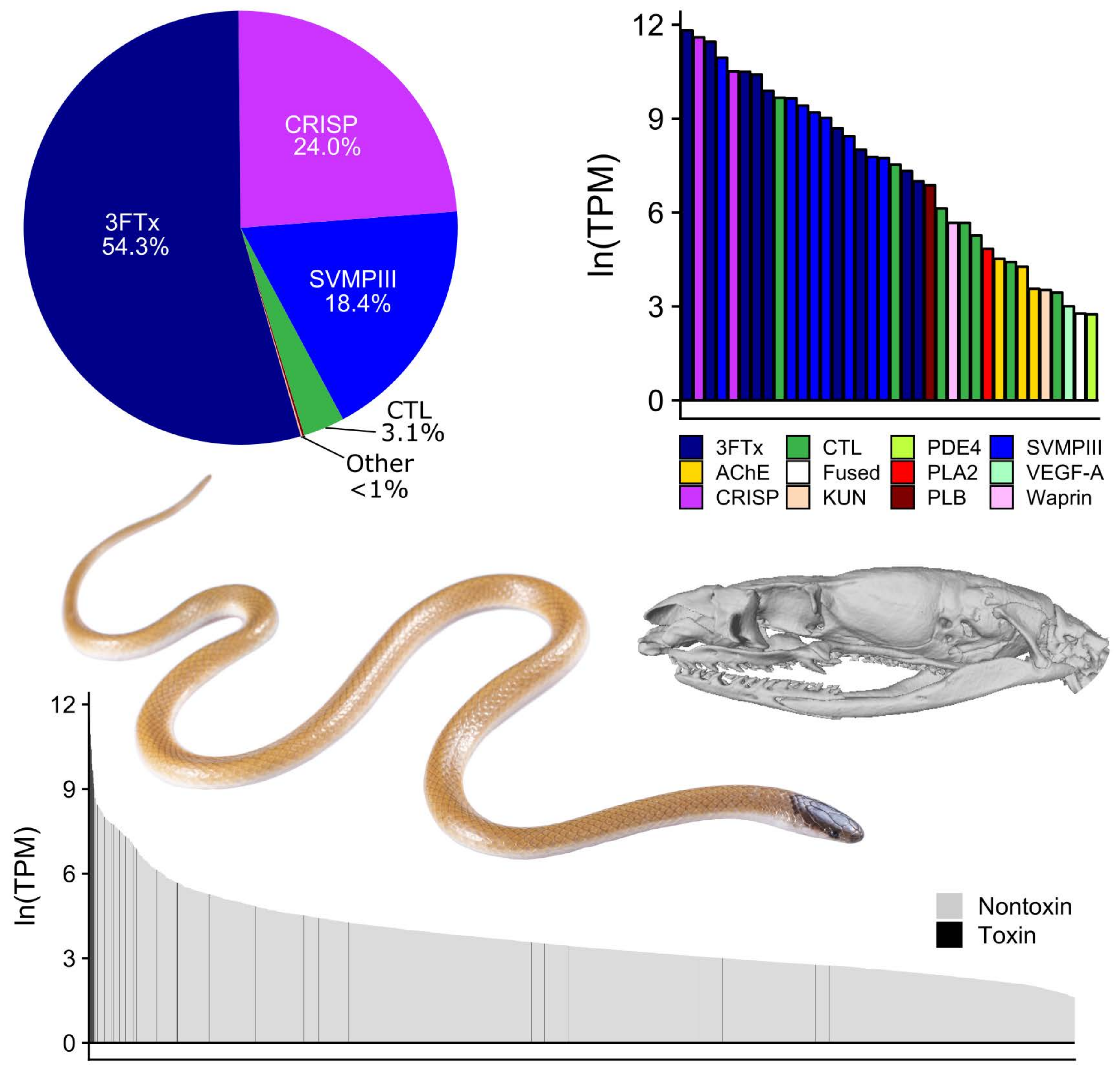
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sample and Gland Collection
5.2. RNA Extraction and Sequencing
5.3. Transcriptome Assembly and Annotation
5.4. Transcriptomic Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Schendel, V.; Rash, L.D.; Jenner, R.A.; Undheim, E.A. The diversity of venom: The importance of behavior and venom system morphology in understanding its ecology and evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef] [PubMed]
- Zancolli, G.; Casewell, N.R. Venom systems as models for studying the origin and regulation of evolutionary novelties. Mol. Biol. Evol. 2020, 37, 2777–2790. [Google Scholar] [CrossRef] [PubMed]
- Kazandjian, T.D.; Petras, D.; Robinson, S.D.; Van Thiel, J.; Greene, H.W.; Arbuckle, K.; Barlow, A.; Carter, D.A.; Wouters, R.M.; Whiteley, G.; et al. Convergent evolution of pain-inducing defensive venom components in spitting cobras. Science 2021, 371, 386–390. [Google Scholar] [CrossRef]
- Deufel, A.; Cundall, D. Functional plasticity of the venom delivery system in snakes with a focus on the poststrike prey release behavior. Zool. Anz. 2006, 245, 249–267. [Google Scholar] [CrossRef]
- Ward-Smith, H.; Arbuckle, K.; Naude, A.; Wüster, W. Fangs for the memories? A survey of pain in snakebite patients does not support a strong role for defense in the evolution of snake venom composition. Toxins 2020, 12, 201. [Google Scholar] [CrossRef]
- Holding, M.L.; Biardi, J.E.; Gibbs, H.L. Coevolution of venom function and venom resistance in a rattlesnake predator and its squirrel prey. Proc. R. Soc. B. Biol. Sci. 2016, 283. [Google Scholar] [CrossRef]
- Holding, M.L.; Margres, M.J.; Rokyta, D.R.; Gibbs, H.L. Local prey community composition and genetic distance predict venom divergence among populations of the Northern Pacific Rattlesnake (Crotalus oreganus). J. Evol. Biol. 2018, 31, 1513–1528. [Google Scholar] [CrossRef]
- Davies, E.L.; Arbuckle, K. Coevolution of snake venom toxic activities and diet: Evidence that ecological generalism favours toxicological diversity. Toxins 2019, 11, 711. [Google Scholar] [CrossRef]
- Healy, K.; Carbone, C.; Jackson, A.L. Snake venom potency and yield are associated with prey-evolution, predator metabolism and habitat structure. Ecol. Lett. 2019, 22, 527–537. [Google Scholar] [CrossRef]
- Lyons, K.; Dugon, M.M.; Healy, K. Diet breadth mediates the prey specificity of venom potency in snakes. Toxins 2020, 12, 74. [Google Scholar] [CrossRef]
- Savitzky, A.H. The role of venom delivery strategies in snake evolution. Evolution 1980, 34, 1194. [Google Scholar] [CrossRef]
- Mason, A.; Margres, M.; Strickland, J.; Rokyta, D.; Sasa, M.; Parkinson, C. Trait differentiation and modular toxin expression in palm-pitvipers. BMC Genom. 2020, 21. [Google Scholar] [CrossRef]
- Barua, A.; Mikheyev, A.S.; Russo, C. Many options, few solutions: Over 60 My snakes converged on a few optimal venom formulations. Mol. Biol. Evol. 2019, 36, 1964–1974. [Google Scholar] [CrossRef]
- Barua, A.; Mikheyev, A.S. Toxin expression in snake venom evolves rapidly with constant shifts in evolutionary rates. Proc. R. Soc. Biol. Sci. 2020, 287. [Google Scholar] [CrossRef]
- Young, B.A.; Lee, C.E.; Daley, K.M. Do snakes meter venom? BioScience 2002, 51, 1121–1126. [Google Scholar] [CrossRef]
- Saviola, A.J.; Chiszar, D.; Busch, C.; Mackessy, S.P. Molecular basis for prey relocation in viperid snakes. BMC Biol. 2013, 11. [Google Scholar] [CrossRef]
- Casewell, N.R.; Jackson, T.N.; Laustsen, A.H.; Sunagar, K. Causes and consequences of snake venom variation. Trends Pharmacol. Sci. 2020, 41, 570–581. [Google Scholar] [CrossRef]
- Kasturiratne, A.; Wickremasinghe, A.R.; De Silva, N.; Kithsiri Gunawardena, N.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; Janaka De Silva, H.; Winkel, K. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Prim. 2017, 3, 1–21. [Google Scholar] [CrossRef]
- Margres, M.J.; Wray, K.P.; Hassinger, A.T.; Ward, M.J.; McGivern, J.J.; Lemmon, E.M.; Lemmon, A.R.; Rokyta, D.R. Quantity, not quality: Rapid adaptation in a polygenic trait proceeded exclusively through expression differentiation. Mol. Biol. Evol. 2017, 34, 3099–3110. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, E.P.; Rautsaw, R.M.; Strickland, J.L.; Holding, M.L.; Hogan, M.P.; Mason, A.J.; Rokyta, D.R.; Parkinson, C.L. Comparative venom-gland transcriptomics and venom proteomics of four Sidewinder Rattlesnake (Crotalus cerastes) lineages reveal little differential expression despite individual variation. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Strickland, J.L.; Smith, C.F.; Mason, A.J.; Schield, D.R.; Borja, M.; Castañeda-Gaytán, G.; Spencer, C.L.; Smith, L.L.; Trápaga, A.; Bouzid, N.M.; et al. Evidence for divergent patterns of local selection driving venom variation in Mojave Rattlesnakes (Crotalus scutulatus). Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Rautsaw, R.M.; Hofmann, E.P.; Margres, M.J.; Holding, M.L.; Strickland, J.L.; Mason, A.J.; Rokyta, D.R.; Parkinson, C.L. Intraspecific sequence and gene expression variation contribute little to venom diversity in Sidewinder Rattlesnakes (Crotalus cerastes). Proc. R. Soc. B. Biol. Sci. 2019, 286. [Google Scholar] [CrossRef]
- Junqueira de Azevedo, I.L.M.; Campos, P.F.; Ching, A.T.C.; Mackessy, S.P. Colubrid venom vomposition: An -Omics perspective. Toxins 2016, 8, 230. [Google Scholar] [CrossRef]
- Modahl, C.M.; Mackessy, S.P. Venoms of rear-fanged snakes: New proteins and novel activities. Front. Ecol. Evol. 2019, 7. [Google Scholar] [CrossRef]
- McKinstry, D.M. Evidence of toxic saliva in some colubrid snakes of the United States. Toxicon 1978, 16, 523–534. [Google Scholar] [CrossRef]
- Rodríguez-Robles, J.A. Are the Duvernoy’s gland secretions of colubrid snakes venoms? J. Herpetol. 1994, 28, 388–390. [Google Scholar] [CrossRef]
- Rodríguez-Robles, J.A.; Thomas, R. Venom function in the Puerto Rican Racer, Alsophis portoricensis (Serpentes: Colubridae). Copeia 1992, 1, 62–68. [Google Scholar] [CrossRef]
- Kardong, K. Colubrid snakes and Duvernoy’s “venom” glands. J. Toxicol. Rev. 2002, 21, 1–19. [Google Scholar] [CrossRef]
- Mackessy, S.P.; Saviola, A.J. Understanding biological roles of venoms among the Caenophidia: The importance of rear-fanged snakes. Integr. Comp. Biol. 2016, 56, 1004–1021. [Google Scholar] [CrossRef]
- Weinstein, S.; Smith, T.; Kardong, K. Reptile Venom Glands: Form, Function, and Future. In Handbook of Reptile Venoms and Toxins; Mackessy, S., Ed.; CRC, Taylor Francis: Boca Raton, FL, USA, 2010; pp. 65–91. [Google Scholar]
- Rosenberg, H.I. An improved method for collecting secretion from Duvernoy’s gland of colubrid snakes. Copeia 1992, 1992, 244–246. [Google Scholar] [CrossRef]
- Mackessy, S.P. Biochemistry and pharmacology of colubrid snake venoms. J. Toxicol. Toxin Rev. 2002, 21, 43–83. [Google Scholar] [CrossRef]
- Fry, B.G.; Wüster, W.; Fadil, S.; Ramjan, R.; Jackson, T.; Martelli, P.; Kini, R.M. Analysis of Colubroidea snake venoms by liquid chromatography with mass spectrometry: Evolutionary and toxinological implications. Rapid Commun. Mass Spectrom. 2003, 17, 2047–2062. [Google Scholar] [CrossRef]
- Modahl, C.M.; Saviola, A.J.; Mackessy, S.P. Venoms of Colubrids. In Venom Genomics and Proteomics; Gopalakrishnakone, P., Calvete, J., Eds.; Springer: Berlin, Germany, 2016; pp. 51–79. [Google Scholar] [CrossRef]
- Jackson, T.N.W.; Jouanne, H.; Vidal, N. Snake venom in context: Neglected clades and concepts. Front. Ecol. Evol. 2019, 7, 332. [Google Scholar] [CrossRef]
- Bayona-Serrano, J.D.; Viala, V.L.; Rautsaw, R.M.; Schramer, T.D.; Barros-Carvalho, G.A.; Nishiyama, M.Y.; Freitas-de-Sousa, A.L.; Moura-da Silva, A.M.; Parkinson, C.L.; Grazziotin, F.G.; et al. Replacement and parallel simplification of nonhomologous proteinases maintain venom phenotypes in rear-fanged snakes. Mol. Biol. Evol. 2020, 37, 3563–3575. [Google Scholar] [CrossRef]
- Heyborne, W.H.; Mackessy, S.P. Identification and characterization of a taxon-specific three-finger toxin from the venom of the Green Vinesnake (Oxybelis fulgidus; Family Colubridae). Biochimie 2013, 95, 1923–1932. [Google Scholar] [CrossRef]
- Modahl, C.M.; Mrinalini; Frietze, S.; Mackessy, S.P. Adaptive evolution of distinct prey-specific toxin genes in rear-fanged snake venom. Proc. R. Soc. B Biol. Sci. 2018, 285. [Google Scholar] [CrossRef]
- Fry, B.G.; Scheib, H.; de Azevedo, I.d.L.J.; Silva, D.A.; Casewell, N.R. Novel transcripts in the maxillary venom glands of advanced snakes. Toxicon 2012, 59, 696–708. [Google Scholar] [CrossRef]
- Von Reumont, B.M. Studying smaller and neglected organisms in modern evolutionary venomics implementing RNASeq (Transcriptomics)—A critical guide. Toxins 2018, 10, 292. [Google Scholar] [CrossRef]
- Haney, R.A.; Clarke, T.H.; Gadgil, R.; Fitzpatrick, R.; Hayashi, C.Y.; Ayoub, N.A.; Garb, J.E. Effects of gene duplication, positive selection, and shifts in gene expression on the evolution of the venom gland transcriptome in widow spiders. Genome Biol. Evol. 2016, 8, 228–242. [Google Scholar] [CrossRef]
- Drukewitz, S.H.; Fuhrmann, N.; Undheim, E.A.; Blanke, A.; Giribaldi, J.; Mary, R.; Laconde, G.; Dutertre, S.; von Reumont, B.M. A dipteran’s novel sucker punch: Evolution of arthropod atypical venom with a neurotoxic component in robber flies (Asilidae, Diptera). Toxins 2018, 10, 29. [Google Scholar] [CrossRef]
- Ellsworth, S.A.; Nystrom, G.S.; Ward, M.J.; Freitas de Sousa, L.A.; Hogan, M.P.; Rokyta, D.R. Convergent recruitment of adamalysin-like metalloproteases in the venom of the Red Bark Centipede (Scolopocryptops sexspinosus). Toxicon 2019, 168, 1–15. [Google Scholar] [CrossRef]
- Modahl, C.M.; Brahma, R.K.; Koh, C.Y.; Shioi, N.; Kini, R.M. Omics technologies for profiling toxin diversity and evolution in snake venom: Impacts on the discovery of therapeutic and diagnostic agents. Annu. Rev. Anim. Biosci. 2020, 8, 91–116. [Google Scholar] [CrossRef]
- Calvete, J.J.; Bonilla, F.; Granados-Martínez, S.; Sanz, L.; Lomonte, B.; Sasa, M. Venomics of the Duvernoy’s gland secretion of the False Coral Snake Rhinobothryum bovallii (Andersson, 1916) and assessment of venom lethality towards synapsid and diapsid animal models. J. Proteom. 2020, 225. [Google Scholar] [CrossRef]
- Uetz, P.; Freed, P.; Hošek, J. The Reptile Database. Available online: http://www.reptile-database.org/ (accessed on 1 May 2020).
- Anderson, P. The Reptiles of Missouri; University of Missouri Press: Colombia, MO, USA, 1965; pp. 1–319. [Google Scholar]
- Trauth, S. Posterior maxillary fangs of the Flathead Snake, Tantilla gracilis (Serpentes: Colubridae), using scanning electron microscopy. J. Ark. Acad. Sci. 1991, 45, 133–136. [Google Scholar]
- Greene, H. Snakes: The Evolution of Mystery in Nature; University of California Press: Berkeley, CA, USA, 1997. [Google Scholar]
- Hill, R.E.; Mackessy, S.P. Venom yields from several species of colubrid snakes and differential effects of ketamine. Toxicon 1997, 35, 671–678. [Google Scholar] [CrossRef]
- Hill, R.E.; Mackessy, S.P. Characterization of venom (Duvernoy’s secretion) from twelve species of colubrid snakes and partial sequence of four venom proteins. Toxicon 2000, 38, 1663–1687. [Google Scholar] [CrossRef]
- Wilson, L.D.; Mata-Silva, V. A checklist and key to the snakes of the Tantilla clade (Squamata: Colubridae), with comments on taxonomy, distribution, and conservation. Mesoamerican Herpetol. 2015, 2, 418–498. [Google Scholar]
- Antúnez-Fonseca, C.A.; Castro, J.A.; España, F.G.; Townsend, J.H.; Wilson, L.D. A new species of Tantilla of the taeniata group (Squamata: Colubridae) from Refugio de Vida Silvestre Barras de Cuero y Salado in Caribbean coastal Honduras. Amphib. Reptile Conserv. 2020, 14, 86–102. [Google Scholar]
- Ernst, C.; Ernst, E. Snakes of the United States and Canada; Smithsonian Books: Washington, DC, USA, 2003. [Google Scholar]
- Holm, P. Phylogenetic Biology of the Burrowing Snake Tribe Sonorini (Colubridae). Ph.D. Thesis, University of Arizona, Tucson, AZ, USA, 2008. [Google Scholar]
- Todd, B.D.; Willson, J.D.; Winne, C.T.; Semlitsch, R.D.; Gibbons, J.W. Ecology of the Southeastern Crowned Snake, Tantilla coronata. Copeia 2008, 2, 388–394. [Google Scholar] [CrossRef]
- Farrell, T.M.; Smiley-Walters, S.A.; McColl, D.E. Prey species influences foraging behaviors: Rattlesnake (Sistrurus miliarius) predation on Little Brown Skinks (Scincella lateralis) and Giant Centipedes (Scolopendra viridis). J. Herpetol. 2018, 52, 156–161. [Google Scholar] [CrossRef]
- Hamanaka, K.; Mori, A. Toxicity of venom from the Mamushi, Gloydius blomhoffii, (Squamata, Crotalinae) to centipedes. Toxicon 2020, 188, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, W.; Painter, C.; Price, A. Amphibians and Reptiles of New Mexico; University of New Mexico Press: Albuquerque, NM, USA, 1996. [Google Scholar]
- Parga, V. Arthropod Diets in Chihuahuan Desert Snakes. Master’s Thesis, University of Texas, El Paso, TX, USA, 2018. [Google Scholar]
- Saviola, A.J.; Peichoto, M.E.; Mackessy, S.P. Rear-fanged snake venoms: An untapped source of novel compounds and potential drug leads. Toxin Rev. 2014, 33, 185–201. [Google Scholar] [CrossRef]
- Boyer, D.M.; Gunnell, G.F.; Kaufman, S.; McGeary, T.M. MorphoSource: Archiving and sharing 3-D digital specimen data. Paleontol. Soc. Pap. 2016, 22, 157–181. [Google Scholar] [CrossRef]
- Hargreaves, A.D.; Swain, M.T.; Hegarty, M.J.; Logan, D.W.; Mulley, J.F. Restriction and recruitment-gene duplication and the origin and evolution of snake venom toxins. Genome Biol. Evol. 2014, 6, 2088–2095. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef]
- Mcgivern, J.J.; Wray, K.P.; Margres, M.J.; Couch, M.E.; Mackessy, S.P.; Rokyta, D.R. RNA-seq and high-definition mass spectrometry reveal the complex and divergent venoms of two rear-fanged colubrid snakes. BMC Genom. 2014, 15, 1061. [Google Scholar] [CrossRef]
- Modahl, C.M.; Frietze, S.; Mackessy, S.P. Transcriptome-facilitated proteomic characterization of rear-fanged snake venoms reveal abundant metalloproteinases with enhanced activity. J. Proteom. 2018, 187, 223–234. [Google Scholar] [CrossRef]
- de Oliveira, F.A.; de França, R.C.; França, F.G.R. Geographical ecology of Tantilla melanocephala (Squamata: Serpentes: Colubridae) in a Neotropical region: A comparison of northeastern Atlantic Forest and Caatinga populations. Stud. Neotrop. Fauna Environ. 2020, 1–10. [Google Scholar] [CrossRef]
- Cobb, V.A. Diet and prey size of the Flathead Snake, Tantilla gracilis. Copeia 2004, 2, 397–402. [Google Scholar] [CrossRef]
- Conroy, C.; Papenfuss, T.; Parker, J.; Hahn, N. Use of Tricaine Methanesulfonate (MS222) for euthanasia of reptiles. J. Am. Assoc. Lab. Anim. Sci. 2009, 48, 28–32. [Google Scholar]
- Rokyta, D.R.; Lemmon, A.R.; Margres, M.J.; Aronow, K. The venom-gland transcriptome of the Eastern Diamondback Rattlesnake (Crotalus adamanteus). BMC Genom. 2012, 16, 312. [Google Scholar] [CrossRef]
- Rokyta, D.R.; Wray, K.P.; McGivern, J.J.; Margres, M.J. The transcriptomic and proteomic basis for the evolution of a novel venom phenotype within the Timber Rattlesnake (Crotalus horridus). Toxicon 2015, 98, 34–48. [Google Scholar] [CrossRef]
- Rokyta, D.R.; Margres, M.J.; Ward, M.J.; Sanchez, E.E. The genetics of venom ontogeny in the Eastern Diamondback Rattlesnake (Crotalus adamanteus). PeerJ 2017, 5, e3249. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef]
- Holding, M.L.; Margres, M.J.; Mason, A.J.; Parkinson, C.L.; Rokyta, D.R. Evaluating the performance of de novo assembly methods for venom-gland transcriptomics. Toxins 2018, 10, 249. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Stothard, P. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. BioTechniques 2000, 28. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Palarea-Albaladejo, J.; Martín-Fernández, J.A. ZCompositions—R package for multivariate imputation of left-censored data under a compositional approach. Chemom. Intell. Lab. Syst. 2015, 143, 85–96. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| ASNHC No. | Field ID | Locality | Sex | Read Pairs | Merged Reads |
|---|---|---|---|---|---|
| 15178 | CLP2590 | Hudspeth Co., TX, USA | F | 27,832,797 | 22,093,170 |
| 15179 | CLP2591 | Hudspeth Co., TX, USA | M | 28,872,668 | 24,078,211 |
| 15180 | CLP2592 | Duval Co., TX, USA | F | 22,736,233 | 17,826,005 |
| 15181 | CLP2753 | Hidalgo Co., NM, USA | F | 14,445,533 | 10,648,351 |
| 15182 | CLP2754 | Hidalgo Co., NM, USA | M | 8,088,121 | 6,232,048 |
| 15183 | CLP2759 | Hidalgo Co., NM, USA | M | 18,480,258 | 15,030,748 |
| ASNHC | ASNHC | ASNHC | ASNHC | ASNHC | ASNHC | Avg. | |
|---|---|---|---|---|---|---|---|
| 15178 | 15179 | 15180 | 15181 | 15182 | 15183 | ||
| 3FTx | 34.9% | 72.8% | 86.4% | 51.9% | 47.6% | 43.3% | 54.3% |
| CRISP | 39.8% | 10.9% | 0.4% | 27.6% | 24.3% | 32.7% | 24.0% |
| SVMPIII | 24.8% | 7.1% | 1.0% | 19.6% | 27.9% | 23.9% | 18.4% |
| CTL | <0.1% | 8.7% | 11.9% | 0.8% | <0.1% | <0.1% | 3.1% |
| Others | 0.4% | 0.43% | 0.3% | 0.1% | 0.2% | 0.1% | 0.2% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hofmann, E.P.; Rautsaw, R.M.; Mason, A.J.; Strickland, J.L.; Parkinson, C.L. Duvernoy’s Gland Transcriptomics of the Plains Black-Headed Snake, Tantilla nigriceps (Squamata, Colubridae): Unearthing the Venom of Small Rear-Fanged Snakes. Toxins 2021, 13, 336. https://doi.org/10.3390/toxins13050336
Hofmann EP, Rautsaw RM, Mason AJ, Strickland JL, Parkinson CL. Duvernoy’s Gland Transcriptomics of the Plains Black-Headed Snake, Tantilla nigriceps (Squamata, Colubridae): Unearthing the Venom of Small Rear-Fanged Snakes. Toxins. 2021; 13(5):336. https://doi.org/10.3390/toxins13050336
Chicago/Turabian StyleHofmann, Erich P., Rhett M. Rautsaw, Andrew J. Mason, Jason L. Strickland, and Christopher L. Parkinson. 2021. "Duvernoy’s Gland Transcriptomics of the Plains Black-Headed Snake, Tantilla nigriceps (Squamata, Colubridae): Unearthing the Venom of Small Rear-Fanged Snakes" Toxins 13, no. 5: 336. https://doi.org/10.3390/toxins13050336
APA StyleHofmann, E. P., Rautsaw, R. M., Mason, A. J., Strickland, J. L., & Parkinson, C. L. (2021). Duvernoy’s Gland Transcriptomics of the Plains Black-Headed Snake, Tantilla nigriceps (Squamata, Colubridae): Unearthing the Venom of Small Rear-Fanged Snakes. Toxins, 13(5), 336. https://doi.org/10.3390/toxins13050336