Effect of Lonomia obliqua Venom on Human Neutrophils

 , , , , and
, , , , and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
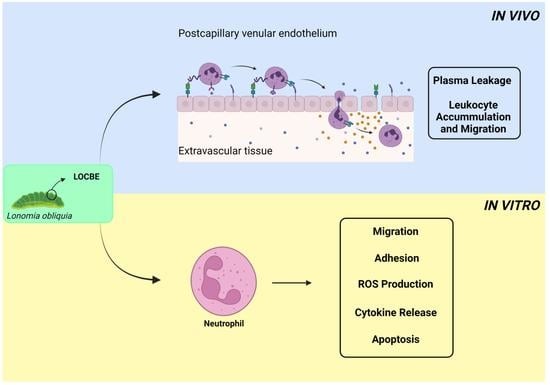
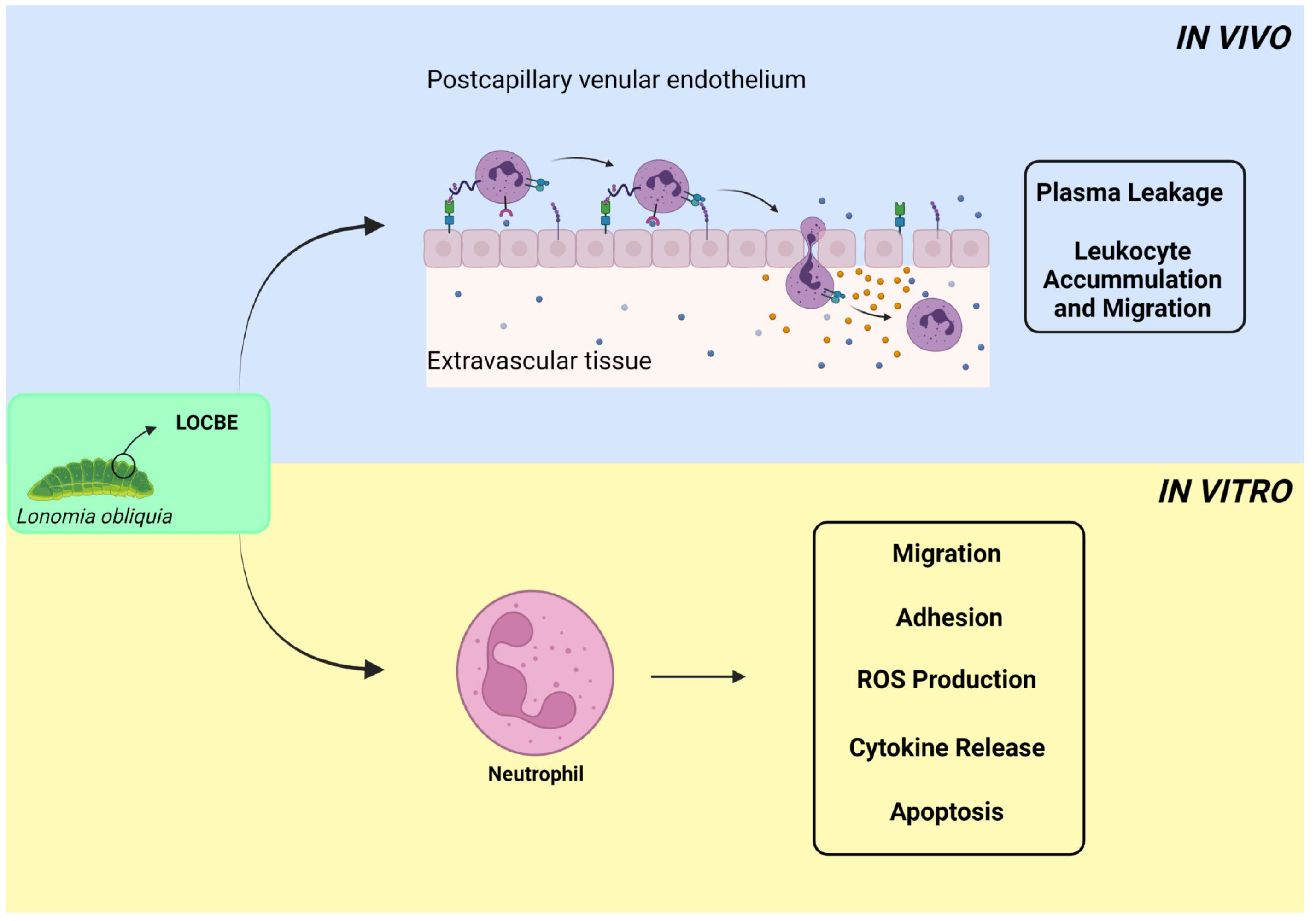
2. Results
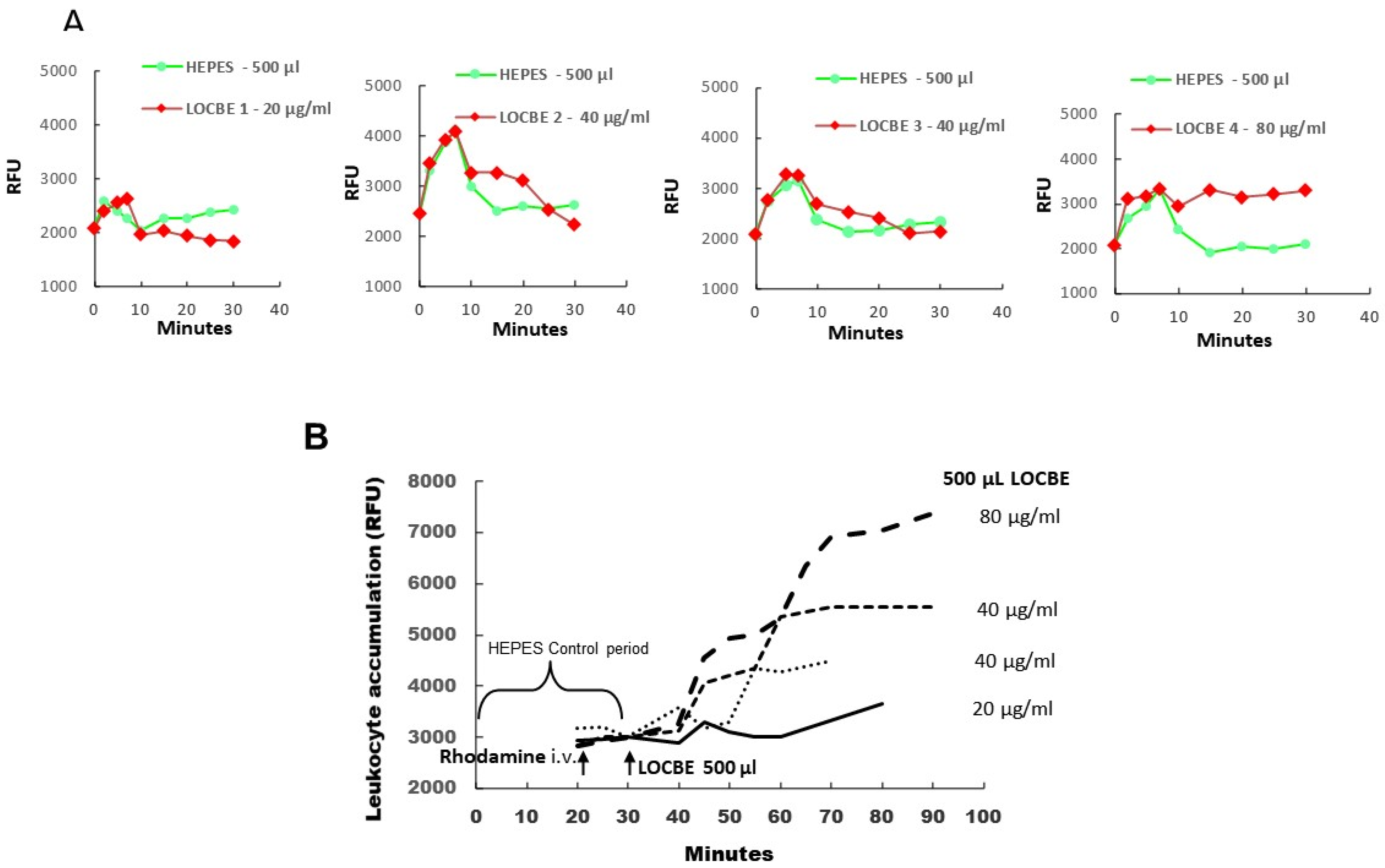
2.1. In Vivo Effect of LOCBE on Plasma Leakage and Leukocyte Accumulation
2.2. Effect of LOCBE on PMN Migration
2.3. Effect of LOCBE on PMN Adhesion
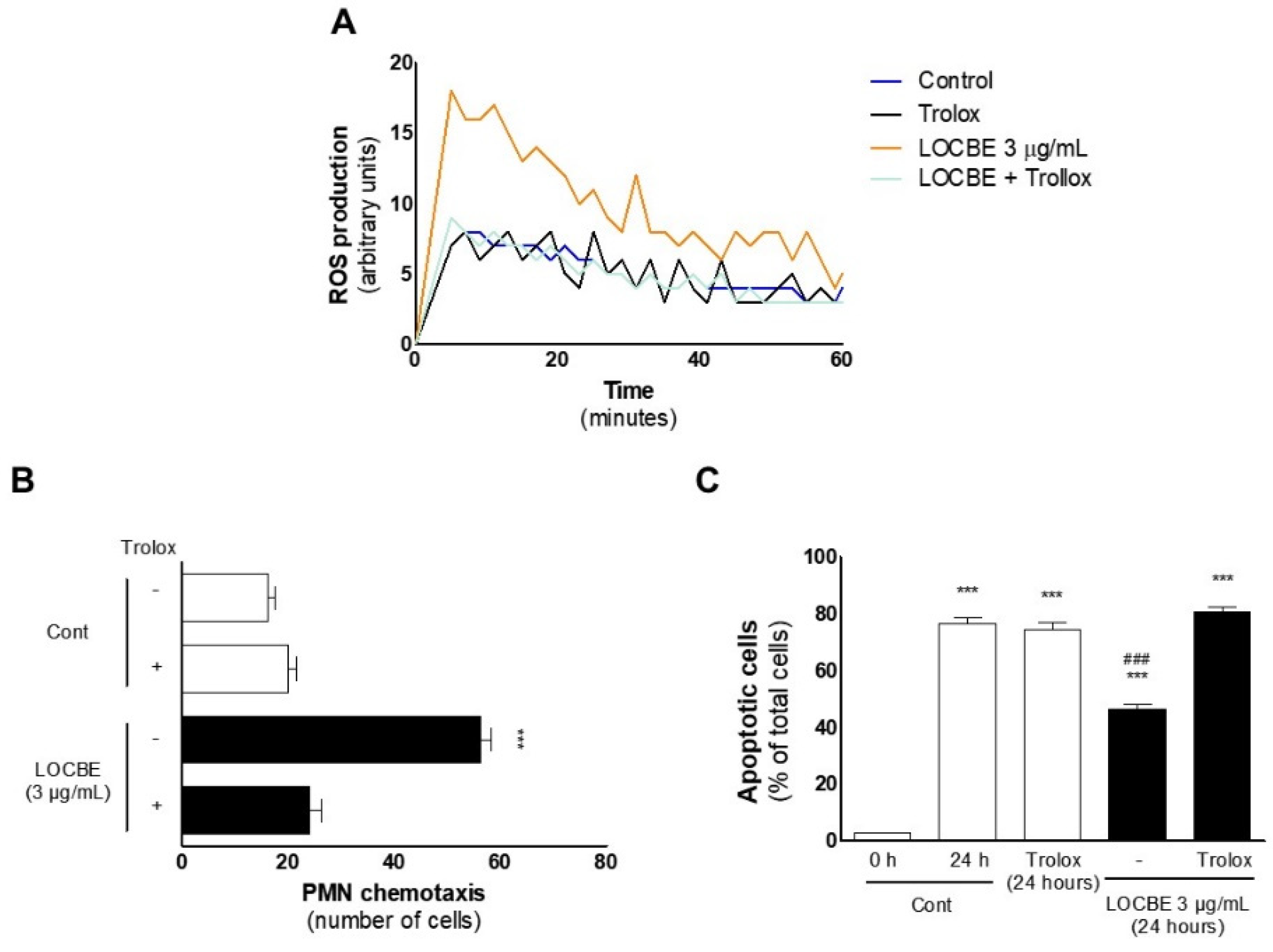
2.4. Effect of LOCBE on PMN ROS Production
2.5. Effect of LOCBE on PMN Survival
2.6. Oxidative Effect of LOCBE on PMN
3. Discussion
4. Conclusions
5. Material and Methods
5.1. Reagents
5.2. Lonomia obliqua Caterpillar Bristle Extract (LOCBE)
5.3. Intravital Microscopy
5.4. PMN Isolation
5.5. Cell Migration Assay
5.6. Cytochemistry
5.7. Western Blot Analysis
5.8. Cell Adhesion Assay
5.9. ROS Production Assay
5.10. ELISA
5.11. Apoptosis Analysis
5.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arocha-Pinango, C.L.; Guerrero, B. Lonomia genus caterpillar envenomation: Clinical and biological aspects. Haemostasis 2001, 31, 288–293. [Google Scholar] [CrossRef]
- Arocha-Pinango, C.L.; Layrisse, M. Fibrinolysis produced by contact with a caterpillar. Lancet 1969, 1, 810–812. [Google Scholar] [CrossRef]
- Carrijo-Carvalho, L.C.; Chudzinski-Tavassi, A.M. The venom of the Lonomia caterpillar: An overview. Toxicon 2007, 49, 741–757. [Google Scholar] [CrossRef]
- Arocha-Pinango, C.L.; Guerrero, B. Hemorrhagic syndrome induced by caterpillars. Clinical and experimental studies. Rev. Investig. Clin. 2003, 44, 155–163. [Google Scholar]
- Maggi, S.; Faulhaber, G.A. Lonomia obliqua Walker (Lepidoptera: Saturniidae): Hemostasis implications. Rev. Assoc. Med. Bras. 2015, 61, 263–268. [Google Scholar] [CrossRef]
- Pinto, A.F.; Dragulev, B.; Guimaraes, J.A.; Fox, J.W. Novel perspectives on the pathogenesis of Lonomia obliqua caterpillar envenomation based on assessment of host response by gene expression analysis. Toxicon 2008, 51, 1119–1128. [Google Scholar] [CrossRef]
- Favalesso, M.M.; Lorini, L.M.; Peichoto, M.E.; Guimaraes, A.T.B. Potential distribution and ecological conditions of Lonomia obliqua Walker 1855 (Saturniidae: Hemileucinae) in Brazil. Acta Trop. 2019, 192, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.; Reck, J., Jr.; Terra, R.M.; Beys da Silva, W.O.; Santi, L.; Pinto, A.F.; Vainstein, M.H.; Termignoni, C.; Guimaraes, J.A. Lonomia obliqua venomous secretion induces human platelet adhesion and aggregation. J. Thromb. Thrombolysis 2010, 30, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Kelen, E.M.A.; Picarelli, Z.P.; Duarte, A.C. Hemorrhagic syndrome induced by contact with caterpillars of the genus Lonomia (Saturniidae, Hemileucinae). J. Toxicol. Toxin Rev. 1995, 14, 283–308. [Google Scholar] [CrossRef]
- Veiga, A.B.; Pinto, A.F.; Guimaraes, J.A. Fibrinogenolytic and procoagulant activities in the hemorrhagic syndrome caused by Lonomia obliqua caterpillars. Thromb. Res. 2003, 111, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.; de Moraes, J.A.; Beys-da-Silva, W.O.; Santi, L.; Terraciano, P.B.; Driemeier, D.; Cirne-Lima, E.O.; Passos, E.P.; Vieira, M.A.R.; Barja-Fidalgo, T.C.; et al. Renal and vascular effects of kallikrein inhibition in a model of Lonomia obliqua venom-induced acute kidney injury. PLoS Negl. Trop. Dis. 2019, 13, e0007197. [Google Scholar] [CrossRef]
- Moraes, J.A.; Rodrigues, G.; Nascimento-Silva, V.; Renovato-Martins, M.; Berger, M.; Guimaraes, J.A.; Barja-Fidalgo, C. Effects of Lonomia obliqua venom on vascular smooth muscle cells: Contribution of NADPH oxidase-derived reactive oxygen species. Toxins 2017, 9, 360. [Google Scholar] [CrossRef]
- Nascimento-Silva, V.; Rodrigues da Silva, G.; Moraes, J.A.; Cyrino, F.Z.; Seabra, S.H.; Bouskela, E.; Almeida Guimaraes, J.; Barja-Fidalgo, C. A pro-inflammatory profile of endothelial cell in Lonomia obliqua envenomation. Toxicon 2012, 60, 50–60. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Colome, A.M.; Lee-Rivera, I.; Benavides-Hidalgo, R.; Lopez, E. Paxillin: A crossroad in pathological cell migration. J. Hematol. Oncol. 2017, 10, 50. [Google Scholar] [CrossRef]
- Mitra, S.K.; Hanson, D.A.; Schlaepfer, D.D. Focal adhesion kinase: In command and control of cell motility. Nat. Rev. Mol. Cell Biol. 2005, 6, 56–68. [Google Scholar] [CrossRef]
- Zeng, M.Y.; Miralda, I.; Armstrong, C.L.; Uriarte, S.M.; Bagaitkar, J. The roles of NADPH oxidase in modulating neutrophil effector responses. Mol. Oral Microbiol. 2019, 34, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Lou, J.; Mao, Y.Y.; Lai, T.W.; Liu, L.Y.; Zhu, C.; Zhang, C.; Liu, J.; Li, Y.Y.; Zhang, F.; et al. Activation of MTOR in pulmonary epithelium promotes LPS-induced acute lung injury. Autophagy 2016, 12, 2286–2299. [Google Scholar] [CrossRef]
- Lai, J.L.; Liu, Y.H.; Liu, C.; Qi, M.P.; Liu, R.N.; Zhu, X.F.; Zhou, Q.G.; Chen, Y.Y.; Guo, A.Z.; Hu, C.M. Indirubin inhibits LPS-induced inflammation via TLR4 abrogation mediated by the NF-kB and MAPK signaling pathways. Inflammation 2017, 40, 1–12. [Google Scholar] [CrossRef]
- Rane, M.J.; Klein, J.B. Regulation of neutrophil apoptosis by modulation of PKB/Akt activation. Front. Biosci. 2009, 14, 2400–2412. [Google Scholar] [CrossRef] [PubMed]
- Ricci-Silva, M.E.; Valente, R.H.; León, I.R.; Tambourgi, D.V.; Ramos, O.H.; Perales, J.; Chudzinski-Tavassi, A.M. Immunochemical and proteomic technologies as tools for unravelling toxins involved in envenoming by accidental contact with Lonomia obliqua caterpillars. Toxicon 2008, 51, 1017–1028. [Google Scholar] [CrossRef]
- Bosch, R.V.; Alvarez-Flores, M.P.; Maria, D.A.; Chudzinski-Tavassi, A.M. Hemolin triggers cell survival on fibroblasts in response to serum deprivation by inhibition of apoptosis. Biomed. Pharmacother. 2016, 82, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Waismam, K.; Chudzinski-Tavassi, A.M.; Carrijo-Carvalho, L.C.; Pacheco, M.T.; Farsky, S.H. Lopap: A non-inflammatory and cytoprotective molecule in neutrophils and endothelial cells. Toxicon 2009, 53, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Svitkina, T. The actin cytoskeleton and actin-based motility. Cold Spring Harb. Perspect. Biol. 2018, 10, a018267. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive oxygen species in metabolic and inflammatory signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef] [PubMed]
- da Silva, G.H.; Hyslop, S.; Alice da Cruz-Hofling, M. Lonomia obliqua caterpillar venom increases permeability of the blood-brain barrier in rats. Toxicon 2004, 44, 625–634. [Google Scholar] [CrossRef]
- Bohrer, C.B.; Reck Junior, J.; Fernandes, D.; Sordi, R.; Guimaraes, J.A.; Assreuy, J.; Termignoni, C. Kallikrein-kinin system activation by Lonomia obliqua caterpillar bristles: Involvement in edema and hypotension responses to envenomation. Toxicon 2007, 49, 663–669. [Google Scholar] [CrossRef]
- Svensjo, E.; Saraiva, E.M.; Amendola, R.S.; Barja-Fidalgo, C.; Bozza, M.T.; Lerner, E.A.; Teixeira, M.M.; Scharfstein, J. Maxadilan, the Lutzomyia longipalpis vasodilator, drives plasma leakage via PAC1-CXCR1/2-pathway. Microvasc. Res. 2012, 83, 185–193. [Google Scholar] [CrossRef]
- Svensjo, E.; Saraiva, E.M.; Bozza, M.T.; Oliveira, S.M.; Lerner, E.A.; Scharfstein, J. Salivary gland homogenates of Lutzomyia longipalpis and its vasodilatory peptide maxadilan cause plasma leakage via PAC1 receptor activation. J. Vasc. Res. 2009, 46, 435–446. [Google Scholar] [CrossRef]
- Monteiro, A.C.; Scovino, A.; Raposo, S.; Gaze, V.M.; Cruz, C.; Svensjo, E.; Narciso, M.S.; Colombo, A.P.; Pesquero, J.B.; Feres-Filho, E.; et al. Kinin danger signals proteolytically released by gingipain induce Fimbriae-specific IFN-gamma- and IL-17-producing T cells in mice infected intramucosally with Porphyromonas gingivalis. J. Immunol. 2009, 183, 3700–3711. [Google Scholar] [CrossRef]
- Moraes, J.A.; Frony, A.C.; Barcellos-de-Souza, P.; Menezes da Cunha, M.; Brasil Barbosa Calcia, T.; Benjamim, C.F.; Boisson-Vidal, C.; Barja-Fidalgo, C. Downregulation of microparticle release and pro-inflammatory properties of activated human polymorphonuclear neutrophils by LMW fucoidan. J. Innate Immun. 2019, 11, 330–346. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moraes, J.A.; Rodrigues, G.; Guimarães-Bastos, D.; Nascimento-Silva, V.; Svensjö, E.; Renovato-Martins, M.; Berger, M.; Guimarães, J.; Barja-Fidalgo, C. Effect of Lonomia obliqua Venom on Human Neutrophils. Toxins 2021, 13, 908. https://doi.org/10.3390/toxins13120908
Moraes JA, Rodrigues G, Guimarães-Bastos D, Nascimento-Silva V, Svensjö E, Renovato-Martins M, Berger M, Guimarães J, Barja-Fidalgo C. Effect of Lonomia obliqua Venom on Human Neutrophils. Toxins. 2021; 13(12):908. https://doi.org/10.3390/toxins13120908
Chicago/Turabian StyleMoraes, João Alfredo, Genilson Rodrigues, Daniel Guimarães-Bastos, Vany Nascimento-Silva, Erik Svensjö, Mariana Renovato-Martins, Markus Berger, Jorge Guimarães, and Christina Barja-Fidalgo. 2021. "Effect of Lonomia obliqua Venom on Human Neutrophils" Toxins 13, no. 12: 908. https://doi.org/10.3390/toxins13120908
APA StyleMoraes, J. A., Rodrigues, G., Guimarães-Bastos, D., Nascimento-Silva, V., Svensjö, E., Renovato-Martins, M., Berger, M., Guimarães, J., & Barja-Fidalgo, C. (2021). Effect of Lonomia obliqua Venom on Human Neutrophils. Toxins, 13(12), 908. https://doi.org/10.3390/toxins13120908