Reduced Toxicity of Centruroides vittatus (Say, 1821) May Result from Lowered Sodium β Toxin Gene Expression and Toxin Protein Production

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Sodium β Toxin Variability Analysis
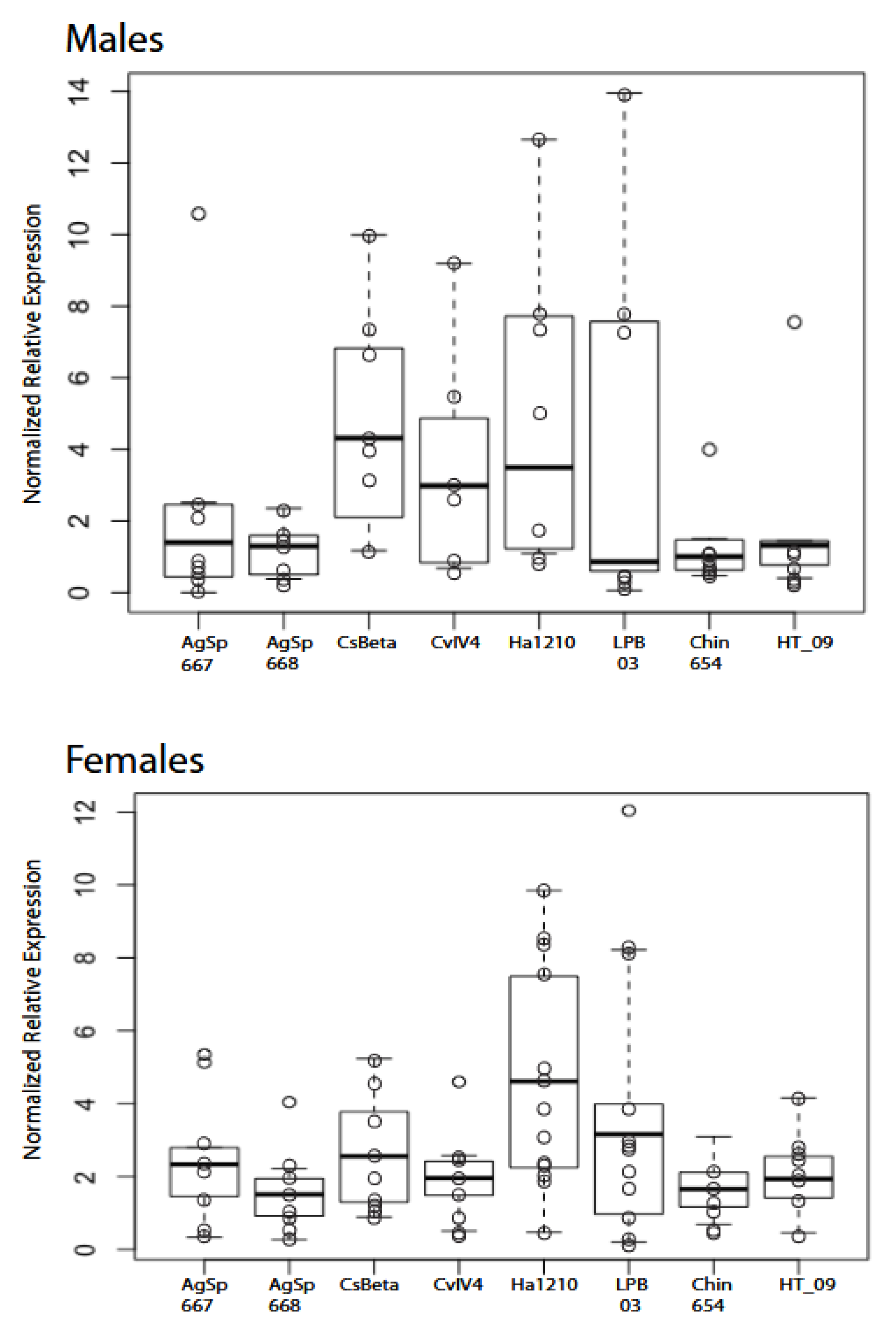
2.2. RT-qPCR Experiments
2.3. Transcriptome
2.4. Toxin Proteomics
3. Discussion
3.1. Sodium β Toxin Gene Diversity
3.2. Sodium β Toxin Gene Expression Variation
3.3. Sodium β Toxin Transcriptome and Proteome
4. Conclusions
5. Materials and Methods
5.1. Population Sodium β Toxin Gene Variability Analysis
5.2. Sodium β Toxin Gene Expression (RT-qPCR)
5.3. Transcriptome
5.4. Toxin Proteomics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gantenbein, B.; Fet, V.; Barker, M.D. Mitochondrial DNA reveals a deep, divergent phylogeny in Centruroides exilicauda (Wood, 1863) (Scorpiones: Buthidae). In Scorpions 2001. In Memoriam Gary A. Polis; Fet, V., Seldon, P.A., Eds.; British Arachnology Society: Burnham Beeches, Bucks, UK, 2001; pp. 235–244. [Google Scholar]
- Santibáñez-López, C.E.; Francke, O.F.; Ureta, C.; Possani, L.D. Scorpions from Mexico: From species diversity to venom complexity. Toxins 2016, 8, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sissom, W.D. Life history. In The Biology of the Scorpions; Polis, G.A., Ed.; Stanford University Press: Stanford, CA, USA, 1990; pp. 161–223. [Google Scholar]
- Teruel, R.; Fet, V.; Graham, M.R. The first mitochondrial DNA phylogeny of Cuban Buthidae (Scorpiones: Buthoidea). Boletín Soc. Entomológica Aragonesa 2006, 39, 219–226. [Google Scholar]
- Borges, A.; Miranda, R.J.; Pascale, J.M. Scorpionism in Central America, with special reference to the case of Panama. J. Ven. Ani. Tox. Trop. Dis. 2012, 18, 130–143. [Google Scholar] [CrossRef] [Green Version]
- Carcamo-Noriega, E.N.; Olamendi-Portugal, T.; Restano-Cassulini, R.; Rowe, A.; Uribe-Romero, S.J.; Becerril, B.; Possani, L.D. Intraspecific variation of Centruroides sculpturatus scorpion venom from two Regions of Arizona. Arch. Biochem. Biophys. 2018, 638, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Ward, M.J.; Ellsworth, S.A.; Hogan, M.P.; Nystrom, G.S.; Martinez, P.; Budhdeo, A.; Zelaya, R.; Perez, A.; Powell, B.; He, H.; et al. Female-biased population divergence in the venom of the Hentz striped scorpion (Centruroides hentzi). Toxicon 2018, 152, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.W.; Jones, A.D.; Goldston, J.S.; Rowe, M.P.; Rowe, A.H. Sex differences in defensive behavior and venom of the striped bark scorpion Centruroides vittatus (Scorpiones: Buthidae). Integ. Comp. Biol. 2016, 56, 1022–1031. [Google Scholar] [CrossRef] [Green Version]
- McElroy, T.; McReynolds, C.N.; Gulledge, A.; Knight, K.R.; Smith, W.E.; Albrecht, E.A. Differential toxicity and venom gland gene expression in Centruroides vittatus. PLoS ONE 2017, 12, e0184695. [Google Scholar] [CrossRef] [Green Version]
- Ward, M.J.; Ellsworth, S.A.; Rokyta, D.R. Venom-gland transcriptomics and venom proteomics of the hentz striped scorpion (Centruroides hentzi; Buthidae) reveal high toxin diversity in a harmless member of a lethal family. Toxicon 2018, 142, 14–29. [Google Scholar] [CrossRef]
- Pedraza Escalona, M.; Possani, L.D. Scorpion beta-toxins and voltage-gated sodium channels: Interactions and effects. Front. Biosci. 2013, 18, 572–587. [Google Scholar] [CrossRef]
- Gurevitz, M. Mapping of scorpion toxin receptor sites at voltage-gated sodium channels. Toxicon 2012, 60, 502–511. [Google Scholar] [CrossRef]
- Cid-Uribe, J.I.; Meneses, E.P.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. Dissecting toxicity: The venom gland transcriptome and the venom proteome of the highly venomous scorpion Centruroides limpidus (karsch, 1879). Toxins 2019, 11, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, M.; Peigneur, S.; Tytgat, J. Neurotoxins and their binding areas on voltage-gated sodium channels. Front. Pharm. 2011, 2, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catterall, W.A. Voltage-gated sodium channels at 60: Structure, function and pathophysiology. J. Physiol. 2012, 590, 2577–2589. [Google Scholar] [CrossRef]
- Gordon, D.; Gilles, N.; Bertrand, D.; Molgo, J.; Nicholson, G.M.; Sauviat, M.P.; Benoit, E.; Shichor, I.; Lotan, I.; Gurevitz, M.; et al. Scorpion toxins differentiating among neuronal sodium channel subtypes: Nature’s guide for design of selective drugs. In Perspectives in Molecular Toxinology; Menez, A., Ed.; John Wiley & Sons, Ltd.: New York, NY, USA, 2002; pp. 215–238. [Google Scholar]
- Rendon-Anaya, M.; Delaye, L.; Possani, L.D.; Herrera-Estrella, A. Global Transcriptome Analysis of the Scorpion Centruroides noxius: New Toxin Families and Evolutionary Insights from an Ancestral Scorpion Species. PLoS ONE 2012, 7, e43331. [Google Scholar] [CrossRef] [Green Version]
- Cohen, L.; Karbat, I.; Gilles, N.; Froy, O.; Ilan, N.; Benveniste, M.; Gordon, D.; Gurevitz, M. Common features in the functional surface of scorpion β-toxins and elements that confer specificity for insect and mammalian voltage-gated sodium channels. J. Biol. Chem. 2005, 280, 5045–5053. [Google Scholar] [CrossRef] [Green Version]
- Cohen, L.; Lipstein, N.; Karbat, I.; Ilan, N.; Gilles, N.; Kahn, R.; Gordon, D.; Gurevitz, M. Miniaturization of Scorpion β-Toxins Uncovers a Putative Ancestral Surface of Interaction with Voltage-gated Sodium Channels. J. Biol. Chem. 2008, 282, 15169–15176. [Google Scholar] [CrossRef] [Green Version]
- Del Rio-Portilla, F.; Hernandez-Marin, E.; Pimienta, G.; Coronas, F.V.; Zamudio, F.Z.; Rodriguez de la Vega, R.C.; Wanke, E.; Possani. L.D. NMR solution structure of Cn12, a novel peptide from the Mexican scorpion Centruroides noxius with a typical β-toxin sequence but with α-like physiological activity. Eur. J. Biochem. 2004, 271, 2504–2516. [Google Scholar] [CrossRef] [PubMed]
- Kang, A.M.; Brooks, D.E. Nationwide Scorpion Exposures Reported to US Poison Control Centers from 2005 to 2015. J. Med. Toxicol. 2017, 13, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Rowe, A.H.; Rowe, M.P. Physiological resistance of grasshopper mice (Onychomys spp.) to Arizona bark scorpion (Centruroides exilicauda) venom. Toxicon 2008, 52, 597–605. [Google Scholar] [CrossRef]
- Shelley, R.M.; Sissom, W.D. Distributions of the scorpions Centruroides vittatus (say) and Centruroides hentzi (banks) in the United States and Mexico (scorpiones, buthidae). J. Arach. 1995, 23, 100–110. [Google Scholar]
- Yamashita, T.; Rhoads, D.R. Species delimitation and morphological divergence in the scorpion Centruroides vittatus (Say, 1821): Insights from phylogeography. PLoS ONE 2013, 8, e68282. [Google Scholar] [CrossRef] [Green Version]
- Pucca, M.B.; Cerni, F.A.; Peigneur, S.; Bordon, K.C.; Tytgat, J.; Arantes, E.C. Revealing the Function and the Structural Model of Ts4: Insights into the “Non-Toxic” Toxin from Tityus serrulatus Venom. Toxins 2015, 7, 2534–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, T. Surface activity, biomass, and phenology of the striped scorpion, Centruroides vittatus (Buthidae) in Arkansas, USA. Euscorpius 2004, 17, 25–33. [Google Scholar]
- Corona, M.; Valdez-Cruz, N.A.; Merino, E.; Zurita, M.; Possani, L.D. Genes and peptides from the scorpion centruroides sculpturatus ewing, that recognize Na+-channels. Toxicon 2001, 39, 1893–1898. [Google Scholar] [CrossRef]
- Valdez-Velázquez, L.L.; Quintero-Herna´ndez, V.; Romero-Gutie´rrez, M.T.; Coronas, F.I.V.; Possani, L.D. Mass Fingerprinting of the Venom and Transcriptome of Venom Gland of Scorpion Centruroides tecomanus. PLoS ONE 2013, 8, e66486. [Google Scholar] [CrossRef] [Green Version]
- Valdez-Velázquez, L.L.; Cid-Uribe, J.; Romero-Gutierrez, M.T.; Olamendi-Portugal, T.; Jimenez-Vargas, J.M.; Possani, L.D. Transcriptomic and proteomic analyses of the venom and venom glands of Centruroides hirsutipalpus, a dangerous scorpion from Mexico. Toxicon 2020, 179, 21–32. [Google Scholar] [CrossRef]
- De Oliveira, U.C.; Nishiyama, M.Y., Jr.; Dos Santos, M.; Santos-da-Silva, A.P.; Chalkidis, H.M.; Souza-Imberg, A.; Candido, D.M.; Yamanouye, N.; Dorce, V.; Junqueira-de-Azevedo, I. Proteomic endorsed transcriptomic profiles of venom glands from Tityus obscurus and T. serrulatus scorpions. PLoS ONE 2018, 13, e0193739. [Google Scholar] [CrossRef]
- Grashof, D.; Kerkkamp, H.; Afonso, S.; Archer, J.; Harris, D.J.; Richardson, M.K.; Vonk, F.J.; van der Meijden, A. Transcriptome annotation and characterization of novel toxins in six scorpion species. BMC Genom. 2019, 20, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, F.N.; Mortari, M.R.; Carneiro, F.P.; Guerrero-Vargas, J.A.; Santos, D.M.; Pimenta, A.M.; Schwartz, E.F. Another record of significant regional variation in toxicity of Tityus serrulatus venom in Brazil: A step towards understanding the possible role of sodium channel modulators. Toxicon 2013, 73, 33–46. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The clustal_x windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Geneious v4.6. Available online: http://www.geneious.com/ (accessed on 10 October 2016).
- Romero-Gutierrez, T.; Peguero-Sanchez, E.; Cevallos, M.A.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. A Deeper Examination of Thorellius atrox Scorpion Venom Components with Omic Technologies. Toxins 2017, 9, 399. [Google Scholar] [CrossRef] [Green Version]
- Kuang, J.; Yan, X.; Genders, A.J.; Granata, C.; Bishop, D.J. An overview of technical considerations when using quantitative real-time PCR analysis of gene expression in human exercise research. PLoS ONE 2018, 13, e0196438. [Google Scholar] [CrossRef] [PubMed]
- Fastqc. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 15 July 2019).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.; Bowden, J.; Couger, M.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.; Schein, J.; Chiu, R.; Corbett, R.; Field, M.; Jackman, S.D.; Mungall, K.; Lee, S.; Okada, H.M.; Qian, J.Q.; et al. De novo assembly and analysis of RNA-seq data. Nat. Methods 2010, 7, 909–912. [Google Scholar] [CrossRef]
- Schulz, M.H.; Zerbino, D.R.; Vingron, M.; Birney, E. Oases: Robust de novo RNA-seq assembly across the dynamic range of expression levels. Bioinformatics 2012, 28, 1086–1092. [Google Scholar] [CrossRef]
- Li, R.; Yu, C.; Li, Y.; Lam, T.W.; Yiu, S.M.; Kristiansen, K.; Wang, J. SOAP2: An improved ultrafast tool for short read alignment. Bioinformatics 2009, 25, 1966–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, D.G. Genes of the pig, Sus scrofa, reconstructed with EvidentialGene. PeerJ 2019, 7, e6374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NCGAS/de-Novo-Transcriptome-Assembly-Pipeline (Version 4.0.1). Available online: https://zenodo.org/record/3945505#.YZthaboRVPZ (accessed on 15 July 2019).
- Shi, Y.; Shang, J. Long Noncoding RNA Expression Profiling Using Arraystar LncRNA Microarrays. Methods Mol. Biol. 2016, 1402, 43–61. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| CviNaTBet_HT_02 REGYLAIRGCIYSCVTDRECETECKKYGGKIGFCRFLTCYCEGLPKSVPTVPKPGRKCA |
| 8 populations: HT, AgSp, Chin, LBR, LPb, PaDu, Kisa, KS |
| CviNaTBet_HT_01 KEGYLVNKSDGCKYGCVMLIGDEGCNKECKAPNQGGTKGWCYAFGCWCTGMPESTQVYPLPNKSCGKK |
| 6 populations: HT, AgSp, LBR, LPb, PaDu, Scv |
| CviNaTBet_Chin_05 KEGYLVRKSDNCKHGCIPGIDEDFCDDICKARNRGGKKGWCQKYGCWCTGMPESTQTYPIPGKPCSS |
| 4 populations: AgSp, Chin, BCR, Kisat |
| CviNaTBet_Chin_01 KEGYLVKKSDGCKYGCVMLIGDSNCDMECKAPNQGGQKGWCYAFGCWCTGMPESTQVYPLPGKSCGKK |
| 3 populations: Chin, LBR, Kisat |
| CviNaTBet_HT_08 REGYLVNKKTGCKYGCTPKLGDDHCDKECKAPNQGGKKGWCKNFGCWCTGMPESTQTWPIPGKSCSS |
| 3 populations: HT, AgSp, BCR |
| Na Tox Gene | Diff lwr | upr | p adj | |
|---|---|---|---|---|
| Ha1210-Chin654 | 2.958 | 0.619 | 5.297 | 0.0039 |
| Ha1210-AgSp668 | 3.104 | 0.765 | 5.442 | 0.002 |
| Ha1210-CvIV4 | 2.651 | 0.312 | 4.99 | 0.0149 |
| Assembled Contig Number | 226,162 |
|---|---|
| Assembled contigs > 500 bp | 48,463 |
| Assembled contigs > 1000 bp | 26,909 |
| Average length of assembled contigs (bp) | 463 |
| Longest assembled contig (bp) | 9430 |
| Total length (bp) | 104,750,193 |
| Assembled contig N50 | 1149 |
| MC1 | MT1 | MC2 | MT2 | FC1 | FT2 | |
|---|---|---|---|---|---|---|
| CviNaTBet_Ha1210 | 4 | 425 | 15 | 7638 | 4 | 4134 |
| CviNaTBet_Chin654 | 5 | 502 | 17 | 8923 | 5 | 4932 |
| CviNaTBet_AgSp667 | 3 | 523 | 12 | 6482 | 2 | 329 |
| CviNaTBet_AgSp668 | 4 | 437 | 16 | 8682 | 4 | 3476 |
| JF938594.1 Cv Alpha Tox IV4 | 0 | 224 | 1 | 406 | 0 | 199 |
| AF338462.1 CsEv3b | 4 | 365 | 21 | 11,318 | 4 | 1776 |
| L05060.1_CUDNATOXA C_noxius | ||||||
| Na channel blocker | 4 | 363 | 21 | 11,278 | 4 | 1703 |
| HQ262494.1 Css beta neurotox | 1 | 356 | 4 | 2030 | 1 | 738 |
| AF491130.1 C_limp Na channel Mod tox 5c | 2 | 303 | 7 | 3976 | 1 | 165 |
| AF491134.1 C_limp Na channel 8 | 2 | 227 | 8 | 4031 | 2 | 1411 |
| AY164271.1 C_nox ergtox1 | 1 | 429 | 4 | 1830 | 1 | 469 |
| AY159340.1 C_gracil ergtox1 | 0 | 59 | 0 | 43 | 0 | 54 |
| AY649871.1 C_ex 13 tox | 5 | 542 | 17 | 8941 | 5 | 4344 |
| AY649868.1 C_ex10 | 2 | 151 | 8 | 4086 | 3 | 2657 |
| AF338460.1 CsEIa | 0 | 82 | 3 | 1471 | 1 | 1 |
| AF338450.1 Cse8 | 4 | 386 | 22 | 12,095 | 4 | 1845 |
| AF338448.1 CsE1x | 2 | 268 | 9 | 4712 | 2 | 1565 |
| AY649860.1 Cex2 | 4 | 360 | 21 | 11,258 | 4 | 1714 |
| CviNaTBet_HT_04 | 2 | 121 | 11 | 6028 | 2 | 92 |
| CviNaTBet_HT_08 | 3 | 176 | 10 | 5376 | 2 | 1597 |
| CViNaTBet_HT_09 | 4 | 348 | 13 | 7181 | 4 | 3127 |
| CviNaTBet_Kisa_01 | 2 | 200 | 9 | 4681 | 3 | 2544 |
| CviNaTBet_LBR_03 | 2 | 161 | 8 | 3815 | 2 | 1944 |
| CviNaTBet_LPb_04 | 2 | 122 | 8 | 3700 | 2 | 2255 |
| CviNaTBet_Scv_02 | 3 | 139 | 12 | 6419 | 2 | 176 |
| AF439766.1 C_limpidus 16s ribosomal | 3392 | 2005 | 1894 | 1933 | 2204 | 1268 |
| AH010433.1 CEX RNA poly | 0 | 36 | 37 | 22 | 32 | 43 |
| AY995829.1 C_nox COI | 5951 | 2594 | 3837 | 1696 | 5481 | 2014 |
| AY995849.1 C_Sc COII | 2849 | 1251 | 1719 | 419 | 3458 | 1342 |
| EU381110.1 C_vit COI | 2357 | 1268 | 2393 | 547 | 2788 | 1352 |
| CB334087.1 M_gib EF1 | 244 | 297 | 167 | 78 | 230 | 302 |
| BU092001.1 M_gib EF2 | 1142 | 1103 | 721 | 369 | 881 | 944 |
| CB334044.1 M_gib RPL19 | 255 | 260 | 147 | 105 | 273 | 244 |
| Molecular Weight | Female 1 | Female 2 | Female 3 | Female 4 | Male 1 | Male 2 | Male 3 | |
|---|---|---|---|---|---|---|---|---|
| CviNaTBet_Ha1210 | 7 kDa | 4.66% | 15.08% | 12.69% | 10.03% | 5.79% | 5.66% | 6.91% |
| CviNaTBet_AgSp668 | 7 kDa | 1.09% | 1.88% | 0.58% | 1.23% | na | na | 0.35% |
| alpha-toxin, partial [C. vittatus] | 11 kDa | 0.75% | 0.65% | 1.21% | 0.64% | 0.81% | 0.44% | 0.73% |
| CviNaTBet_HT_01 | 7 kDa | 2.15% | 8.12% | 5.48% | na | 2.70% | na | na |
| CviNaTBet_HT_06 | 7 kDa | 2.07% | 2.94% | 1.66% | 1.05% | 7.13% | 3.62% | 0.79% |
| CviNaTBet_Chin05 | 7 kDa | 1.09% | 1.88% | 1.46% | 1.23% | 0.78% | 3.51% | 0.70% |
| CviNaTBet_BCR_10 | 7 kDa | 1.09% | 1.88% | 1.46% | 1.23% | 0.78% | 3.51% | 0.70% |
| alpha-toxin CvIV4 [C. sculpturatus] | 9 kDa | 0.29% | 0.95% | 0.47% | 0.25% | 0.32% | 0.52% | 0.28% |
| RecName: Full = Toxin Cg2 | 8 kDa | na | na | 0.87% | 0.30% | na | na | 0.35% |
| RecName: Full = Alpha-toxin Cn12 | 7 kDa | na | na | 0.29% | 0.31% | na | na | 0.70% |
| beta-toxin Im-2-like [C. sculpturatus] | 9 kDa | na | na | 0.23% | na | na | na | 0.55% |
| beta-toxin CeII8-like [C. sculpturatus] | 10 kDa | na | na | 0.23% | 0.25% | na | na | na |
| RecName: Full = Alpha-toxin CsE5 | 7 kDa | na | na | 0.31% | 0.33% | na | na | 0.37% |
| beta-neurotoxin CssIX precursor [C. suffusus suffusus] | 9 kDa | na | na | 0.24% | na | na | na | na |
| Scorpion Species | Sodium Channel Toxin Transcript Diversity | Sodium Channel Toxin Protein Diversity | Reference |
|---|---|---|---|
| Centruroides vittatus | 19 | 7 | This paper |
| Centruroides vittatus | 13 | - | McElroy et al. 2017 [9] |
| Centruroides hentzi | 36 | 9 | Ward et al. 2018 [10] |
| Centruroides sculpturatus * | 22 | - | Corona et al. 2001 [27] |
| Centruroides sculpturatus * | - | 5/6 | Carcamo-Noriega et al. 2018 [6] |
| Centruroides limpidus * | 59 | 26 | Cid-Uribe et al. 2019 [13] |
| Centruroides noxius * | 27 | - | Rendón-Anaya et al. 2012 [17] |
| Centruroides tecomanus * | 24 | 30 | Valdez-Velázquez et al. 2013 [28] |
| Centruroides hirsutipalpus * | 77 | 31 | Valdez-Velázquez et al. 2020 [29] |
| Tityus obscurus * | 48 | 3 | Oliveira et al. 2018 [30] |
| Tityus serrulatus * | 24 | 7 | Oliveira et al. 2018 [30] |
| Hottentotta gentili * | 52 | - | Grashof et al. 2019 [31] |
| Androctonus mauretanicus * | 42 | - | Grashof et al. 2019 [31] |
| Babycurus gigas | 13 | - | Grashof et al. 2019 [31] |
| Grosphus grandideri | 22 | - | Grashof et al. 2019 [31] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bowman, A.; Fitzgerald, C.; Pummill, J.F.; Rhoads, D.D.; Yamashita, T. Reduced Toxicity of Centruroides vittatus (Say, 1821) May Result from Lowered Sodium β Toxin Gene Expression and Toxin Protein Production. Toxins 2021, 13, 828. https://doi.org/10.3390/toxins13110828
Bowman A, Fitzgerald C, Pummill JF, Rhoads DD, Yamashita T. Reduced Toxicity of Centruroides vittatus (Say, 1821) May Result from Lowered Sodium β Toxin Gene Expression and Toxin Protein Production. Toxins. 2021; 13(11):828. https://doi.org/10.3390/toxins13110828
Chicago/Turabian StyleBowman, Aimee, Chloe Fitzgerald, Jeff F. Pummill, Douglas D. Rhoads, and Tsunemi Yamashita. 2021. "Reduced Toxicity of Centruroides vittatus (Say, 1821) May Result from Lowered Sodium β Toxin Gene Expression and Toxin Protein Production" Toxins 13, no. 11: 828. https://doi.org/10.3390/toxins13110828
APA StyleBowman, A., Fitzgerald, C., Pummill, J. F., Rhoads, D. D., & Yamashita, T. (2021). Reduced Toxicity of Centruroides vittatus (Say, 1821) May Result from Lowered Sodium β Toxin Gene Expression and Toxin Protein Production. Toxins, 13(11), 828. https://doi.org/10.3390/toxins13110828