Genetic Responses and Aflatoxin Inhibition during Co-Culture of Aflatoxigenic and Non-Aflatoxigenic Aspergillus flavus


 ,
,  ,
,  , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Aflatoxin
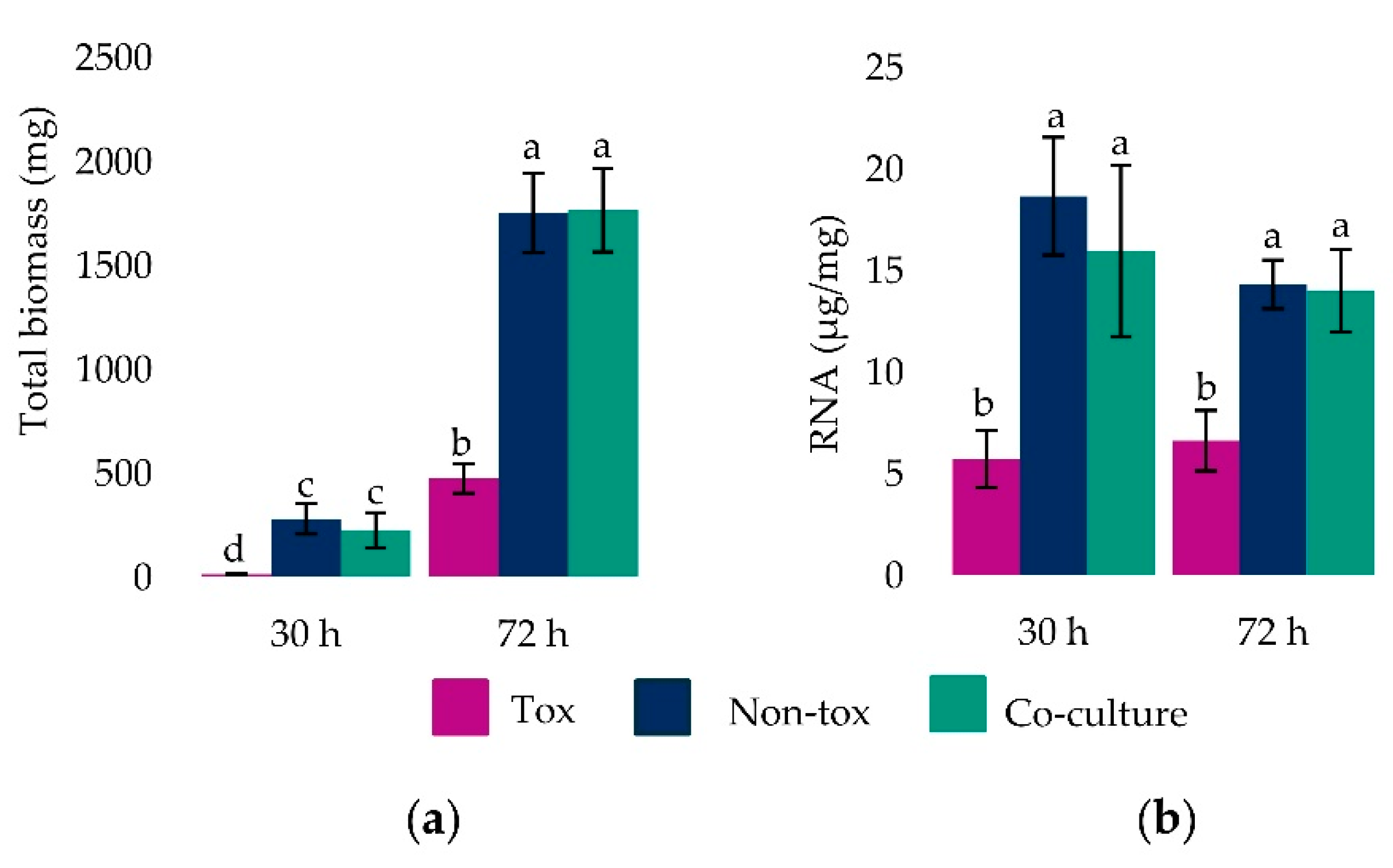
2.2. Fungal Biomass and Total RNA
2.3. RNA Sequencing
2.3.1. Equating Growth with RNA Production
2.3.2. Differential Gene Expression
2.3.3. Functional Analysis of Differentially Expressed Genes
2.3.4. Gene Expression in the Aflatoxin Biosynthesis Cluster
2.3.5. Genes Highly Upregulated in Biocontrol Non-Tox 17 Compared to Tox 53
2.3.6. Genes Upregulated in Response to Tox 53
2.3.7. Differential Expression of Imizoquin Biosynthesis Genes
3. Discussion
4. Materials and Methods
4.1. Fungal Isolates
4.2. Biocontrol Interaction Cultural Experimental Design
4.3. Aflatoxin Extraction and Quantification
4.4. Whole Fungal Mycelia Harvest and RNA Extraction
4.5. RNA Sequencing and Analysis
4.6. Other Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horn, B.W. Ecology and population biology of aflatoxigenic fungi in soil. J. Toxicol. Toxin Rev. 2003, 22, 351–379. [Google Scholar] [CrossRef]
- Mitchell, N.J.; Bowers, E.; Hurburgh, C.; Wu, F. Potential economic losses to the USA corn industry from aflatoxin contamination. Food Addit. Contam. A 2016, 33, 540–550. [Google Scholar] [CrossRef]
- Richard, J.L. Discovery of aflatoxins and significant historical features. Toxin Rev. 2008, 27, 171–201. [Google Scholar] [CrossRef]
- Wicklow, D.T. Epidemiology of Aspergillus flavus in Corn. In Aflatoxin in Corn: New Perspectives; Shotwell, O.L., Hurburgh, C.R., Jr., Eds.; Research Bulletin—Iowa State University, Agricultural and Home Economics Experiment Station: Ames, IA, USA, 1991; Volume 599, pp. 315–328. [Google Scholar]
- Yu, J.; Cleveland, T.E.; Nierman, W.C.; Bennett, J.W. Aspergillus flavus genomics: Gateway to human and animal health, food safety, and crop resistance to diseases. Rev. Iberoam. Micol. 2005, 22, 194–202. [Google Scholar] [CrossRef]
- Cotty, P.J. Virulence and cultural characteristics of two Aspergillus flavus strains pathogenic on cotton. Phytopathology 1989, 79, 808–814. [Google Scholar] [CrossRef]
- Geiser, D.M.; Pitt, J.I.; Taylor, J.W. Cryptic speciation and recombination in the aflatoxin-producing fungus Aspergillus flavus. Proc. Natl. Acad. Sci. USA 1998, 95, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Geiser, D.M.; Dorner, J.W.; Horn, B.W.; Taylor, J.W. The phylogenetics of mycotoxin and sclerotium production in Aspergillus flavus and Aspergillus oryzae. Fungal Genet. Biol. 2000, 31, 169–179. [Google Scholar] [CrossRef]
- Probst, C.; Callicott, K.A.; Cotty, P.J. Deadly strains of Kenyan Aspergillus are distinct from other aflatoxin producers. Eur. J. Plant Pathol. 2012, 132, 419–429. [Google Scholar] [CrossRef]
- Singh, P.; Orbach, M.J.; Cotty, P.J. Aspergillus texensis: A novel aflatoxin producer with S morphology from the United States. Toxins 2018, 10, 513. [Google Scholar] [CrossRef]
- Singh, P.; Callicott, K.A.; Orbach, M.J.; Cotty, P.J. Molecular analysis of S-morphology aflatoxin producers from the United States reveals previously unknown diversity and two new taxa. Front. Microbiol. 2020, 11, e1236. [Google Scholar] [CrossRef]
- Varga, J.; Frisvad, J.C.; Samson, R.A. A reappraisal of fungi producing aflatoxins. World Mycotoxin J. 2009, 2, 263–277. [Google Scholar] [CrossRef]
- Probst, C.; Njapau, H.; Cotty, P.J. Outbreak of an acute aflatoxicosis in Kenya in 2004: Identification of the causal agent. Appl. Environ. Microb. 2007, 7, 2762–2764. [Google Scholar] [CrossRef]
- Probst, C.; Schulthess, F.; Cotty, P.J. Impact of Aspergillus section Flavi community structure of the development of lethal levels of aflatoxins in Kenyan maize (Zea mays). J. Appl. Microbiol. 2010, 108, 600–610. [Google Scholar] [CrossRef]
- Abbas, H.K.; Weaver, M.A.; Horn, B.W.; Carbone, I.; Monacell, J.T.; Shier, W.T. Selection of Aspergillus flavus isolates for biological control of aflatoxins in corn. Toxin Rev. 2011, 30, 59–70. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; Ortega-Beltran, A.; Akande, A.; Mutegi, C.; Atehnkeng, J.; Kaptoge, L.; Senghor, A.L.; Adhikari, B.N.; Cotty, P.J. Biological control of aflatoxins in Africa: Current status and potential challenges in the face of climate change. World Mycotoxin J. 2016, 9, 771–789. [Google Scholar] [CrossRef]
- Cotty, P.J. Influence of field application of an atoxigenic strain of Aspergillus flavus on the populations of A. flavus infecting cotton bolls and on the aflatoxin content of cottonseed. Phytopathology 1994, 84, 1270–1277. [Google Scholar] [CrossRef]
- Cotty, P.J.; Bhatnagar, D. Variability among atoxigenic Aspergillus flavus strains in ability to prevent aflatoxin contamination and production of aflatoxin biosynthetic pathway enzymes. Appl. Environ. Microb. 1994, 60, 2248–2251. [Google Scholar] [CrossRef]
- Dorner, J.W.; Cole, R.J. Effect of application of nontoxigenic strains of Aspergillus flavus and A. parasiticus on subsequent aflatoxin contamination of peanuts in storage. J. Stored Prod. Res. 2002, 38, 329–339. [Google Scholar] [CrossRef]
- Moral, J.; Garcia-Lopez, M.T.; Camiletti, B.X.; Jaime, R.; Michailides, T.J.; Bandyopadhyay, R.; Ortega-Beltran, A. Present status and perspective on the future use of aflatoxin biocontrol products. Agronomy 2020, 10, 491. [Google Scholar] [CrossRef]
- Pitt, J.I. The pros and cons of using biocontrol by competitive exclusion as a means for reducing aflatoxin in maize in Africa. World Mycotoxin J. 2019, 12, 103–112. [Google Scholar] [CrossRef]
- Ezekiel, C.N.; Ortega-Beltran, A.; Oyedeji, E.O.; Atehnkeng, J.; Kössler, P.; Tairu, F.; Hoeschle-Zeledon, I.; Karlovsky, P.; Cotty, P.J.; Bandyopadhyay, R. Aflatoxin in chili peppers in Nigeria: Extent of contamination and control using atoxigenic Aspergillus flavus genotypes as biocontrol agents. Toxins 2019, 11, 429. [Google Scholar] [CrossRef]
- Alaniz Zanon, M.S.; Barros, G.G.; Chulze, S.N. Non-aflatoxigenic Aspergillus flavus as potential biocontrol agents to reduce aflatoxin contamination in peanuts harvested in Northern Argentina. Int. J. Food Microbiol. 2016, 16, 63–68. [Google Scholar] [CrossRef]
- Savić, Z.; Dudaš, T.; Loc, M.; Grahovac, M.; Budakov, D.; Jajić, I.; Krstović, S.; Barošević, T.; Krska, R.; Sulyok, M.; et al. Biological Control of Aflatoxin in Maize Grown in Serbia. Toxins 2020, 12, 162. [Google Scholar] [CrossRef] [PubMed]
- Cotty, P.J.; Bayman, P. Competitive exclusion of atoxigenic strain of Aspergillus flavus by an atoxigenic strain. Phytopathology 1993, 83, 1283–1287. [Google Scholar] [CrossRef]
- Dorner, J.W.; Cole, R.J.; Connick, W.J.; Daigle, D.J.; McGuire, M.R.; Shasha, B.S. Evaluation of biological control formulations to reduce aflatoxin contamination in peanuts. Biol. Control 2003, 26, 318–324. [Google Scholar] [CrossRef]
- Horn, B.W.; Dorner, J.W. Effect of nontoxigenic Aspergillus flavus and A. parasiticus on aflatoxin contamination of wounded peanut seeds inoculated with agricultural soil containing natural fungal populations. Biocontrol Sci. Technol. 2009, 19, 249–262. [Google Scholar] [CrossRef]
- Weaver, M.A.; Abbas, H.K. Field displacement of aflatoxigenic Aspergillus flavus strains through repeated biological control applications. Front. Microbiol. 2019, 10, e1788. [Google Scholar] [CrossRef]
- Damann, K.E., Jr. Atoxigenic Aspergillus flavus biological control of aflatoxin contamination: What is the mechanism? World Mycotoxin J. 2015, 8, 235–244. [Google Scholar] [CrossRef]
- Ehrlich, K.C. Non-aflatoxigenic Aspergillus flavus to prevent aflatoxin contamination in crops: Advantages and limitations. Front. Microbiol. 2014, 5, e50. [Google Scholar] [CrossRef]
- Moore, G.G. Practical considerations will ensure the continued success of pre-harvest biocontrol using non-aflatoxigenic Aspergillus flavus strains. Crit. Rev. Food Sci. Nutr. 2021, 1–18. [Google Scholar] [CrossRef]
- Wicklow, D.T.; Bobell, J.R.; Palmquist, D.E. Effect of intraspecific competition by Aspergillus flavus on aflatoxin formation in suspended disc culture. Mycol. Resour. 2003, 107, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Mehl, H.L.; Cotty, P.J. Variation in competitive ability among isolates of Aspergillus flavus from different vegetative compatibility groups during maize infection. Phytopathology 2010, 100, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Jha, A.; Sweany, R.; DeRobertis, C.; Damann, K.E., Jr. Intraspecific aflatoxin inhibition in Aspergillus flavus is thigmoregulated, independent of vegetative compatibility group and is strain dependent. PLoS ONE 2011, 6, e23470. [Google Scholar] [CrossRef] [PubMed]
- Mauro, A.; Garcia-Cela, E.; Pietri, A.; Cotty, P.J.; Battilani, P. Biological control products for aflatoxin prevention in Italy: Commercial field evaluation of atoxigenic Aspergillus flavus active ingredients. Toxins 2018, 10, 30. [Google Scholar] [CrossRef]
- Lanubile, A.; Maschietto, V.; Battilani, P.; Marocco, A. Infection with toxigenic and atoxigenic strains of Aspergillus flavus induces different transcriptional signatures in maize kernels. J. Plant Interact. 2017, 12, 21–30. [Google Scholar] [CrossRef][Green Version]
- Alshannaq, A.F.; Gibbons, J.G.; Lee, M.-K.L.; Han, K.-H.; Hong, S.-B.; Yu, J.-H. Controlling aflatoxin contamination and propagation of Aspergillus flavus by a soy-fermenting Aspergillus oryzae strain. Sci. Rep. 2018, 8, e16871. [Google Scholar] [CrossRef] [PubMed]
- Moore, G.G.; Lebar, M.D.; Carter-Wientjes, C.H. The role of extrolites secreted by nonaflatoxigenic Aspergillus flavus in biocontrol efficacy. J. Appl. Microbiol. 2019, 126, 1257–1264. [Google Scholar] [CrossRef]
- Sweany, R.R. Investigations into Aspergillus flavus Infection of Corn and Regulation of Aflatoxin Production by Volatiles and Biocontrol Strains. Ph.D. Thesis, Louisiana State University, Baton Rouge, LA, USA, 2019; p. e4780. [Google Scholar]
- Sweany, R.R.; DeRobertis, D.D.; Damann, K.E., Jr. Intra-specific growth and aflatoxin inhibition responses to atoxigenic Aspergillus flavus: Evidence of secreted, inhibitory substance(s) in biocontrol. Phytopathology 2021. in review. [Google Scholar]
- Maxwell, L.A.; Callicott, K.A.; Bandyopadhyay, R.; Mehl, H.L.; Orbach, M.J.; Cotty, P.J. Degradation of aflatoxins B1 by atoxigenic Aspergillus flavus biocontrol agents. Plant Dis. First Look 2021. [Google Scholar] [CrossRef]
- Sweany, R.R.; Damann, K.E., Jr.; Kaller, M.D. Comparison of soil and corn kernel Aspergillus flavus populations: Evidence for niche specialization. Phytopathology 2011, 101, 952–959. [Google Scholar] [CrossRef]
- Doyle, M.P.; Marth, E.H. Bisulfite degrades aflatoxin: Effect of citric acid and methanol and possible mechanism of degradation. J. Food Prot. 1978, 41, 891–896. [Google Scholar] [CrossRef]
- Khalid, N.; Seifuddin, F.T.; Turner, G.; Haft, D.; Nierman, W.C.; Wolfe, K.H.; Fedorova, N.D. SMURF: Genomic mapping of fungal secondary metabolite clusters. Fungal Genet. Biol. 2010, 47, 736–741. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Fedorova, N.D.; Burroughs, J.L.; Dolezal, A.L.; Bok, J.W.; Horowitz-Brown, S.; Woloshuk, C.P.; Yu, J.; Keller, N.P.; Payne, G.A. Beyond aflatoxin: Four distinct expression patterns and functional roles associated with Aspergillus flavus secondary metabolism gene clusters. Mol. Plant Pathol. 2010, 11, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.-M.; Szewczyk, E.; Davidson, A.D.; Keller, N.; Oakley, B.R.; Wang, C.C.C. A gene cluster containing two fungal polyketide synthases encodes the biosynthetic pathway for a polyketide, asperfuranone, in Aspergillus nidulans. J. Am. Chem. Soc. 2009, 131, 2965–2970. [Google Scholar] [CrossRef]
- Love, M.I.; Wolfgang, H.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Medema, M.H.; Blin, K.; Cimermancic, P.; De Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: Rapid identification, annotation, and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Resour. 2011, 39, W339–W346. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.L.; Nielsen, J.B.; Rank, C.; Klejnstrup, M.L.; Holm, D.K.; Brogaard, K.H.; Hansen, B.G.; Frisvad, J.C.; Larsen, T.O.; Mortensen, U.H. A genome-wide polyketide synthase deletion library uncovers novel genetic links to polyketides and meroterpenoids in Aspergillus nidulans. FEMS Microbiol. Lett. 2011, 321, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Matsuda, Y.; Gao, H.; Hu, D.; Yao, X.S.; Abe, I. Biosynthesis of LL-Z1272β: Discovery of a new member of NRPS-like enzymes for aryl-aldehyde formation. ChemBioChem 2016, 10, 904–907. [Google Scholar] [CrossRef] [PubMed]
- Ammar, H.A.M.; Srour, A.Y.; Ezzat, S.M.; Hoseny, A.M. Identification and characterization of genes involved in kojic acid biosynthesis in Aspergillus flavus. Ann. Microbiol. 2017, 67, 691–702. [Google Scholar] [CrossRef]
- Khalid, S.; Baccile, J.A.; Spraker, J.E.; Tannous, J.; Imran, M.; Schroeder, F.C.; Keller, N.P. NRPS-derived isoquinolines and lipopetides mediate antagonism between plant pathogenic fungi and bacteria. ACS Chem. Biol. 2018, 13, 171–179. [Google Scholar] [CrossRef]
- Chankhamjon, P.; Boettger-Schmidt, D.; Scherlach, K.; Urbansky, B.; Lackner, G.; Kalb, D.; Dahse, H.-M.; Hoffmeister, D.; Hertweck, C. Biosynthesis of the halogenated mycotoxin aspirochlorine in koji mold involves a cryptic amino acid conversion. Angew. Chem. Int. Ed. 2014, 53, 13409–13413. [Google Scholar] [CrossRef] [PubMed]
- Baidya, S.; Duran, R.M.; Lohmar, J.M.; Harris-Coward, P.Y.; Cary, J.W.; Hong, S.-Y.; Roze, L.V.; Linz, J.E.; Calvo, A.M. VeA is associated with the response to oxidative stress in the aflatoxin producer Aspergillus flavus. Eukaryot. Cell 2014, 13, 1095–1103. [Google Scholar] [CrossRef]
- Fountain, J.C.; Scully, B.T.; Chen, Z.-Y.; Gold, S.E.; Glenn, A.E.; Abbas, H.K.; Lee, R.D.; Kemerait, R.C.; Guo, B. Effects of hydrogen peroxide on different toxigenic and atoxigenic isolates of Aspergillus flavus. Toxins 2015, 7, 2985–2999. [Google Scholar] [CrossRef] [PubMed]
- Fountain, J.C.; Bajaj, P.; Pandey, M.; Nayak, S.N.; Yang, L.; Kumar, V.; Jayale, A.S.; Chitikineni, A.; Zhuang, W.; Scully, B.T.; et al. Oxidative stress and carbon metabolism influence Aspergillus flavus transcriptome composition and secondary metabolite production. Sci. Rep. 2016, 6, e38747. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M.; Mukerji, K.G.; Raj, H.G. Positive correlation exists between glutathione S-transferase activity and aflatoxin formation in Aspergillus flavus. Biochem. J. 1988, 254, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Ziglari, T.; Allameh, A.; Razzaghi-Abyaneh, M.; Khosravi, A.R.; Yadegari, M.H. Comparison of glutathione S-transferase activity and concentration in aflatoxin-producing and their non-toxigenic counterpart isolates. Mycopathologia 2008, 166, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Hua, S.S.T.; Parfitt, D.E.; Sarreal, S.B.L.; Sidhu, G. Dual culture of atoxigenic and toxigenic strains of Aspergillus flavus to gain insight into repression of aflatoxin biosynthesis and fungal interaction. Mycotoxin Res. 2019, 35, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Geng, Q.; Song, F.; He, X.; Hu, T.; Wang, S.; Tian, J. Transcriptome sequencing revealed an inhibitory mechanism of Aspergillus flavus asexual development and aflatoxin metabolism by soy-fermenting non-aflatoxigenic Aspergillus. Int. J. Mol. Sci. 2020, 21, 6994. [Google Scholar] [CrossRef]
- Burdock, G.A.; Soni, M.G.; Carabin, I.G. Evaluation of health aspects of Kojic acid in food. Regul. Toxicol. Pharm. 2001, 33, 80–101. [Google Scholar] [CrossRef]
- Schroeckh, V.; Scherlach, K.; Nützmann, H.-W.; Shelest, E.; Schmidt-Heck, W.; Schuemann, J.; Martin, K.; Hertweck, C.; Brakhage, A.A. Intimate bacterial-fungal interaction triggers biosynthesis of archetypal polyketides in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 2009, 106, 14558–14563. [Google Scholar] [CrossRef]
- Wang, C.C.; Chiang, Y.M.; Praseuth, M.B.; Kuo, P.L.; Liang, H.L.; Hsu, Y.L. Asperfuranone from Aspergillus nidulans inhibits proliferation of human non-small cell lung cancer A549 cells via blocking cell cycle progression and inducing apoptosis. Basic Clin. Pharmacol. Toxicol. 2010, 107, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Spraker, J.E.; Jewell, K.; Roze, L.V.; Scherf, J.; Ndagano, D.; Beaudry, R.; Linz, J.E.; Allen, C.; Keller, N.P. A volatile relationship: Profiling an inter-kingdom dialogue between two plant pathogens, Ralstonia Solanacearum and Aspergillus Flavus. J. Chem. Ecol. 2014, 40, 502–513. [Google Scholar] [CrossRef] [PubMed]
- Reyes Pineda, J.A. Characterization of Aspergillus flavus Soil and Corn Kernel Populations from Eight Mississippi River States. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2017; p. e4350. [Google Scholar]
- Sweany, R.R.; Damann, K.E., Jr. Influence of neighboring clonal-colonies on aflatoxin production by Aspergillus flavus. Front. Microbiol. 2020, 10, e3038. [Google Scholar] [CrossRef]
- Moore, G.G.; Lebar, M.D.; Carter-Wientjes, C.H.; Gilbert, M.K. The potential role of fungal volatile organic compounds in Aspergillus flavus biocontrol efficacy. Biol. Control 2021, 160, e104686. [Google Scholar] [CrossRef]
- Sobolev, V.S.; Dorner, J.W. Cleanup procedure for determination of aflatoxins in major agricultural commodities by liquid chromatography. J. AOAC Int. 2002, 85, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Joshua, H. Determination of aflatoxins by reversed-phase high performance liquid chromatography with post-column in-line photochemical derivatization and fluorescence detection. J. Chromatogr. 1993, 654, 247–254. [Google Scholar] [CrossRef]
- Raruang, Y.; Omolehin, O.; Hu, D.; Wei, Q.; Han, Z.Q.; Rajasekaran, K.; Cary, J.W.; Wang, K.; Chen, Z.Y. Host induced gene silencing targeting Aspergillus flavus aflM reduced aflatoxin contamination in transgenic maize under field conditions. Front. Microbiol. 2020, 11, e754. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, B. BBTools Software Package. 2014. Available online: http://sourceforge.net/projects/bbmap578 (accessed on 8 April 2019).
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Garrison, E.; Marth, G. Haplotype-based variant detection from short-read sequencing. arXiv 2012, arXiv:1207.3907. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome project data processing subgroup. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- De Matos Simoes, R.; Emmert-Streib, F. Bagging statistical network inference from large-scale gene expression data. PLoS ONE 2012, 7, e33624. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Evans, C.; Hardin, J.; Stoebel, D.M. Selecting between-sample RNA-Seq normalization methods from the perspective of their assumptions. Brief. Bioinform. 2018, 19, 776–792. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| 30 h | 72 h | 96 h | |
|---|---|---|---|
| Cultures | Aflatoxin B1 ppb ± S. D. | ||
| Tox 53 | <0.05 c 1 | 680 ± 35 b | 1902 ± 163 a |
| Non-tox 17 | <0.05 c | <0.05 c | <0.05 c |
| Co-culture | 0.2 ± 0.1 c | 1.8 ± 0.2 c | <0.05 c |
| 30 h | 72 h | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Rep 1 | Total 2 | Non-Tox 3 | Tox 4 | P Tox 5 | Rep 1 | Total 2 | Non-Tox 3 | Tox 4 | P Tox 5 | |
| Tox 53 | a | 14.2 | 0.092 | 4.336 | 0.979 | d | 8.6 | 0.050 | 2.985 | 0.984 |
| b | 17.4 | 0.105 | 5.143 | 0.980 | e | 15.9 | 0.087 | 5.484 | 0.984 | |
| c | 14.3 | 0.085 | 4.206 | 0.980 | f | 14.1 | 0.105 | 4.817 | 0.979 | |
| Non-tox 17 | a | 16.7 | 5.107 | 0.082 | 0.016 | d | 16.9 | 5.819 | 0.085 | 0.014 |
| b | 16.6 | 5.047 | 0.084 | 0.016 | e | 14.7 | 5.075 | 0.075 | 0.015 | |
| c | 18 | 5.358 | 0.090 | 0.017 | f | 14.2 | 5.040 | 0.074 | 0.014 | |
| Co-culture | a | 29.6 | 8.267 | 0.299 | 0.035 | d | 14.0 | 4.891 | 0.140 | 0.028 |
| b | 12.5 | 3.902 | 0.127 | 0.032 | e | 15.1 | 5.220 | 0.181 | 0.033 | |
| c | 11.3 | 3.510 | 0.120 | 0.033 | f | 29.1 | 9.497 | 0.261 | 0.027 | |
| 30 h | 72 h | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Functional Annotation Terms 1 | Genes 2 | Non vs. Tox 3 | Co vs. Tox | Co vs. Non | Non vs. Tox | Co vs. Tox | Co vs. Non | ||||||
| Oxidation/reduction | 1477 | 274 | -229 | −235 | −43 | 277 | 284 | ||||||
| Signal peptide | 1188 | 236 | −182 | 221 | −176 | 28 | 220 | 212 | |||||
| Extracellular | 876 | −146 | −140 | 21 | 180 | 175 | |||||||
| Apoplastic | 537 | 114 | −93 | −91 | 19 | 121 | 119 | ||||||
| NAD(P)-binding | 596 | −104 | −118 | 114 | 124 | ||||||||
| Oxidoreductase activity | 542 | −94 | 76 | −95 | 102 | 113 | |||||||
| Major facilitator family | 389 | 87 | 89 | ||||||||||
| Alpha/Beta hydrolase | 313 | 85 | 81 | ||||||||||
| Zn(2)-C6 transcr. factor | 289 | 67 | 69 | 57 | 57 | ||||||||
| Iron ion binding | 266 | 64 | 58 | ||||||||||
| FAD/NAD(P)-binding | 289 | −13 | 55 | 56 | |||||||||
| Heme binding | 246 | 48 | 50 | ||||||||||
| Monooxygenase activity | 214 | −12 | 46 | 49 | |||||||||
| FAD binding | 179 | 42 | 37 | ||||||||||
| Cytochrome P450 | 166 | 40 | 41 | ||||||||||
| GroES-like | 117 | −30 | −33 | ||||||||||
| Fatty acid biosynthesis | 99 | −26 | −24 | ||||||||||
| Alcohol dehydrogenase | 93 | −26 | −29 | ||||||||||
| Peroxisome | 71 | −25 | −23 | ||||||||||
| Polyketide synthase | 94 | −24 | −28 | ||||||||||
| Tyrosine metabolism | 71 | 23 | 20 | ||||||||||
| ATPase movement | 66 | 18 | 19 | ||||||||||
| Isomerase activity | 56 | −19 | −20 | ||||||||||
| AMP-binding site | 57 | −18 | |||||||||||
| Phenylalanine metab. | 42 | 15 | 14 | ||||||||||
| Aflatoxin synth. cluster | 29 | −14 | −11 | 16 | −28 | −28 | 9 | ||||||
| Obsolete peroxidase rxn | 34 | −14 | −14 | ||||||||||
| Sterigmatocystin synth. | 25 | 7 | −14 | −13 | 5 | ||||||||
| Imizoquin synth. | 11 | 7 | −6 | 11 | 11 | ||||||||
| Crotonase superfamily | 19 | −9 | −11 | ||||||||||
| L-phenylalanine metab. | 23 | 10 | |||||||||||
| Styrene catabolism | 20 | 10 | 9 | ||||||||||
| Aromatic amino acid | 20 | 10 | 10 | ||||||||||
| Phosphorelay sensor | 20 | −10 | −8 | ||||||||||
| Protein histidine kinase | 20 | −10 | −8 | ||||||||||
| Disulphide reductase | 10 | 8 | 9 | ||||||||||
| Enoyl-CoA hydratase | 9 | −7 | −7 | ||||||||||
| Sulfatase, conserved site | 8 | 6 | 6 | 5 | |||||||||
| Asperfuranone synth. | 5 | 5 | 5 | ||||||||||
| Mycotoxin biosynthesis | 4 | 4 | 4 | ||||||||||
| Cyclopiazonic acid | 4 | −4 | −4 | 3 | −4 | −4 | 2 | ||||||
| Haem peroxidase | 4 | −4 | −4 | ||||||||||
| 30 h 1 | 72 h | |||||||
|---|---|---|---|---|---|---|---|---|
| Gene ID 2 | Chr 3 | Non vs. Tox | Co vs. Tox | Co vs. Non | Non vs. Tox | Co vs. Tox | Co vs. Non | Gene Synonyms or Putative Function |
| AFLA_139150 | 3 | - | - | - | −15.1 | −11.3 | - | aflY/hypA/hypP |
| AFLA_139160 | 3 | - | - | - | −14.8 | −10.8 | - | aflX/ordB |
| AFLA_139170 | 3 | - | 5.1 | 5.1 | −16.1 | −11.3 | - | aflW/moxY |
| AFLA_139180 | 3 | - | - | 5.3 | −15.2 | −10.7 | - | aflV/cypX |
| AFLA_139190 | 3 | - | 4.4 | 6.8 | −15.3 | −10.9 | - | aflK/vbs/verB |
| AFLA_139200 | 3 | - | - | - | −14.3 | −10.9 | - | aflQ/ordA/ord-1 |
| AFLA_139210 | 3 | - | - | - | −15.5 | −10.7 | - | aflP/omtA/omt-1 |
| AFLA_139220 | 3 | - | - | - | −15.2 | −9.9 | - | aflO/omtB/dmtA |
| AFLA_139230 | 3 | - | - | - | −9.8 | −9.1 | - | aflI/avfA |
| AFLA_139240 | 3 | - | - | - | - | - | - | |
| AFLA_139250 | 3 | - | - | - | −15.3 | −11.0 | - | aflL/verB |
| AFLA_139260 | 3 | - | - | - | −15.1 | −10.2 | - | aflG/avnA/ord-1 |
| AFLA_139270 | 3 | −11.3 | −5.3 | 6.0 | −16.1 | −9.1 | 7.0 | |
| AFLA_139280 | 3 | - | - | 4.4 | −14.2 | −9.8 | - | aflN/verA |
| AFLA_139290 | 3 | - | - | - | −11.9 | −12.1 | - | aflMa/hypE |
| AFLA_139300 | 3 | - | 3.0 | 3.5 | −17.0 | −10.5 | 6.5 | aflM/ver-1 |
| AFLA_139310 | 3 | −5.9 | - | 5.9 | −16.0 | −11.9 | - | aflE/norA/aad/adh-2 |
| AFLA_139320 | 3 | −9.6 | −4.7 | 4.9 | −15.7 | −10.5 | 5.2 | aflJ/estA |
| AFLA_139330 | 3 | −9.9 | −4.0 | 5.9 | −16.0 | −10.7 | - | aflH/adhA |
| AFLA_139340 | 3 | −11.6 | −4.9 | 6.7 | −15.0 | −9.8 | 5.3 | aflS |
| AFLA_139360 | 3 | −11.6 | −5.1 | 6.6 | −11.8 | −7.9 | - | aflR/apa-2/afl-2 |
| AFLA_139370 | 3 | −10.2 | −4.1 | 6.1 | −15.1 | −10.3 | 4.7 | aflB/fas-1 |
| AFLA_139380 | 3 | −9.2 | −3.6 | 5.6 | −14.7 | −10.4 | 4.2 | aflA/fas-2/hexA |
| AFLA_139390 | 3 | −8.1 | −2.5 | 5.5 | −15.8 | −11.2 | 4.5 | aflD/nor-1 |
| AFLA_139400 | 3 | −5.6 | −3.2 | - | −15.0 | −10.9 | - | aflCa/hypC |
| AFLA_139410 | 3 | −8.2 | −4.1 | 4.1 | −16.0 | −10.5 | 5.5 | aflC/pksA/pksL1 |
| AFLA_139420 | 3 | −8.2 | −3.1 | 5.0 | −14.4 | −8.7 | 5.7 | aflT/aflT |
| AFLA_139430 | 3 | −4.0 | - | - | −11.2 | −9.1 | - | aflU/cypA |
| AFLA_139440 | 3 | −3.5 | - | - | −10.2 | −8.6 | - | aflF/norB |
| AFLA_139450 | 3 | - | - | - | - | - | - | |
| AFLA_139460 | 3 | −9.3 | −5.7 | - | −14.2 | −9.7 | - | MFS multidrug transporter |
| AFLA_139470 | 3 | −14.3 | −7.2 | 7.1 | −13.1 | −6.3 | 6.8 | FAD-dependent oxidoreductase |
| AFLA_139480 | 3 | −12.4 | −6.4 | 6.0 | −12.7 | −5.5 | 7.2 | Tryptophan synthase |
| AFLA_139490 | 3 | −11.9 | −6.7 | 5.2 | −9.9 | −7.0 | - | hybrid PKS/NRPS enzyme |
| 30 h 1 | 72 h | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Gene ID 2 | Chr 3 | Non vs. Tox | Co vs. Tox | Co vs. Non | Non vs. Tox | Co vs. Tox | Co vs. Non | SM 4 | Putative Function |
| (a) Genes with at least 8 log2-fold differential expression | |||||||||
| AFLA_085640 | 1 | 8.4 | 8.0 | - | 4.7 | 4.8 | - | -, - | Peroxisome biogenesis |
| AFLA_025220 | 2 | 11.9 | 11.4 | - | 7.8 | 7.8 | - | -, - | UPF0047 protein family |
| AFLA_126420 | 2 | 7.9 | 8.5 | - | 9.6 | 9.4 | - | -, - | Protein glycosylation |
| AFLA_060320 | 5 | 8.6 | 8.4 | - | 7.3 | 7.1 | - | -, - | Perforin domain |
| AFLA_060350 | 5 | 9.0 | 8.5 | - | 6.1 | 6.5 | - | -, - | Unknown |
| AFLA_062960 | 5 | 10.1 | 10.2 | - | 10.3 | 10.2 | - | -, 20 | Zn(2)-C6 transcript. factor |
| AFLA_062980 | 5 | 9.8 | 10.0 | - | 9.8 | 9.7 | - | -, 20 | Crotonase activity |
| AFLA_062990 | 5 | 8.7 | 8.7 | - | 8.4 | 8.3 | - | -, 20 | Oxidase |
| AFLA_095290 | 5 | 9.5 | 9.6 | - | 2.3 | 2.9 | - | -, - | Unknown |
| AFLA_095300 | 5 | 8.4 | 8.3 | - | 2.6 | 3.0 | - | -, - | Protein-protein interactions |
| AFLA_095800 | 5 | 7.9 | 8.0 | - | 6.1 | 5.7 | - | -, - | Short-chain reductase |
| AFLA_066370 | 6 | 8.9 | 8.7 | - | 8.7 | 8.8 | - | -, - | Phosphorylation |
| AFLA_008080 | 8 | 10.4 | 9.6 | - | - | - | - | -, - | Unknown |
| AFLA_117340 | 8 | 6.4 | 8.0 | - | 7.9 | - | - | -, - | 2-methylcitrate catabolism |
| (b) Differential gene expression in SMURF-predicted secondary metabolite cluster 20 (asperfuranone) | |||||||||
| AFLA_062800 | 5 | 2.4 | 2.4 | - | 1.9 | 1.9 | - | -, 20 | Aldo-keto reductase |
| AFLA_062810 | 5 | 3.6 | 3.6 | - | 5.1 | 4.9 | - | -, 20 | Hypothetical protein |
| AFLA_062820 | 5 | 3.3 | 4.5 | 1.2 | 5.3 | 5.0 | - | -, 20 | Polyketide synthase |
| AFLA_062830 | 5 | 3.8 | 4.2 | - | 5.0 | 4.8 | - | -, 20 | Monoxy./oxidoreductase |
| AFLA_062840 | 5 | 2.2 | 2.3 | - | 2.9 | 2.7 | - | -, 20 | Serine hydrolase |
| AFLA_062850 | 5 | 3.2 | 2.9 | - | - | - | - | -, 20 | Fatty acid oxidoreductase |
| AFLA_062860 | 5 | 3.5 | 4.4 | - | 5.4 | 5.1 | - | -, 20 | Polyketide synthase |
| AFLA_062890 | 5 | 1.2 | - | - | 1.6 | 1.2 | - | -, 20 | Hypothetical protein |
| AFLA_062940 | 5 | 7.4 | 7.5 | - | 6.6 | 6.6 | - | -, 20 | Choline transport protein |
| AFLA_062950 | 5 | 4.8 | 4.8 | - | 5.1 | 4.8 | - | -, 20 | Hypothetical protein |
| AFLA_062960 | 5 | 10.1 | 10.2 | - | 10.3 | 10.2 | - | -, 20 | Zn(2)-C6 transcript. factor |
| AFLA_062970 | 5 | 3.8 | 4.9 | - | 4.9 | 5.0 | - | -, 20 | Copper oxidase |
| AFLA_062980 | 5 | 9.8 | 10.0 | - | 9.8 | 9.7 | - | -, 20 | Crotonase activity |
| AFLA_062990 | 5 | 8.7 | 8.7 | - | 8.4 | 8.3 | - | -, 20 | Iron-binding oxidase |
| AFLA_063000 | 5 | 5.4 | 5.3 | - | 6.2 | 5.7 | - | -, 20 | Metal-binding hydrolase |
| ALFA_063020 | 5 | 4.3 | 5.1 | - | 5.4 | 6.0 | - | -, 20 | Multidrug resistance pump |
| 30 h 1 | 72 h | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene ID 2 | Chr 3 | Tox 53 | Non-Tox 17 | Co-Culture | Tox 53 | Non-Tox 17 | Co-Culture | SM 4 | Putative Function | ||||||||||||
| (a) Genes which were upregulated in Co-cultures compared to Tox 53 and Non-tox 17 | |||||||||||||||||||||
| 029700 | 2 | 3 | ±0.4 | * | 10 | ±2 | ** | 8 | ±0.7 | ** | 20 | ±2.2 | * | 27 | ±4 | * | 140 | ±6.8 | ** | 0, 0 | Peptidase |
| 037820 | 2 | 75 | ±22 | * | 105 | ±37 | ** | 78 | ±27 | * | 17 | ±2.2 | * | 51 | ±9.7 | ** | 167 | ±3.6 | *** | 0, 0 | Hsp20 heat shock protein |
| 124980 | 2 | 2 | ±0.1 | * | 3 | ±1.2 | * | 2 | ±0.3 | * | 6 | ±0.9 | * | 31 | ±12 | ** | 88 | ±16 | *** | 0, 0 | Oxidoreduction decarboxylase |
| 125000 | 2 | 0 | ±0 | * | 0 | ±0.1 | * | 0 | ±0.1 | * | 0 | ±0 | * | 6 | ±2.2 | ** | 22 | ±4.6 | *** | 0, 0 | Membrane transport facilitator |
| 126260 | 2 | 5 | ±0.5 | ** | 1 | ±0.3 | * | 1 | ±0.5 | * | 1 | ±0.2 | * | 2 | ±0.3 | ** | 9 | ±0.7 | *** | 0, 0 | FAD-dependent oxidoreductase |
| 126390 | 2 | 10 | ±1 | * | 30 | ±3.2 | *** | 18 | ±2.4 | ** | 35 | ±0.5 | * | 74 | ±17 | ** | 281 | ±24 | *** | 0, 0 | Metal-binding monoxygenase |
| 135320 | 3 | 10 | ±1.3 | * | 50 | ±2.1 | ** | 70 | ±3.3 | *** | 55 | ±2.6 | *** | 15 | ±0.8 | * | 33 | ±2.1 | ** | 0, 0 | Membrane transport facilitator |
| 000840 | 4 | 1 | ±0.1 | * | 4 | ±0.4 | ** | 7 | ±1.7 | *** | 2 | ±0 | * | 27 | ±1.8 | ** | 88 | ±35 | *** | 0, 0 | Membrane transport facilitator |
| 000870 | 4 | 19 | ±3.2 | * | 57 | ±13 | ** | 27 | ±3.1 | * | 2 | ±0.2 | * | 11 | ±1.1 | ** | 26 | ±3.8 | *** | 0, 0 | Hypothetical protein |
| 001010 | 4 | 0 | ±0 | * | 58 | ±2.5 | *** | 56 | ±4.1 | ** | 41 | ±2.5 | ** | 28 | ±3.6 | * | 83 | ±2.5 | *** | 0, 0 | Cyt P450 oxygenase |
| 013270 | 4 | 0 | ±0.1 | * | 1 | ±0.2 | * | 0 | ±0.1 | * | 1 | ±0.1 | * | 18 | ±8.9 | ** | 80 | ±13 | *** | 0, 0 | 4-oxalocrotonate tautomerase |
| 013680 | 4 | 0 | ±0.1 | * | 1 | ±0.3 | ** | 1 | ±0.1 | ** | 1 | ±0.1 | * | 6 | ±0.4 | ** | 12 | ±1.2 | *** | 0, 0 | Phospholipase C |
| 016350 | 4 | 0 | ±0.1 | * | 0 | ±0.1 | * | 2 | ±1.6 | ** | 0 | ±0.1 | * | 13 | ±1.6 | ** | 23 | ±5.7 | ** | 0, 0 | NAD(P)H-dependent reductase |
| 059120 | 5 | 0 | ±0 | * | 0 | ±0 | * | 0 | ±0 | * | 1 | ±0.1 | * | 1 | ±0.1 | * | 3 | ±0.5 | ** | 0, 0 | Metal binding fumarylacetoacetase |
| 060770 | 5 | 0 | ±0.1 | * | 1 | ±0.1 | ** | 1 | ±0.4 | ** | 2 | ±0.1 | * | 26 | ±1.8 | ** | 47 | ±3.1 | *** | 0, 0 | Acetate transport/EtOH synthesis |
| 061090 | 5 | 0 | ±0.1 | * | 6 | ±0.4 | *** | 4 | ±0.1 | ** | 3 | ±0 | * | 2 | ±0.2 | * | 5 | ±0.5 | ** | 0, 0 | Serine/threonine MAP-kinase |
| 091690 | 5 | 16 | ±0.9 | * | 24 | ±0.8 | *** | 20 | ±0.5 | ** | 23 | ±0.2 | * | 42 | ±3.7 | ** | 94 | ±8.4 | *** | 0, 0 | Isocitrate lyase |
| 096040 | 5 | 3 | ±0.2 | * | 73 | ±3.7 | ** | 68 | ±8.6 | ** | 11 | ±1.6 | * | 632 | ±110 | ** | 1589 | ±530 | *** | 0, 0 * | FAD-dependent oxidoreductase |
| 096060 | 5 | 0 | ±0 | * | 1 | ±0.1 | ** | 1 | ±0.1 | ** | 2 | ±0.2 | * | 21 | ±5 | ** | 95 | ±31 | *** | 0, 0 * | Membrane transport facilitator |
| 097430 | 6 | 17 | ±4.1 | * | 14 | ±0.4 | * | 11 | ±1.4 | * | 4 | ±0.6 | * | 11 | ±1.2 | ** | 70 | ±32 | *** | 0, 0 | NAD-dependent dehydratase |
| 097440 | 6 | 9 | ±0.8 | * | 10 | ±0.3 | * | 7 | ±0.9 | * | 3 | ±0.2 | * | 6 | ±1.3 | ** | 32 | ±13 | *** | 0, 0 | Unknown-NAD(P) binding |
| 040120 | 7 | 100 | ±13 | * | 302 | ±10 | *** | 314 | ±23 | ** | 352 | ±11 | * | 313 | ±22 | * | 665 | ±21 | ** | 0, 0 | FAD-binding oxidoreductase |
| 117330 | 8 | 5 | ±0.3 | * | 94 | ±15 | ** | 103 | ±5.6 | *** | 24 | ±3.9 | * | 23 | ±3.4 | * | 71 | ±9.5 | ** | 0, 0 | Membrane transport facilitator |
| 118940 | 8 | 0 | ±0 | * | 0 | ±0 | * | 0 | ±0 | * | 0 | ±0 | * | 3 | ±0.9 | ** | 5 | ±2.7 | *** | 8.5, 46 | Polyketide synthase |
| 118960 | 8 | 0 | ±0 | * | 0 | ±0 | * | 0 | ±0 | * | 0 | ±0 | * | 2 | ±0.6 | ** | 5 | ±3.2 | *** | 8.5, 46 | Polyketide synthase |
| 118970 | 8 | 0 | ±0.1 | * | 0 | ±0 | * | 0 | ±0 | * | 0 | ±0.1 | * | 4 | ±1.3 | ** | 11 | ±5.6 | *** | 8.5, 46 | FAD-dependent oxygenase |
| 118990 | 8 | 0 | ±0 | * | 0 | ±0 | * | 0 | ±0 | * | 0 | ±0 | * | 14 | ±4.3 | ** | 46 | ±25 | *** | 8.5, 0 | Efflux pump, major facilitator |
| 119000 | 8 | 0 | ±0 | * | 0 | ±0 | * | 0 | ±0 | * | 0 | ±0 | * | 65 | ±19 | ** | 198 | ±120 | *** | 8.5, 0 | O-methyl transferase |
| 122110 | 8 | 5 | ±0.2 | * | 2 | ±0.3 | * | 3 | ±0.6 | * | 3 | ±0.7 | * | 30 | ±5.1 | ** | 136 | ±54 | *** | 0, 0 | Haem bifunctional peroxidase |
| (b) Genes which were highly overexpressed in Non-tox 17 compared to Tox 53 | |||||||||||||||||||||
| 085640 | 1 | 0 | ±0 | * | 6 | ±0.3 | ** | 5 | ±0.7 | ** | 0 | ±0 | * | 5 | ±0.8 | ** | 5 | ±1.2 | ** | 0, 0 | Peroxisome biogenesis |
| 025220 | 2 | 0 | ±0 | * | 109 | ±7.5 | ** | 81 | ±5.3 | ** | 0 | ±0.4 | * | 103 | ±3.9 | ** | 104 | ±7 | ** | 0, 0 | UPF0047 protein family |
| 126420 | 2 | 1 | ±0.3 | * | 3 | ±0.2 | ** | 5 | ±0.2 | ** | 0 | ±0 | * | 5 | ±0.2 | ** | 4 | ±0.2 | ** | 0, 0 | Protein glycosylation |
| 060320 | 5 | 0 | ±0 | * | 5 | ±0.9 | ** | 5 | ±0.9 | ** | 0 | ±0 | * | 7 | ±0.6 | ** | 6 | ±0.4 | ** | 0, 0 | Perforin domain |
| 060350 | 5 | 0 | ±0 | * | 12 | ±1.6 | ** | 8 | ±2 | ** | 0 | ±0 | * | 2 | ±0.4 | ** | 3 | ±0.5 | ** | 0, 0 | Unknown |
| 062960 | 5 | 0 | ±0 | * | 7 | ±0.1 | ** | 8 | ±0.1 | ** | 0 | ±0 | * | 14 | ±0.8 | ** | 13 | ±0.5 | ** | 0, 20 | Zn(2)-C6 transcription factor |
| 062980 | 5 | 0 | ±0 | * | 12 | ±1.3 | ** | 15 | ±0.5 | ** | 0 | ±0 | * | 40 | ±2.2 | ** | 36 | ±0.7 | ** | 0, 20 | Crotonase activity |
| 062990 | 5 | 0 | ±0 | * | 5 | ±0.2 | ** | 5 | ±0.1 | ** | 0 | ±0 | * | 12 | ±0.3 | ** | 10 | ±0.2 | ** | 0, 20 | Oxidase |
| 095290 | 5 | 0 | ±0.3 | * | 90 | ±3.6 | ** | 103 | ±4.3 | ** | 1 | ±0.3 | * | 8 | ±0.5 | ** | 13 | ±1.2 | ** | 0, 0 | Unknown |
| 095300 | 5 | 5 | ±0.3 | * | 102 | ±9.4 | ** | 97 | ±6.6 | ** | 1 | ±0.2 | * | 6 | ±0.1 | ** | 8 | ±1.1 | ** | 0, 0 | Protein-protein interactions |
| 095800 | 5 | 4 | ±1.5 | * | 71 | ±3.1 | ** | 80 | ±3.6 | ** | 1 | ±0.2 | * | 62 | ±2.8 | ** | 48 | ±0.4 | ** | 0, 0 | Short-chain reductase |
| 066370 | 6 | 0 | ±0 | * | 12 | ±0.3 | ** | 11 | ±0.8 | ** | 0 | ±0 | * | 33 | ±1.9 | ** | 34 | ±1.2 | ** | 0, 0 | Phosphorylation |
| 008080 | 8 | 1 | ±0.3 | * | 216 | ±23 | ** | 128 | ±14 | ** | 65 | ±2.7 | * | 112 | ±4.6 | ** | 128 | ±1.1 | ** | 0, 0 | Unknown |
| 117340 | 8 | 0 | ±0 | * | 1 | ±0.2 | ** | 3 | ±2.1 | ** | 0 | ±0 | * | 4 | ±3.6 | ** | 0 | ±0 | ** | 0, 0 | 2-methylcitrate catabolism |
| 30 h 1 | 72 h | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene ID 2 | Chr 3 | Tox 53 | Non-Tox 17 | Co-Culture | Tox 53 | Non-Tox 17 | Co-Culture | SM 4 | ||||||||||||
| 064230 | 6 | 30 | ±3 | * | 33 | ±0.9 | * | 21 | ±1.2 | * | 10 | ±1.6 | * | 209 | ±12.3 | ** | 134 | ±4.7 | ** | 0, 21 |
| 064240 | 6 | 6 | ±0.8 | * | 9 | ±0.7 | * | 7 | ±0.3 | * | 4 | ±0.3 | * | 68 | ±2 | ** | 41 | ±1.4 | ** | 0, 21 |
| 064250 | 6 | 31 | ±8.6 | * | 168 | ±3.9 | * | 79 | ±12.2 | * | 13 | ±0.9 | * | 613 | ±29.2 | * | 333 | ±6.2 | * | 0, 21 |
| 064260 | 6 | 4 | ±0.9 | * | 15 | ±1.3 | * | 9 | ±1.4 | * | 10 | ±0.5 | * | 153 | ±4.2 | * | 140 | ±1 | * | 0, 21 |
| 064270 | 6 | 37 | ±9.7 | * | 278 | ±9.2 | * | 141 | ±23.4 | * | 15 | ±1.1 | * | 1132 | ±36.9 | * | 711 | ±17.5 | * | 0, 21 |
| 064280 | 6 | 24 | ±7.3 | * | 150 | ±10.9 | * | 66 | ±10.2 | * | 6 | ±0.2 | * | 610 | ±24.1 | * | 290 | ±12.3 | * | 0, 21 |
| 064290 | 6 | 355 | ±90.6 | ** | 615 | ±23.8 | ** | 291 | ±42.5 | * | 43 | ±1.7 | * | 1393 | ±57.3 | ** | 762 | ±31.9 | ** | 0, 21 |
| 064300 | 6 | 15 | ±2.1 | * | 42 | ±0.8 | * | 20 | ±2.8 | * | 6 | ±0.4 | * | 146 | ±5.5 | * | 93 | ±1.2 | * | 0, 21 |
| 064310 | 6 | 5 | ±0.5 | * | 14 | ±0.1 | * | 8 | ±0.9 | * | 5 | ±0.3 | * | 59 | ±3.5 | * | 44 | ±1.2 | * | 0, 21 |
| 064320 | 6 | 18 | ±3.8 | * | 134 | ±7.8 | * | 60 | ±9.4 | * | 8 | ±0.6 | * | 443 | ±20.8 | * | 250 | ±4.7 | * | 0, 21 |
| 064330 | 6 | 7 | ±0.3 | * | 12 | ±1 | * | 7 | ±1.2 | * | 4 | ±0.3 | * | 23 | ±2.1 | ** | 17 | ±1.5 | ** | 0, 21 |
| 064340 | 6 | 6 | ±0.9 | * | 19 | ±0.3 | * | 9 | ±1.4 | * | 2 | ±0.3 | * | 22 | ±0.1 | * | 16 | ±1.5 | * | 0, 21 |
| 064350 | 6 | 0 | ±0.1 | * | 0 | ±0 | * | 0 | ±0 | * | 1 | ±0 | * | 1 | ±0.1 | * | 1 | ±0 | * | 0, 21 |
| 064360 | 6 | 0 | ±0.1 | * | 0 | ±0 | * | 0 | ±0.1 | * | 1 | ±0.1 | * | 1 | ±0 | * | 1 | ±0 | * | 0, 21 |
| 064370 | 6 | 1 | ±0.2 | * | 2 | ±0.1 | * | 1 | ±0.2 | * | 2 | ±0.1 | * | 4 | ±0.2 | * | 3 | ±0.2 | * | 0, 21 |
| 064380 | 6 | 185 | ±15.9 | * | 36 | ±2.3 | * | 63 | ±9.3 | * | 5 | ±0.3 | * | 85 | ±3.2 | * | 47 | ±1.8 | * | 0, 21 |
| 064390 | 6 | 11 | ±0.7 | * | 3 | ±0.4 | * | 5 | ±0.5 | * | 0 | ±0.1 | * | 7 | ±0.1 | * | 4 | ±0.2 | * | 0, 21 |
| 064400 | 6 | 32 | ±4.2 | * | 11 | ±0.5 | * | 18 | ±3.3 | * | 1 | ±0.2 | * | 21 | ±1.3 | * | 11 | ±0.4 | * | 0, 21 |
| 064410 | 6 | 9 | ±1.1 | * | 4 | ±0.7 | * | 5 | ±0.8 | * | 1 | ±0.1 | * | 8 | ±0.9 | ** | 6 | ±0.4 | ** | 0, 21 |
| 064420 | 6 | 112 | ±7.7 | * | 18 | ±0.4 | * | 22 | ±4.3 | * | 4 | ±0.6 | * | 46 | ±3.6 | * | 25 | ±1.5 | * | 0, 21 |
| 064430 | 6 | 89 | ±5.1 | * | 17 | ±1 | * | 22 | ±0.1 | * | 9 | ±0.8 | * | 38 | ±1.7 | * | 26 | ±1 | * | 0, 21 |
| 064440 | 6 | 35 | ±3 | * | 10 | ±0.7 | * | 16 | ±2.1 | * | 1 | ±0.1 | * | 22 | ±1.8 | * | 10 | ±0.8 | * | 0, 21 |
| 064450 | 6 | 44 | ±2.2 | * | 13 | ±1.3 | * | 21 | ±3.2 | * | 1 | ±0.2 | * | 33 | ±1.6 | * | 17 | ±0.8 | * | 0, 21 |
| 064460 | 6 | 16 | ±2 | * | 4 | ±0.6 | * | 8 | ±0.9 | * | 0 | ±0.1 | * | 9 | ±0.8 | * | 4 | ±0.4 | * | 1.1, 21 |
| 064470 | 6 | 32 | ±3.2 | * | 14 | ±0.9 | * | 20 | ±3 | * | 1 | ±0.2 | * | 20 | ±1.7 | * | 10 | ±0.1 | * | 1.1, 21 |
| 064480 | 6 | 137 | ±14.1 | * | 68 | ±7.1 | * | 81 | ±15.4 | * | 5 | ±0.2 | * | 79 | ±3.9 | ** | 40 | ±2.9 | ** | 1.1, 21 |
| 064490 | 6 | 90 | ±7.8 | * | 32 | ±1.2 | * | 46 | ±8.9 | * | 7 | ±0.4 | * | 60 | ±0.7 | ** | 30 | ±1.4 | ** | 1.1, 21 |
| 064500 | 6 | 727 | ±33.2 | ** | 340 | ±28.1 | * | 309 | ±31.9 | * | 179 | ±5.1 | * | 996 | ±19.5 | ** | 717 | ±16.8 | ** | 1.1, 21 |
| 064510 | 6 | 40 | ±3.1 | * | 23 | ±0.8 | * | 34 | ±5.4 | * | 8 | ±0.5 | * | 32 | ±1.2 | ** | 20 | ±0.1 | ** | 1.1, 21 |
| 064520 | 6 | 40 | ±2.4 | * | 19 | ±1.6 | * | 29 | ±4.5 | * | 9 | ±0.4 | * | 33 | ±0.8 | ** | 23 | ±0.5 | ** | 1.1, 21 |
| 064530 | 6 | 64 | ±9.2 | * | 22 | ±2.7 | * | 29 | ±5.5 | * | 1 | ±0.1 | * | 34 | ±1.7 | * | 18 | ±0.8 | * | 1.1, 21 |
| 064540 | 6 | 24 | ±2.2 | * | 8 | ±0.9 | * | 11 | ±1.8 | * | 1 | ±0.1 | * | 17 | ±1.1 | * | 8 | ±0.4 | * | 1.1, 21 |
| 064550 | 6 | 22 | ±2.6 | * | 8 | ±0.9 | * | 14 | ±2.3 | * | 1 | ±0.1 | * | 20 | ±1.3 | * | 11 | ±0.3 | * | 1.1, 21 |
| 064560 | 6 | 6 | ±0.4 | * | 2 | ±0.1 | * | 5 | ±0.6 | * | 0 | ±0 | * | 10 | ±0.2 | * | 5 | ±0.1 | * | 1.1, 21 |
| 064570 | 6 | 56 | ±10.4 | * | 8 | ±0.5 | * | 12 | ±0.4 | * | 1 | ±0.2 | * | 16 | ±0.4 | * | 8 | ±0.2 | * | 1.1, 21 |
| 064580 | 6 | 122 | ±13.4 | * | 42 | ±4.6 | * | 53 | ±9.3 | * | 2 | ±0.4 | * | 67 | ±3.1 | * | 33 | ±2.1 | * | 1.1, 21 |
| 064590 | 6 | 122 | ±13.3 | * | 22 | ±1.5 | * | 32 | ±3 | * | 1 | ±0.1 | * | 42 | ±2.3 | * | 23 | ±0.6 | * | 1.1, 21 |
| 064600 | 6 | 7 | ±0.7 | * | 1 | ±0.3 | * | 2 | ±0.2 | * | 0 | ±0.1 | * | 4 | ±0.1 | * | 2 | ±0 | * | 1.1, 21 |
| 064610 | 6 | 2 | ±0.4 | * | 1 | ±0.2 | * | 1 | ±0.1 | * | 2 | ±0.1 | * | 7 | ±0.2 | ** | 9 | ±2.8 | ** | 1.1, 21 |
| Cultured Isolates | Chemicals Extracted | Hours | Biological Replicates | Dishes per Rep |
|---|---|---|---|---|
| Non-tox 17 | RNA and aflatoxin | 30 | 5 | 9 |
| Tox 53 | RNA and aflatoxin | 30 | 5 | 9 |
| Co-culture of 17 + 53 | RNA and aflatoxin | 30 | 5 | 9 |
| Non-tox 17 | RNA and aflatoxin | 72 | 4 | 1 |
| Tox 53 | RNA and aflatoxin | 72 | 4 | 1 |
| Co-culture of 17 + 53 | RNA and aflatoxin | 72 | 4 | 1 |
| Non-tox 17 | Aflatoxin | 96 | 4 | 1 |
| Tox 53 | Aflatoxin | 96 | 4 | 1 |
| Co-culture of 17 + 53 | Aflatoxin | 96 | 4 | 1 |
| Total samples | 39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sweany, R.R.; Mack, B.M.; Moore, G.G.; Gilbert, M.K.; Cary, J.W.; Lebar, M.D.; Rajasekaran, K.; Damann, K.E., Jr. Genetic Responses and Aflatoxin Inhibition during Co-Culture of Aflatoxigenic and Non-Aflatoxigenic Aspergillus flavus. Toxins 2021, 13, 794. https://doi.org/10.3390/toxins13110794
Sweany RR, Mack BM, Moore GG, Gilbert MK, Cary JW, Lebar MD, Rajasekaran K, Damann KE Jr. Genetic Responses and Aflatoxin Inhibition during Co-Culture of Aflatoxigenic and Non-Aflatoxigenic Aspergillus flavus. Toxins. 2021; 13(11):794. https://doi.org/10.3390/toxins13110794
Chicago/Turabian StyleSweany, Rebecca R., Brian M. Mack, Geromy G. Moore, Matthew K. Gilbert, Jeffrey W. Cary, Matthew D. Lebar, Kanniah Rajasekaran, and Kenneth E. Damann, Jr. 2021. "Genetic Responses and Aflatoxin Inhibition during Co-Culture of Aflatoxigenic and Non-Aflatoxigenic Aspergillus flavus" Toxins 13, no. 11: 794. https://doi.org/10.3390/toxins13110794
APA StyleSweany, R. R., Mack, B. M., Moore, G. G., Gilbert, M. K., Cary, J. W., Lebar, M. D., Rajasekaran, K., & Damann, K. E., Jr. (2021). Genetic Responses and Aflatoxin Inhibition during Co-Culture of Aflatoxigenic and Non-Aflatoxigenic Aspergillus flavus. Toxins, 13(11), 794. https://doi.org/10.3390/toxins13110794