The Potential of Peroxidases Extracted from the Spent Mushroom (Flammulina velutipes) Substrate Significantly Degrade Mycotoxin Deoxynivalenol

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Effects of MnP and LiP on Growth Inhibition of F. graminearum KR1
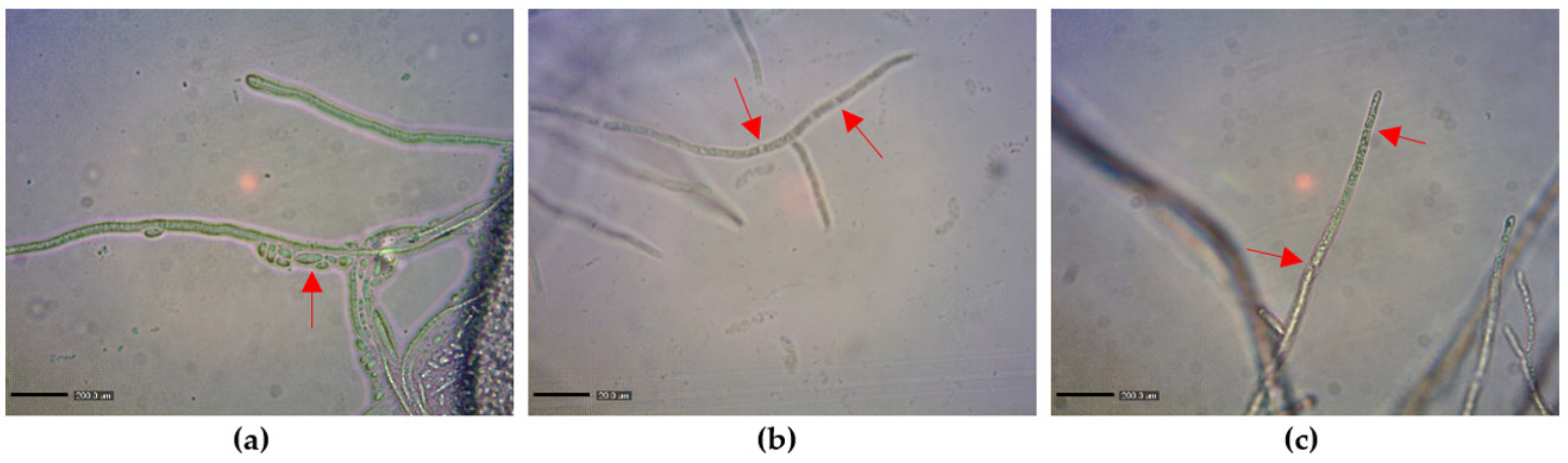
2.2. Influence of MnP and LiP on Cell Wall Hydrolysis and Morphology
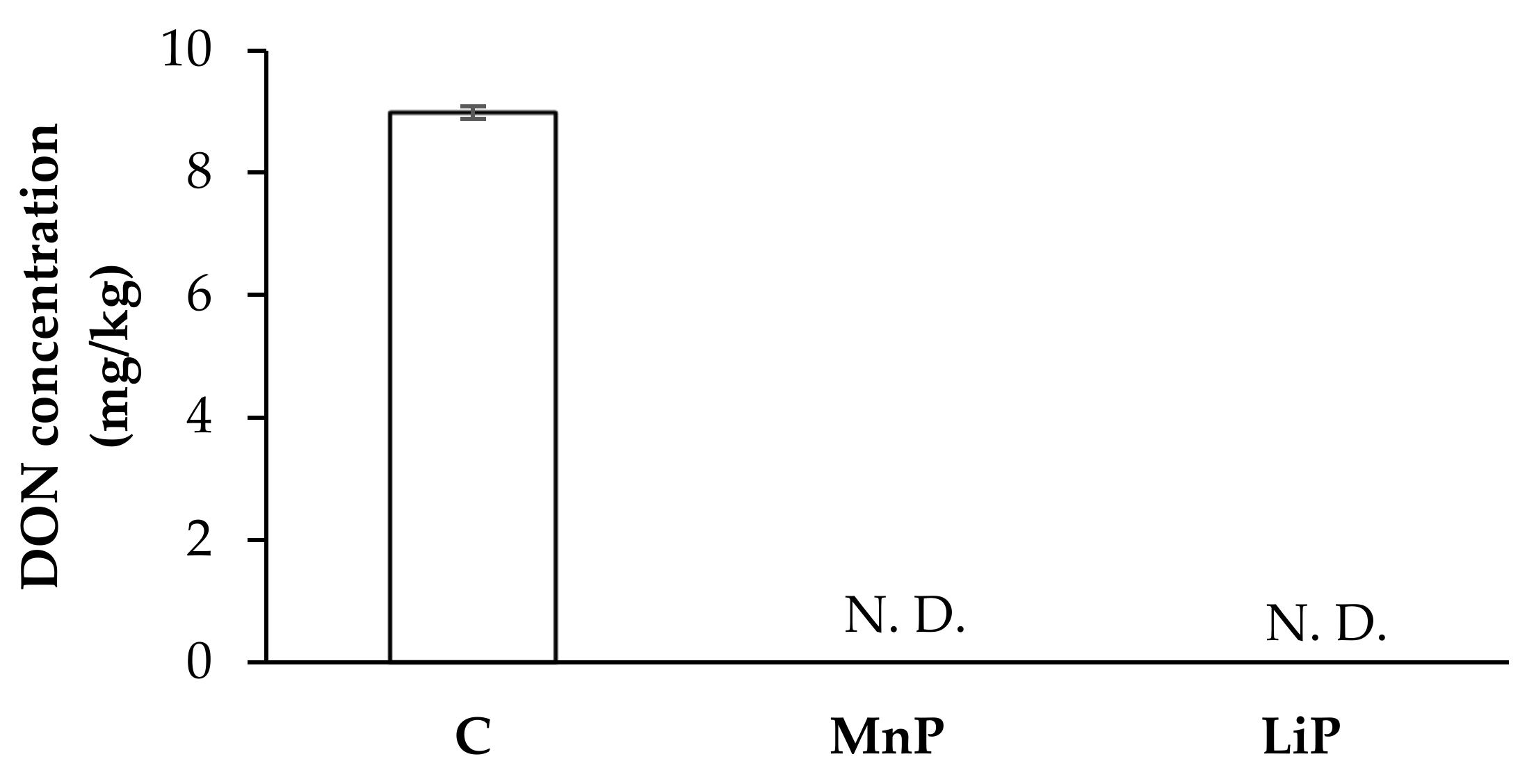
2.3. Effects of MnP and LiP on DON Production of F. graminearum KR1
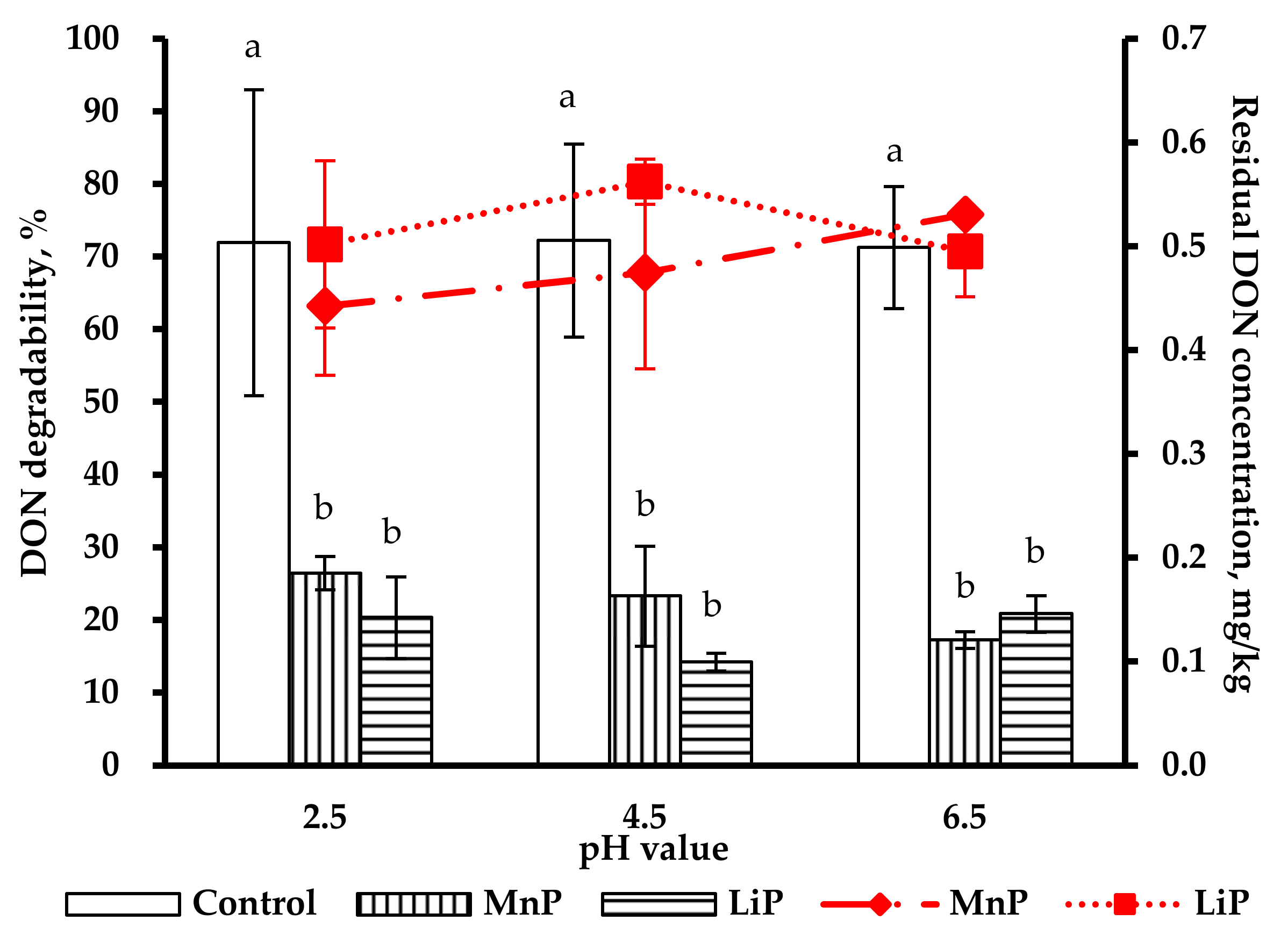
2.4. Pre-Test of DON Degradability by MnP and LiP in Artificial Digestive Juices
2.5. Simulation of Pig Gastrointestinal Tracts in DON Degradability by LiP Treatment
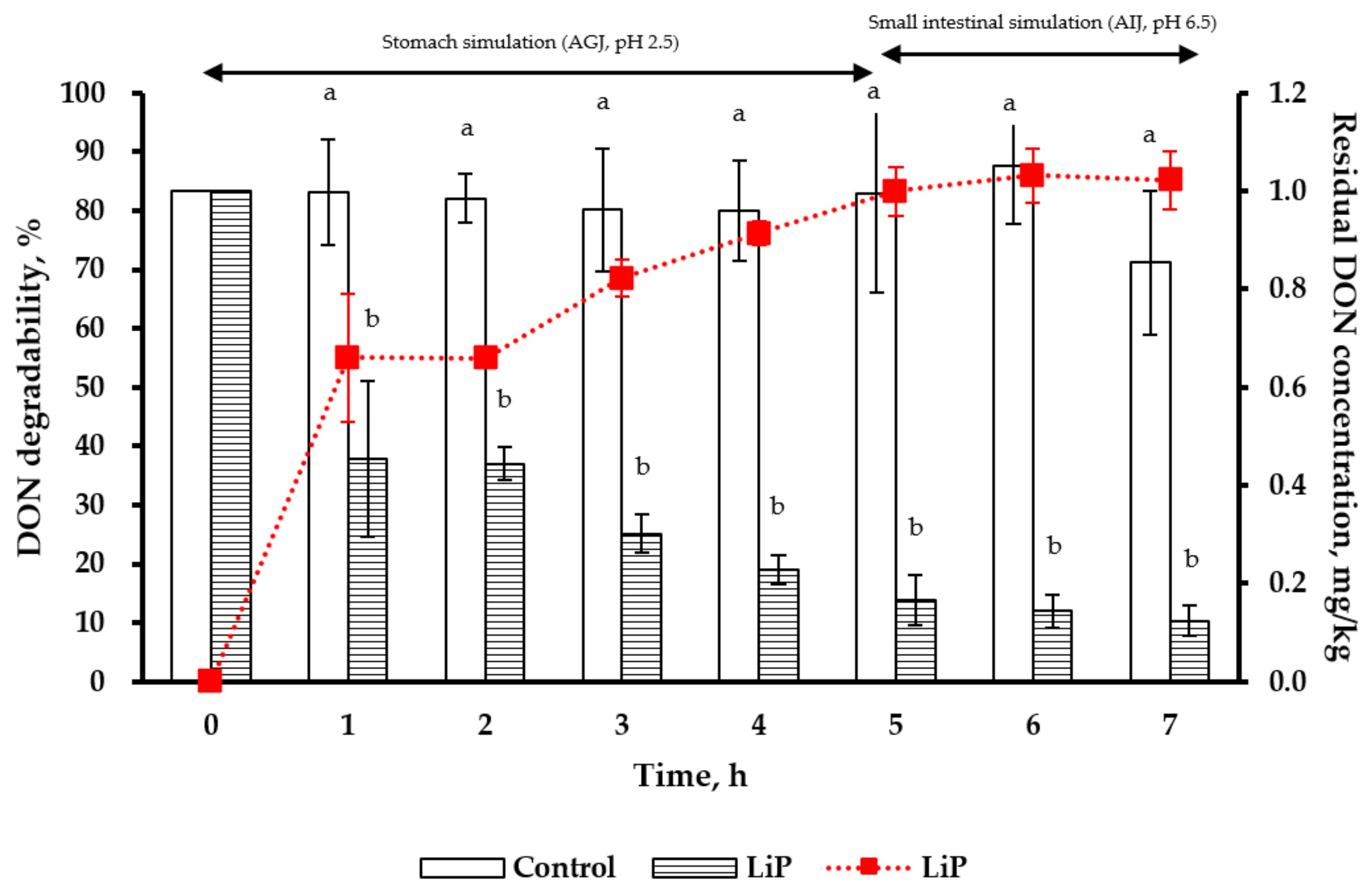
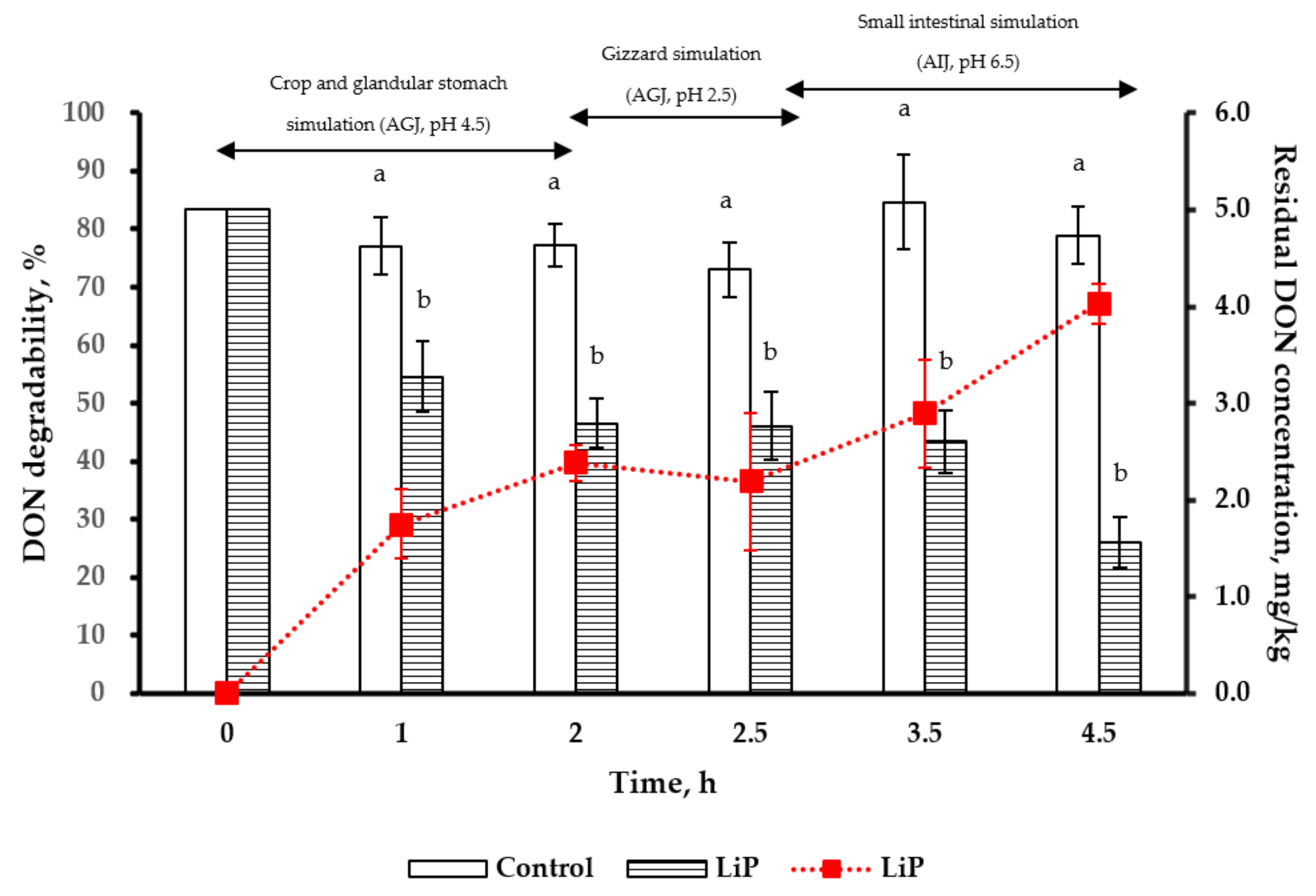
2.6. Simulation of Poultry Gastrointestinal Tracts in DON Degradability by LiP Treatment
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Fungal Strain
5.2. Extraction and Analyses of MnP and LiP
5.3. Antifungal Activity Assays
5.4. Determination of GlcNAc Content, Chitinase Activity, and Hyphae Morphology
5.5. Preparation of Artificial Digestive Juices
5.6. A Pre-Test for DON Degradability of MnP and LiP
5.7. Simulations of Pig and Poultry Gastrointestinal Tracts with LiP Treatment
5.8. DON Extraction and Assay
5.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, M.C.; Madec, S.; Coton, E.; Hymery, N. Natural co-occurrence of mycotoxins in foods and feeds and their in vitro combined toxicological effects. Toxins (Basel) 2016, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajslova, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Z.; Beier, R.C.; Shen, J.; De Smet, D.; De Saeger, S.; Zhang, S. T-2 toxin, a trichothecene mycotoxin: Review of toxicity, metabolism, and analytical methods. J. Agric. Food Chem. 2011, 59, 3441–3453. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, D.M.; Osborne, S.; Kazan, K.; Manners, J.M. Low pH regulates the production of deoxynivalenol by Fusarium graminearum. Microbiol. Sgm. 2009, 155, 3149–3156. [Google Scholar] [CrossRef]
- Magan, N.; Aldred, D.; Hope, R.; Mitchell, D. Environmental factors and interactions with mycobiota of grain and grapes: Effects on growth, deoxynivalenol and ochratoxin production by Fusarium culmorum and Aspergillus carbonarius. Toxins 2010, 2, 353–366. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Parra, R.; Geisen, R.; Magan, N. Modelling the relationship between environmental factors, transcriptional genes and deoxynivalenol mycotoxin production by strains of two Fusarium species. J. R. Soc. Interface 2011, 8, 117–126. [Google Scholar] [CrossRef]
- Moretti, A.; Pascale, M.; Logrieco, A.F. Mycotoxin risks under a climate change scenario in Europe. Trends Food Sci. Technol. 2019, 84, 38–40. [Google Scholar] [CrossRef]
- Rocha, O.; Ansari, K.; Doohan, F.M. Effects of trichothecene mycotoxins on eukaryotic cells: A review. Food Addit. Contam. 2005, 22, 369–378. [Google Scholar] [CrossRef]
- Springler, A.; Hessenberger, S.; Reisinger, N.; Kern, C.; Nagl, V.; Schatzmayr, G.; Mayer, E. Deoxynivalenol and its metabolite deepoxy-deoxynivalenol: Multi-parameter analysis for the evaluation of cytotoxicity and cellular effects. Mycotoxin Res. 2017, 33, 25–37. [Google Scholar] [CrossRef]
- Awad, W.A.; Ghareeb, K.; Dadak, A.; Hess, M.; Bohm, J. Single and combined effects of deoxynivalenol mycotoxin and a microbial feed additive on lymphocyte DNA damage and oxidative stress in broiler chickens. PLoS ONE 2014, 9, e88028. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No 1126/2007, on setting maximum levels for certain contaminants in foodstuffs as regards Fusarium toxins in maize and maize products. Off. J. Eur. Union 2007, 255, 14–17. [Google Scholar]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in occurrence, importance, and mycotoxin control strategies: Prevention and detoxification in foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Bretz, M.; Beyer, M.; Cramer, B.; Knecht, A.; Humpf, H.U. Thermal degradation of the Fusarium mycotoxin deoxynivalenol. J. Agric. Food Chem. 2006, 54, 6445–6451. [Google Scholar] [CrossRef]
- Doll, S.; Danicke, S. In vivo detoxification of Fusarium toxins. Arch. Anim. Nutr. 2004, 58, 419–441. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.G.; Su, X.O.; Fan, X.; Wang, P.L.; Gao, Z.W.; Zhang, Y. Liquid chromatography-tandem mass spectrometry for determination of aflatoxin B1, deoxynivalenol and zearalenone in artificial porcine gastrointestinal digestive juice. Chin. J. Anal. Chem. 2015, 43, 1–6. [Google Scholar] [CrossRef]
- Kihal, A.; Rodriguez-Prado, M.; Godoy, C.; Cristofol, C.; Calsamiglia, S. In vitro assessment of the capacity of certain mycotoxin binders to adsorb some amino acids and water-soluble vitamins. J. Dairy Sci. 2020, 103, 3125–3132. [Google Scholar] [CrossRef]
- Tomasevic-Canovic, M.; Dakovic, A.; Markovic, V.; Radosavljevic-Mihajlovic, A.; Vukicevic, J. Adsorpton effects of mineral adsorbents; Part III: Adsorption behaviour in the presence of vitamin B6 and microelements. Acta. Vet. Beogr. 2000, 50, 23–29. [Google Scholar]
- European Commission. Commission Regulation (EC) No 466/2001 of 8 March 2001 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2001, 77, 1–13. [Google Scholar]
- Al-Rashood, K.A.; Abdel-Aziz, H.A. Thiazolo[3,2-a]benzimidazoles: Synthetic strategies, chemical transformations and biological activities. Molecules 2010, 15, 3775–3815. [Google Scholar] [CrossRef]
- Awad, W.A.; Ghareeb, K.; Bohm, J.; Zentek, J. Decontamination and detoxification strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the effectiveness of microbial biodegradation. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2010, 27, 510–520. [Google Scholar] [CrossRef]
- Young, J.C.; Zhou, T.; Yu, H.; Zhu, H.; Gong, J. Degradation of trichothecene mycotoxins by chicken intestinal microbes. Food Chem. Toxicol. 2007, 45, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Ikunaga, Y.; Sato, I.; Grond, S.; Numaziri, N.; Yoshida, S.; Yamaya, H.; Hiradate, S.; Hasegawa, M.; Toshima, H.; Koitabashi, M. Nocardioides sp. strain WSN05-2, isolated from a wheat field, degrades deoxynivalenol, producing the novel intermediate 3-epi-deoxynivalenol. Appl. Microbiol. Biotechnol. 2011, 89, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Sato, I.; Ishizaka, M.; Yoshida, S.; Koitabashi, M.; Yoshida, S.; Tsushima, S. Bacterial cytochrome P450 system catabolizing the Fusarium toxin deoxynivalenol. Appl. Environ. Microbiol. 2013, 79, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, I.; De Mets, L.; De Boevre, M.; Uka, V.; Di Mavungu, J.D.; De Saeger, S.; De Gelder, L.; Audenaert, K. Microbial Detoxification of deoxynivalenol (DON), assessed via a lemna minor L. bioassay, through biotransformation to 3-epi-DON and 3-epi-DOM-1. Toxins (Basel) 2017, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Binder, E.M.; Heidler, D.; Krska, R. Structural characterization of metabolites after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797. Food Addit. Contam. 2002, 19, 379–386. [Google Scholar] [CrossRef]
- Karlovsky, P. Biological detoxification of the mycotoxin deoxynivalenol and its use in genetically engineered crops and feed additives. Appl. Microbiol. Biotechnol. 2011, 91, 491–504. [Google Scholar] [CrossRef]
- Kumar, A.; Chandra, R. Ligninolytic enzymes and its mechanisms for degradation of lignocellulosic waste in environment. Heliyon 2020, 6, e03170. [Google Scholar] [CrossRef]
- Hofrichter, M. Review: Lignin conversion by manganese peroxidase (MnP). Enzyme Microb. Technol. 2002, 30, 454–466. [Google Scholar] [CrossRef]
- Wang, J.; Ogata, M.; Hirai, H.; Kawagishi, H. Detoxification of aflatoxin B1 by manganese peroxidase from the white-rot fungus Phanerochaete sordida YK-624. FEMS. Microbiol. Lett. 2011, 314, 164–169. [Google Scholar] [CrossRef]
- Wang, X.L.; Qin, X.; Hao, Z.Z.; Luo, H.Y.; Yao, B.; Su, X.Y. Degradation of four major mycotoxins by eight manganese peroxidases in presence of a dicarboxylic acid. Toxins 2019, 11, 566. [Google Scholar] [CrossRef]
- Garda-Buffon, J.; Kupski, L.; Badiale-Furlong, E. Deoxynivalenol (DON) degradation and peroxidase enzyme activity in submerged fermentation. Cienc. E. Tecnol. Alime. 2011, 31, 198–203. [Google Scholar] [CrossRef]
- Feltrin, A.C.P.; Garcia, S.O.; Caldas, S.S.; Primel, E.G.; Badiale-Furlong, E.; Garda-Buffon, J. Characterization and application of the enzyme peroxidase to the degradation of the mycotoxin DON. J. Environ. Sci. Health B 2017, 52, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Akyuz, M.; Kirbag, S. Antimicrobial activity of Pleurotus eryngii var. ferulae grown on various agro-wastes. Eurasia J. Biosci. 2009, 10, 58–63. [Google Scholar] [CrossRef]
- Schalchli, H.; Hormazabal, E.; Rubilar, O.; Briceno, G.; Mutis, A.; Zocolo, G.J.; Diez, M.C. Production of ligninolytic enzymes and some diffusible antifungal compounds by white-rot fungi using potato solid wastes as the sole nutrient source. J. Appl. Microbiol. 2017, 123, 886–895. [Google Scholar] [CrossRef]
- Schalchli, H.; Hormazabal, E.; Becerra, J.; Briceno, G.; Hernandez, V.; Rubilar, O.; Diez, M.C. Volatiles from white-rot fungi for controlling plant pathogenic fungi. Chem. Ecol. 2015, 31, 754–763. [Google Scholar] [CrossRef]
- Vinale, F.; Ghisalberti, E.L.; Flematti, G.; Marra, R.; Lorito, M.; Sivasithamparam, K. Secondary metabolites produced by a root-inhabiting sterile fungus antagonistic towards pathogenic fungi. Lett. Appl. Microbiol. 2010, 50, 380–385. [Google Scholar] [CrossRef]
- Beauvais, A.; Latge, J.P. Special issue: Fungal cell wall. J. Fungi (Basel) 2018, 4, 91. [Google Scholar] [CrossRef]
- Kim, J.E.; Lee, H.J.; Lee, J.; Kim, K.W.; Yun, S.H.; Shim, W.B.; Lee, Y.W. Gibberella zeae chitin synthase genes, GzCHS5 and GzCHS7, are required for hyphal growth, perithecia formation, and pathogenicity. Curr. Genet. 2009, 55, 449–459. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Chen, Q.; Liu, C.H.; Liu, Y.B.; Yi, P.; Niu, K.X.; Wang, Y.Q.; Wang, A.Q.; Yu, H.Y.; Pu, Z.E. Chitin synthase gene FgCHS8 affects virulence and fungal cell wall sensitivity to environmental stress in Fusarium graminearum. Fungal Biol. 2016, 120, 764–774. [Google Scholar] [CrossRef]
- Bansal, N.; Kanwar, S.S. Peroxidase(s) in environment protection. Sci. World J. 2013, 2013, 714639. [Google Scholar] [CrossRef]
- Asgher, M.; Bhatti, H.N.; Ashraf, M.; Legge, R.L. Recent developments in biodegradation of industrial pollutants by white rot fungi and their enzyme system. Biodegradation 2008, 19, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Cambaza, E.; Koseki, S.; Kawamura, S. Fusarium graminearum colors and deoxynivalenol synthesis at different water activity. Foods 2018, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Brana, M.T.; Sergio, L.; Haidukowski, M.; Logrieco, A.F.; Altomare, C. Degradation of aflatoxin B1 by a sustainable enzymatic extract from spent mushroom substrate of Pleurotus eryngii. Toxins (Basel) 2020, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Yehia, R.S. Aflatoxin detoxification by manganese peroxidase purified from Pleurotus ostreatus. Braz. J. Microbiol. 2014, 45, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Couto, S.R.; Moldes, D.; Sanromán, M.A. Optimum stability conditions of pH and temperature for ligninase and manganese-dependent peroxidase from Phanerochaete chrysosporium. Application to in vitro decolorization of Poly R-478 by MnP. World J. Microb. Biot. 2006, 22, 607–612. [Google Scholar] [CrossRef]
- Xu, H.; Guo, M.Y.; Gao, Y.H.; Bai, X.H.; Zhou, X.W. Expression and characteristics of manganese peroxidase from Ganoderma lucidum in Pichia pastoris and its application in the degradation of four dyes and phenol. BMC Biotechnol. 2017, 17, 19. [Google Scholar] [CrossRef] [PubMed]
- Christian, V.; Shrivastava, R.; Shukla, D.; Modi, H.; Vyas, B.R.M. Mediator role of veratryl alcohol in the lignin peroxidase-catalyzed oxidative decolorization of Remazol Brilliant Blue R. Enzyme Microb. Technol. 2005, 36, 327–332. [Google Scholar] [CrossRef]
- Leontievsky, A.A.; Myasoedova, N.M.; Baskunov, B.P.; Golovleva, L.A.; Bucke, C.; Evans, C.S. Transformation of 2,4,6-trichlorophenol by free and immobilized fungal laccase. Appl. Microbiol. Biotechnol. 2001, 57, 85–91. [Google Scholar] [CrossRef]
- De Souza, D.F.; Tychanowicz, G.K.; de Souza, C.G.; Peralta, R.M. Co-production of ligninolytic enzymes by Pleurotus pulmonarius on wheat bran solid state cultures. J. Basic Microbiol. 2006, 46, 126–134. [Google Scholar] [CrossRef]
- Tychanowicz, G.K.; de Souza, D.F.; Souza, C.G.M.; Kadowaki, M.K.; Peralta, R.M. Copper improves the production of laccase by the white-rot fungus Pleurotus pulmonarius in solid state fermentation. Braz. Arch. Biol. Technol. 2006, 49, 699–704. [Google Scholar] [CrossRef]
- Cho, J.S.; Seo, Y.C.; Yim, T.B.; Lee, H.Y. Effect of Nanoencapsulated vitamin B1 derivative on inhibition of both mycelial growth and spore germination of Fusarium oxysporum f. sp. raphani. Int. J. Mol. Sci. 2013, 14, 4283–4297. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Shen, S.; Zhou, W.; Wang, F.; Fan, Y. Fusarium graminearum growth inhibition due to glucose starvation caused by osthol. Int. J. Mol. Sci. 2008, 9, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Tso, K.H.; Ju, J.C.; Fan, Y.K.; Chiang, H.I. Enzyme degradation reagents effectively remove mycotoxins deoxynivalenol and zearalenone from pig and poultry artificial digestive juices. Toxins (Basel) 2019, 11, 599. [Google Scholar] [CrossRef] [PubMed]
- Park, D.L.; Troxell, T.C. US perspective on mycotoxin regulatory issues. Mycotoxins Food Saf. 2002, 504, 277–285. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Incubation Time, d | ||||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| Diameter, cm | |||||||
| Control 1 | 2.86 ± 0.73 | 4.78 ± 1.04 | 5.28 ± 1.16 | 6.50 ± 1.91 | 6.85 ± 1.99 | 7.78 ± 2.43 | 9.50 ± 0.40 |
| MnP | 2.90 ± 0.38 | 4.70 ± 0.63 | 5.28 ± 0.73 | 6.12 ± 1.30 | 6.34 ± 1.12 | 6.62 ± 1.24 | 7.28 ± 1.25 |
| LiP | 1.54 ± 0.36 | 2.02 ± 0.53 | 2.30 ± 0.40 | 2.28 ± 0.12 | 2.26 ± 0.60 | 2.45 ± 0.54 | 2.41 ± 0.29 |
| Inhibition rate, % | |||||||
| MnP | −5.26 ± 18.7 b | −0.50 ± 12.3 b | −2.11 ± 12.8 b | 3.30 ± 11.3 b | 3.97 ± 16.4 b | 9.44 ± 23.6 b | 23.7 ± 10.2 b |
| LiP | 45.2 ± 8.67 a | 57.8 ± 4.88 a | 55.9 ± 3.54 a | 62.1 ± 11.4 a | 66.6 ± 4.77 a | 67.3 ± 5.10 a | 74.7 ± 2.23 a |
| Item | Incubation Time, h | |||
|---|---|---|---|---|
| 1 | 3 | 6 | 12 | |
| Chitinase activity, GlcNAc mg/g fresh weight/h | ||||
| Control 1 | 0.000 ± 0.000 b | 0.012 ± 0.001 c | 0.033 ± 0.001 c | 0.075 ± 0.003 b |
| MnP | 0.014 ± 0.002 a | 0.066 ± 0.011 b | 0.095 ± 0.006 b | 0.117 ± 0.006 b |
| LiP | 0.014 ± 0.003 a | 0.120 ± 0.015 a | 0.179 ± 0.028 a | 0.231 ± 0.043 a |
| GlcNAc content, mg/g fresh weight | ||||
| Control 1 | 0.000 ± 0.000 b | 0.011 ± 0.000 c | 0.040 ± 0.002 c | 0.092 ± 0.002 b |
| MnP | 0.020 ± 0.002 a | 0.072 ± 0.012 b | 0.084 ± 0.009 b | 0.093 ± 0.017 b |
| LiP | 0.019 ± 0.003 a | 0.114 ± 0.010 a | 0.174 ± 0.012 a | 0.286 ± 0.021 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tso, K.-H.; Lumsangkul, C.; Ju, J.-C.; Fan, Y.-K.; Chiang, H.-I. The Potential of Peroxidases Extracted from the Spent Mushroom (Flammulina velutipes) Substrate Significantly Degrade Mycotoxin Deoxynivalenol. Toxins 2021, 13, 72. https://doi.org/10.3390/toxins13010072
Tso K-H, Lumsangkul C, Ju J-C, Fan Y-K, Chiang H-I. The Potential of Peroxidases Extracted from the Spent Mushroom (Flammulina velutipes) Substrate Significantly Degrade Mycotoxin Deoxynivalenol. Toxins. 2021; 13(1):72. https://doi.org/10.3390/toxins13010072
Chicago/Turabian StyleTso, Ko-Hua, Chompunut Lumsangkul, Jyh-Cherng Ju, Yang-Kwang Fan, and Hsin-I Chiang. 2021. "The Potential of Peroxidases Extracted from the Spent Mushroom (Flammulina velutipes) Substrate Significantly Degrade Mycotoxin Deoxynivalenol" Toxins 13, no. 1: 72. https://doi.org/10.3390/toxins13010072
APA StyleTso, K.-H., Lumsangkul, C., Ju, J.-C., Fan, Y.-K., & Chiang, H.-I. (2021). The Potential of Peroxidases Extracted from the Spent Mushroom (Flammulina velutipes) Substrate Significantly Degrade Mycotoxin Deoxynivalenol. Toxins, 13(1), 72. https://doi.org/10.3390/toxins13010072