Proteomic Insights into Starvation of Nitrogen-Replete Cells of Nostoc sp. PCC 7120 under β-N-Methylamino-L-Alanine (BMAA) Treatment

 ,
,  ,
,
Abstract
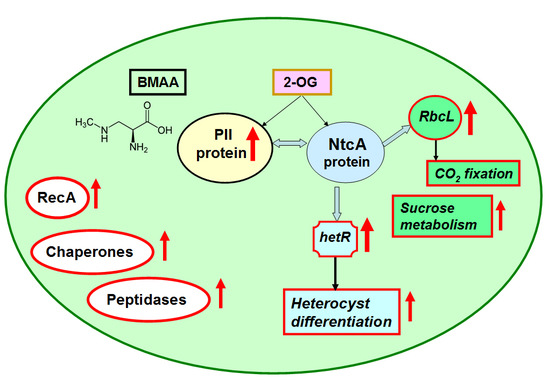
1. Introduction
2. Results and Discussion
2.1. Proteins Affected by BMAA Under Nitrogen-Replete Conditions
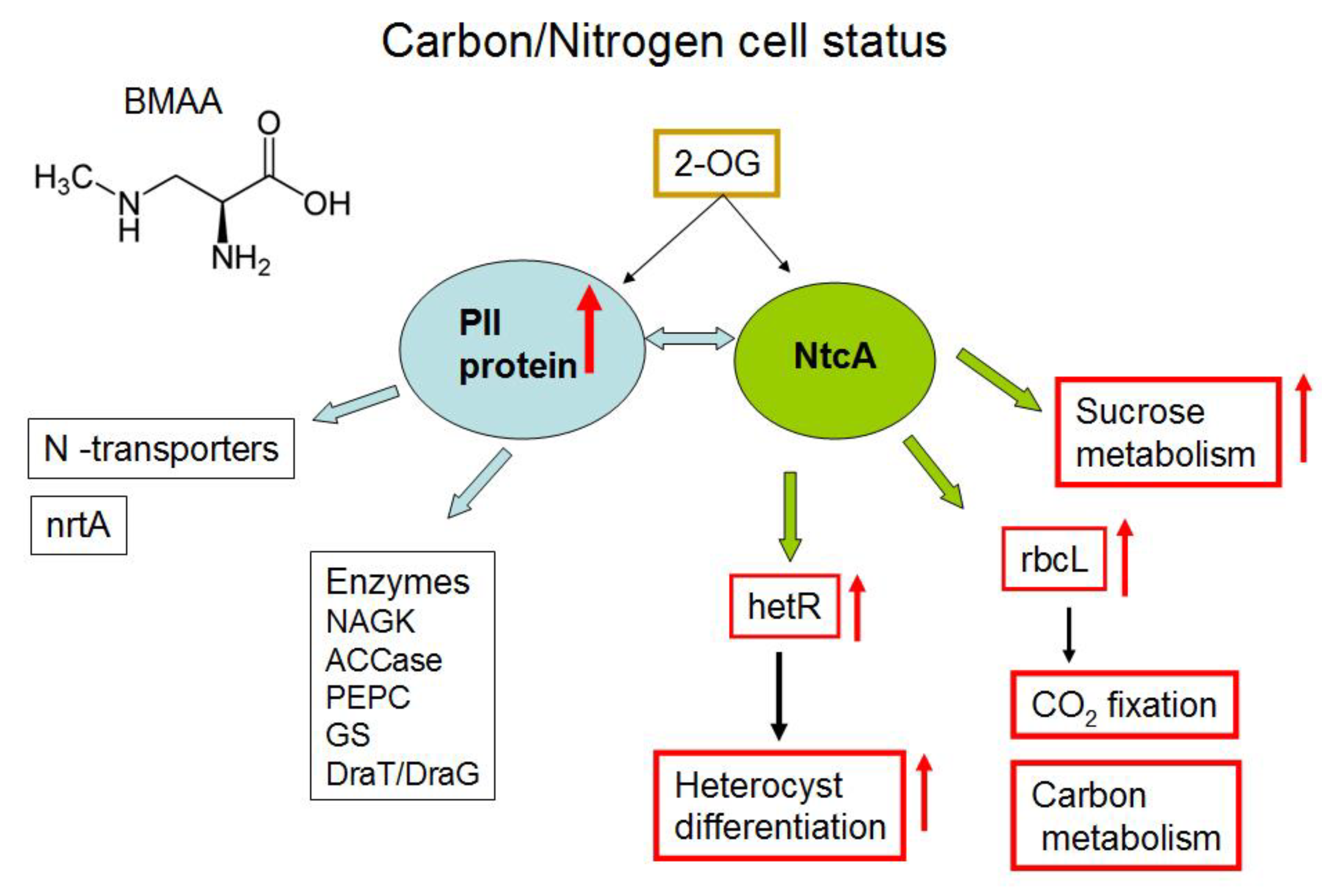
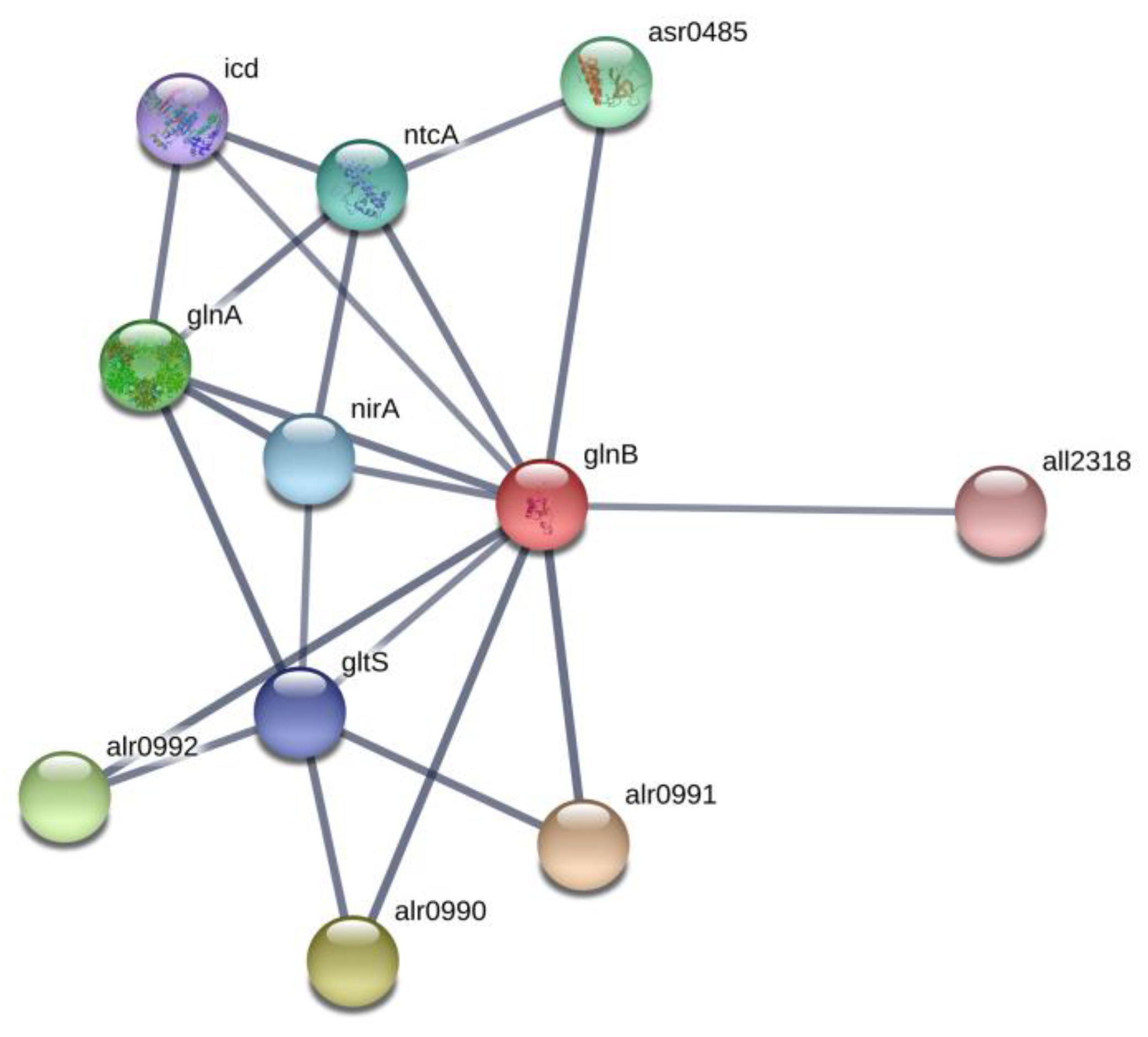
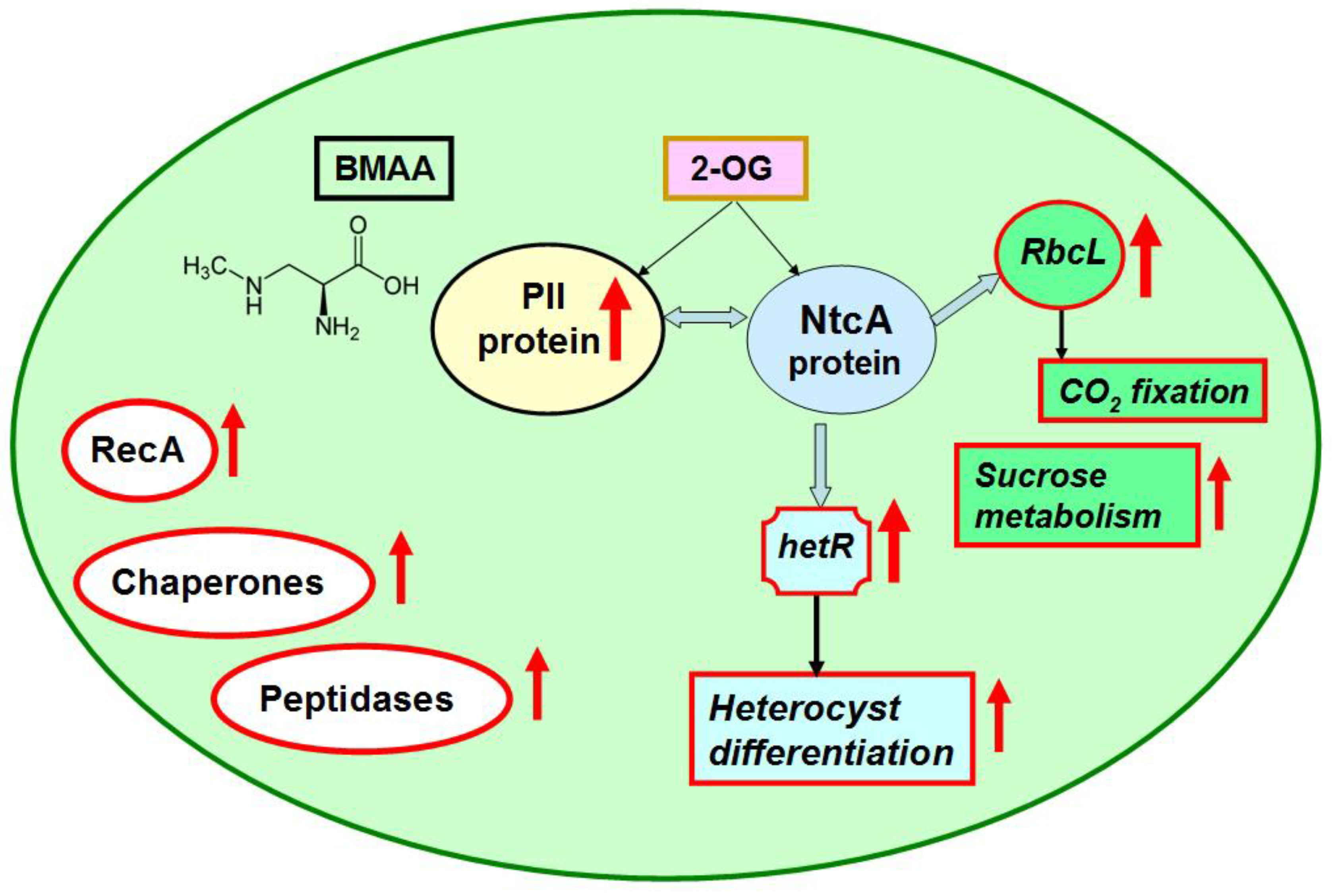
2.2. Nitrogen Status Sensing and Nitrogen Assimilation
2.3. CO2 Fixation and Carbon Dioxide-Concentrating Mechanism
2.4. Starch and Sucrose Metabolism
2.5. Photosynthesis
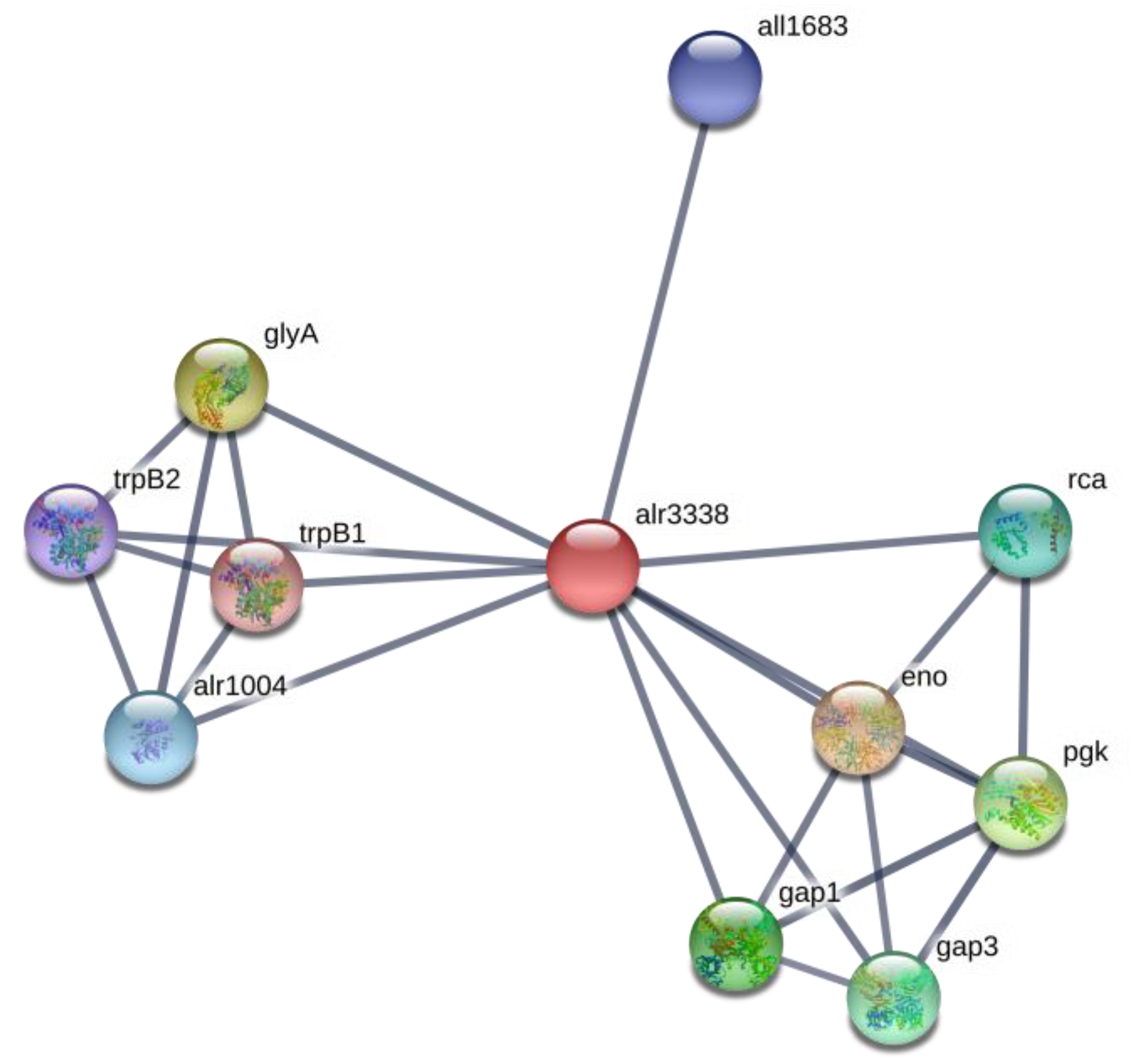
2.6. Amino Acid Metabolism
2.7. Signaling and Stress Response Proteins, Proteases and Chaperones
2.8. Hypothetical Proteins
3. Conclusions
4. Materials and Methods
4.1. Cyanobacterial Strain and Cultivation Conditions
4.2. Trypsin Digestion in Solution
4.3. Liquid Chromatography with Tadem Mass Spetrometry (LC-MS/MS) Analysis
4.4. Protein Identification by LC-MS/MS Data Analysis
4.5. Pathway Analysis Based on LC-MS/MS Data
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mishra, A.K.; Tewari, D.N.; Rai, A.N. Cyanobacteria 1st Edition from Basic Science to Applications; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.A.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef]
- Omidi, A.; Esterhuizen, M.; Pflugmacher, S. Still challenging: The ecological function of the cyanobacterial toxin microcystin—What we know so far. Toxin Rev. 2017, 37, 87–105. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B. In search of environmental role of cylindrospermopsin: A review on global distribution and ecology of its producers. Water Res. 2014, 66, 320–337. [Google Scholar] [CrossRef]
- Cusick, K.D.; Sayler, G.S. An Overview on the Marine Neurotoxin, Saxitoxin: Genetics, Molecular Targets, Methods of Detection and Ecological Functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce -N-methylamino-L-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A Novel Source for the Neurotoxin BMAA in Aquatic Environments. PLoS ONE 2014, 9, e84578. [Google Scholar] [CrossRef] [PubMed]
- Besemer, K. Biodiversity, community structure and function of biofilms in stream ecosystems. Res. Microbiol. 2015, 166, 774–781. [Google Scholar] [CrossRef]
- Murch, S.J.; Cox, P.A.; Banack, S.A. A mechanism for slow release of biomagnified cyanobacterial neurotoxins and neurodegenerative disease in Guam. Proc. Nat. Acad. Sci. USA 2004, 101, 12228–12231. [Google Scholar] [CrossRef]
- Jonasson, S.; Eriksson, J.; Berntzon, L.; Spacil, Z.; Ilag, L.L.; Ronnevi, L.-O.; Rasmussen, U.; Bergman, B. Transfer of a cyanobacterial neurotoxin within a temperate aquatic ecosystem suggests pathways for human exposure. Proc. Natl. Acad. Sci. USA 2010, 107, 9252–9257. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Kiselova, N.; Rosén, J.; Ilag, L.L. Quantification of neurotoxin BMAA (β-N-methylamino-L-alanine) in seafood from Swedish markets. Sci. Rep. 2014, 4, 6931. [Google Scholar] [CrossRef]
- Salomonsson, M.L.; Fredriksson, E.; Alfjorden, A.; Hedeland, M.; Bondesson, U. Seafood sold in Sweden contains BMAA: A study of free and total concentrations with UHPLC-MS/MS and dansyl chloride derivatization. Toxicol. Rep. 2015, 2, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Hirano, A.; Malamud, N.; Elizan, T.S.; Kurland, L.T. Amyotrophic Lateral Sclerosis and Parkinsonism-Dementia Complex on Guam. Arch. Neurol. 1966, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Cox, P.A.; Banack, S.A.; Steele, J.C.; Sacks, O.W. Occurrence of beta-methylamino-l-alanine (BMAA) in ALS/PDC patients from Guam. Acta Neurol. Scand. 2004, 110, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.A.; Koksharova, O.A. Neurotoxic non-proteinogenic amino acid β-N-methylamino-L-alanine and its role in biological systems. Biochemistry 2016, 81, 794–805. [Google Scholar] [CrossRef]
- Downing, S.; Banack, S.; Metcalf, J.S.; Cox, P.A.; Downing, T. Nitrogen starvation of cyanobacteria results in the production of β-N-methylamino-L-alanine. Toxicon 2011, 58, 187–194. [Google Scholar] [CrossRef]
- Downing, T.; Phelan, R.; Downing, S. A potential physiological role for cyanotoxins in cyanobacteria of arid environments. J. Arid. Environ. 2015, 112, 147–151. [Google Scholar] [CrossRef]
- Downing, S.; Downing, T.G. The metabolism of the non-proteinogenic amino acid β-N-methylamino-L-alanine (BMAA) in the cyanobacterium Synechocystis PCC6803. Toxicon 2016, 115, 41–48. [Google Scholar] [CrossRef]
- Nunn, P.B.; Codd, G.A. Metabolic solutions to the biosynthesis of some diaminomonocarboxylic acids in nature: Formation in cyanobacteria of the neurotoxins 3-N-methyl-2,3-diaminopropanoic acid (BMAA) and 2,4-diaminobutanoic acid (2,4-DAB). Phytochemistry 2017, 144, 253–270. [Google Scholar] [CrossRef]
- Berntzon, L.; Erasmie, S.; Celepli, N.; Eriksson, J.; Rasmussen, U.; Bergman, B. BMAA Inhibits Nitrogen Fixation in the Cyanobacterium Nostoc sp. PCC 7120. Mar. Drugs 2013, 11, 3091–3108. [Google Scholar] [CrossRef]
- Popova, A.A.; Rasmussen, U.; Semashko, T.A.; Govorun, V.; Koksharova, O.A. Stress effects of cyanotoxin β-methylamino-L-alanine (BMAA) on cyanobacterial heterocyst formation and functionality. Environ. Microbiol. Rep. 2018, 10, 369–377. [Google Scholar] [CrossRef]
- Popova, A.A.; Semashko, T.A.; Kostina, N.V.; Rasmussen, U.; Govorun, V.; Koksharova, O.A. The Cyanotoxin BMAA Induces Heterocyst Specific Gene Expression in Anabaena sp. PCC 7120 under Repressive Conditions. Toxins 2018, 10, 478. [Google Scholar] [CrossRef] [PubMed]
- Koksharova, O.A.; Butenko, I.; Pobeguts, O.; Safronova, N.; Govorun, V. The First Proteomic Study of Nostoc sp. PCC 7120 Exposed to Cyanotoxin BMAA under Nitrogen Starvation. Toxins 2020, 12, 310. [Google Scholar] [CrossRef] [PubMed]
- Herrero, A.; Stavans, J.; Flores, E. The multicellular nature of filamentous heterocyst-forming cyanobacteria. FEMS Microbiol. Rev. 2016, 40, 831–854. [Google Scholar] [CrossRef] [PubMed]
- Liotenberg, S.; Campbell, D.; Castets, A.-M.; Houmard, J.; De Marsac, N.T. Modification of the pIIprotein in response to carbon and nitrogen availability in filamentous heterocystous cyanobacteria. FEMS Microbiol. Lett. 1996, 144, 185–190. [Google Scholar] [CrossRef][Green Version]
- Forchhammer, K. PII signal transducers: Novel functional and structural insights. Trends Microbiol. 2008, 16, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Forchhammer, K.; Lüddecke, J. Sensory properties of the P II signalling protein family. FEBS J. 2015, 283, 425–437. [Google Scholar] [CrossRef]
- Forchhammer, K.; A Selim, K. Carbon/nitrogen homeostasis control in cyanobacteria. FEMS Microbiol. Rev. 2019, 025. [Google Scholar] [CrossRef]
- Watzer, B.; Spät, P.; Neumann, N.; Koch, M.; Sobotka, R.; Macek, B.; Hennrich, O.; Forchhammer, K. The Signal Transduction Protein PII Controls Ammonium, Nitrate and Urea Uptake in Cyanobacteria. Front. Microbiol. 2019, 10, 1428. [Google Scholar] [CrossRef]
- Wei, T.F.; Ramasubramanian, T.S.; Golden, J.W. Anabaena sp. strain PCC 7120 ntcA gene required for growth on nitrate and heterocyst development. J. Bacteriol. 1994, 176, 4473–4482. [Google Scholar] [CrossRef]
- Flores, E.; Picossi, S.; Valladares, A.; Herrero, A. Transcriptional regulation of development in heterocyst-forming cyanobacteria. Biochim. Biophys. Acta Bioenerg. 2019, 1862, 673–684. [Google Scholar] [CrossRef]
- Herrero, A.; Flores, E. Genetic responses to carbon and nitrogen availability in Anabaena. Environ. Microbiol. 2019, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Knaff, D.B. Glutamate synthase: Structural, mechanistic, and regulatory properties, and role in the amino acid metabolism. Photosynth. Res. 2005, 83, 191–217. [Google Scholar] [CrossRef] [PubMed]
- Huergo, L.F.; Dixon, R. The Emergence of 2-Oxoglutarate as a Master Regulator Metabolite. Microbiol. Mol. Boil. Rev. 2015, 79, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Muro-Pastor, M.I.; Reyes, J.; Florencio, F.J. Cyanobacteria perceive nitrogen status by sensing intracellular 2-oxoglutarate levels. J. Boil. Chem. 2001, 276, 38328. [Google Scholar]
- Chen, H.-L.; Latifi, A.; Zhang, C.-C.; Bernard, C. Biosensors-Based In Vivo Quantification of 2-Oxoglutarate in Cyanobacteria and Proteobacteria. Life 2018, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Alles, L.; Lesniewska, É.; Root, K.; Aubry, N.; Pocholle, N.; Mendoza, C.I.; Bourillot, E.; Barylyuk, K.; Pompon, D.; Zenobi, R.; et al. Spontaneous non-canonical assembly of CcmK hexameric components from β-carboxysome shells of cyanobacteria. PLoS ONE 2017, 12, e0185109. [Google Scholar] [CrossRef]
- Koropatkin, N.M.; Koppenaal, D.W.; Pakrasi, H.B.; Smith, T.J. The Structure of a Cyanobacterial Bicarbonate Transport Protein, CmpA. J. Boil. Chem. 2006, 282, 2606–2614. [Google Scholar] [CrossRef]
- Ball, S.; Colleoni, C.; Arias, M.C. The Transition from Glycogen to Starch Metabolism in Cyanobacteria and Eukaryotes. Starch 2015, 93–158. [Google Scholar] [CrossRef]
- Kadouche, D.; Ducatez, M.; Cenci, U.; Tirtiaux, C.; Suzuki, E.; Nakamura, Y.; Putaux, J.-L.; Durand-Terrasson, A.; Díaz-Troya, S.; Florencio, F.J.; et al. Characterization of Function of the GlgA2 Glycogen/Starch Synthase in Cyanobacterium sp. Clg1 Highlights Convergent Evolution of Glycogen Metabolism into Starch Granule Aggregation1. Plant Physiol. 2016, 171, 1879–1892. [Google Scholar] [CrossRef]
- Chia, T.; Thorneycroft, D.; Chapple, A.; Messerli, G.; Chen, J.; Zeeman, S.; Smith, S.M.; Smith, A.M. A cytosolic glucosyltransferase is required for conversion of starch to sucrose in Arabidopsis leaves at night. Plant J. 2004, 37, 853–863. [Google Scholar] [CrossRef]
- Kolman, M.A.; Nishi, C.N.; Perez-Cenci, M.; Salerno, G. Sucrose in Cyanobacteria: From a Salt-Response Molecule to Play a Key Role in Nitrogen Fixation. Life 2015, 5, 102–126. [Google Scholar] [CrossRef] [PubMed]
- Marcozzi, C.; Cumino, A.C.; Salerno, G. Role of NtcA, a cyanobacterial global nitrogen regulator, in the regulation of sucrose metabolism gene expression in Anabaena sp. PCC 7120. Arch. Microbiol. 2008, 191, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Cumino, A.C.; Marcozzi, C.; Barreiro, R.; Salerno, G. Carbon Cycling in Anabaena sp. PCC 7120. Sucrose Synthesis in the Heterocysts and Possible Role in Nitrogen Fixation1[OA]. Plant Physiol. 2007, 143, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Herrero, A.; Flores, E. The Cyanobacteria: Molecular Biology, Genomics and Evolution; Caister Academic Press: Norfolk, UK, 2008. [Google Scholar]
- Spät, P.; Klotz, A.; Rexroth, S.; Macek, B.; Forchhammer, K. Chlorosis as a Developmental Program in Cyanobacteria: The Proteomic Fundament for Survival and Awakening. Mol. Cell. Proteom. 2018, 17, 1650–1669. [Google Scholar] [CrossRef] [PubMed]
- Quintero, M.J.; Muro-Pastor, A.M.; Herrero, A.; Flores, E. Arginine Catabolism in the Cyanobacterium Synechocystis sp. Strain PCC 6803 Involves the Urea Cycle and Arginase Pathway. J. Bacteriol. 2000, 182, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Colman, B.; Norman, E.G. Serine synthesis in cyanobacteria by a nonphotorespiratory pathway. Physiol. Plant. 1997, 100, 133–136. [Google Scholar] [CrossRef]
- Walton, N.J.; Woolhouse, H.W. Enzymes of serine and glycine metabolism in leaves and non-photosynthetic tissues of Pisum sativum L. Planta 1986, 167, 119–128. [Google Scholar] [CrossRef]
- Timm, S.; Florian, A.; Wittmiß, M.; Jahnke, K.; Hagemann, M.; Fernie, A.R.; Bauwe, H. Serine Acts as a Metabolic Signal for the Transcriptional Control of Photorespiration-Related Genes in Arabidopsis. Plant Physiol. 2013, 162, 379–389. [Google Scholar] [CrossRef]
- Engskog, M.K.R.; Ersson, L.; Haglöf, J.; Arvidsson, T.; Pettersson, C.; Brittebo, E. β-N-Methylamino-L-alanine (BMAA) perturbs alanine, aspartate and glutamate metabolism pathways in human neuroblastoma cells as determined by metabolic profiling. Amino Acids 2017, 49, 905–919. [Google Scholar] [CrossRef]
- Koksharova, O.A.; Butenko, I.O.; Pobeguts, O.V.; Safronova, N.A.; Govorun, V.M. Severe Stress Effect of Cyanotoxin β-N-methylamino-L-alanine (BMAA) on Nostoc sp. PCC 7120 Proteome under Diazotrofic Conditions. Toxins 2020, 12, 310. [Google Scholar] [CrossRef]
- Adams, E.; Goldstone, A. Hydroxyproline metabolism. IV. Enzymatic synthesis of gamma-hydroxyglutamate from Delta 1-pyrroline-3-hydroxy-5-carboxylate. J. Boil. Chem. 1960, 235, 3504–3512. [Google Scholar]
- Weiss, J.H.; Choi, D.W. β-N-methylamino-L-alanine neurotoxicity: Requirement for bicarbonate as a cofactor. Science 1988, 241, 973–975. [Google Scholar] [CrossRef] [PubMed]
- Nunn, P.B.; O’Brien, P. The interaction of beta-N-methylamino-L-alanine with bicarbonate: An 1H-NMR study. FEBS Lett. 1989, 251, 31–35. [Google Scholar] [CrossRef]
- Myers, T.G.; Nelson, S.D. Neuroactive carbamate adducts of beta-N-methylamino-L-alanine and ethyl-enediamine. Detection and quantitation under physiological conditions by 13C NMR, J. Biol. Chem. 1990, 265, 10193–10195. [Google Scholar]
- Moreno-Cinos, C.; Goossens, K.; Salado, I.G.; van der Veken, P.; de Winter, H.; Augustyns, K. ClpP Protease, a Promising Antimicrobial Target. Int. J. Mol. Sci. 2019, 20, 2232. [Google Scholar] [CrossRef]
- Hedstrom, L. Serine Protease Mechanism and Specificity. Chem. Rev. 2002, 102, 4501–4524. [Google Scholar] [CrossRef]
- Dunlop, R.A.; Cox, P.A.; Banack, S.A.; Rodgers, K.J. The Non-Protein Amino Acid BMAA Is Misincorporated into Human Proteins in Place of l-Serine Causing Protein Misfolding and Aggregation. PLoS ONE 2013, 8, e75376. [Google Scholar] [CrossRef]
- Han, N.-C.; Bullwinkle, T.J.; Loeb, K.F.; Faull, K.F.; Mohler, K.; Rinehart, J.; Ibba, M. The mechanism of β-N-methylamino-l-alanine inhibition of tRNA aminoacylation and its impact on misincorporation. J. Boil. Chem. 2019, 295, 1402–1410. [Google Scholar] [CrossRef]
- Khumalo, M.J.; Nzuza, N.; Padayachee, T.; Chen, W.; Yu, J.-H.; Nelson, D.R.; Syed, K. Comprehensive Analyses of Cytochrome P450 Monooxygenases and Secondary Metabolite Biosynthetic Gene Clusters in Cyanobacteria. Int. J. Mol. Sci. 2020, 21, 656. [Google Scholar] [CrossRef]
- Cryle, M.J.; Stok, J.E.; de Voss, J.J. Reactions Catalyzed by Bacterial Cytochromes P450. Aust. J. Chem. 2003, 56, 749–762. [Google Scholar] [CrossRef]
- Penning, T.M. The aldo-keto reductases (AKRs): Overview. Chem. Interact. 2014, 234, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Dalle-Donne, I.; Rossi, R.; Colombo, G.; Giustarini, D.; Milzani, A.D.G. Protein S-glutathionylation: A regulatory device from bacteria to humans. Trends Biochem. Sci. 2009, 34, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Nunn, P.B.; Ponnusamy, M. β-N-Methylaminoalanine (BMAA): Metabolism and metabolic effects in model systems and in neural and other tissues of the rat in vitro. Toxicon 2009, 54, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Nunn, P.B. Three phases of research on?-N-methylamino-L-alanine (BMAA) a neurotoxic amino acid. Amyotroph. Lateral Scler. 2009, 10, 26–33. [Google Scholar] [CrossRef]
- Cassier-Chauvat, C.; Veaudor, T.; Chauvat, F. Comparative Genomics of DNA Recombination and Repair in Cyanobacteria: Biotechnological Implications. Front. Microbiol. 2016, 7, 1809. [Google Scholar] [CrossRef]
- Frías, J.E.; Flores, E.; Herrero, A. Activation of the Anabaena nir operon promoter requires both NtcA (CAP family) and NtcB (LysR family) transcription factors. Mol. Microbiol. 2000, 38, 613–625. [Google Scholar] [CrossRef]
- Luque, I.; Contreras, A.; Zabulon, G.; Herrero, A.; Houmard, J. Expression of the glutamyl-tRNA synthetase gene from the cyanobacterium Synechococcus sp. PCC 7942 depends on nitrogen availability and the global regulator NtcA. Mol. Microbiol. 2002, 46, 1157–1167. [Google Scholar] [CrossRef]
- Herrero, A.; Muro-Pastor, A.M.; Flores, E. Nitrogen Control in Cyanobacteria. J. Bacteriol. 2001, 183, 411–425. [Google Scholar] [CrossRef]
- Buchanan, J.M. The Amidotransferases. Adv. Enzymol. Relat. Areas Mol. Biol. 2006, 39, 91–183. [Google Scholar] [CrossRef]
- Gleason, F.K. Thioredoxins in Cyanobacteria: Structure and Redox Regulation of Enzyme Activity. In Functional Organization of the Plant Nucleus; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1994; pp. 715–729. [Google Scholar]
- Lindahl, M.; Florencio, F.J. Thioredoxin-Linked Processes in Cyanobacteria are as Numerous as in Chloroplasts, But Targets are Different. Proc. Nat. Acad. Sci. USA 2003, 100, 16107–16112. [Google Scholar] [CrossRef]
- Qiu, X.-B.; Shao, Y.-M.; Miao, S.; Wang, L. The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. Cell. Mol. Life Sci. 2006, 63, 2560–2570. [Google Scholar] [CrossRef] [PubMed]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Arapidi, G.; Osetrova, M.; Ivanova, O.; Butenko, I.; Saveleva, T.; Pavlovich, P.; Anikanov, N.; Ivanov, V.; Govorun, V. Peptidomics dataset: Blood plasma and serum samples of healthy donors fractionated on a set of chromatography sorbents. Data Brief 2018, 18, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially Modified Protein Abundance Index (emPAI) for Estimation of Absolute Protein Amount in Proteomics by the Number of Sequenced Peptides per Protein. Mol. Cell. Proteom. 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Picossi, S.; Flores, E.; Herrero, A. ChIP analysis unravels an exceptionally wide distribution of DNA binding sites for the NtcA transcription factor in a heterocyst-forming cyanobacterium. BMC Genom. 2014, 15, 22. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Pathway | Number of Proteins Affected by BMAA | Total Amount | |
|---|---|---|---|---|
| Upregulated | Downregulated | |||
| 1 | Nitrogen metabolism | 2 | 1 | 1 |
| 2 | Photosynthesis | 7 | 2 | 5 |
| 3 | CO2 fixation and CO2-concentrating mechanism | 7 | 3 | 4 |
| 4 | Cell division | 2 | 0 | 2 |
| 5 | Regulatory proteins, proteases | 10 | 5 | 4 |
| 6 | Transporters | 2 | 2 | 0 |
| 7 | Stress response | 6 | 5 | 1 |
| 8 | Translation | 7 | 5 | 2 |
| 9 | Amino acid synthesis and metabolism | 10 | 5 | 5 |
| 10 | Purine and Pyrimidine metabolism | 4 | 2 | 2 |
| 11 | Starch and sucrose metabolism | 2 | 2 | 0 |
| 12 | Sulfur metabolism | 1 | 0 | 1 |
| 13 | Fatty acid biosynthesis | 1 | 1 | 0 |
| 14 | Energy metabolism | 2 | 1 | 1 |
| 15 | Hypothetical proteins | 18 | 10 | 8 |
| Total | 80 | 44 | 36 | |
| No. | Protein | Gene | Function | Fold Change LFQ Ratio BMAA-Treated/Control | p-Value | |
|---|---|---|---|---|---|---|
| Nitrogen Metabolism (Two Proteins) | ||||||
| 1 | GlnB|P-II | all2319* | Nitrogen regulatory protein | 1.75 | 0.0999 | |
| 2 | glutamate synthase (ferredoxin) [EC:1.4.7.1]|(GenBank) gltS; ferredoxin-glutamate synthase | alr4344 | Nitrogen metabolism and Glyoxylate and dicarboxylate metabolism | 0.71 | 0.0226 | |
| Photosynthesis (7 Proteins) | ||||||
| 3 | cpcB | alr0528 | phycocyanin beta chain | 1.14 | 0.0909 | |
| 4 | cpcG4 | alr0537 | phycobilisome rod-core linker protein | 1.18 | 0.0625 | |
| 5 | EC:7.1.2.2,7.2.2.1 atpB ATP synthase F0F1 subunit beta F-type H+/Na+-transporting ATPase subunit beta | all5039 | Oxidative phosphorylation, Photosynthesis | 0.83 | 0.0858 | |
| 6 | psbB | all0138 | photosystem II CP47 protein | 0.82 | 0.0717 | |
| 7 | psbV | all0259 | cytochrome c-550 | 0.71 | 0.0533 | |
| 8 | apcF | all2327 | phycobilisome core component | 0.82 | 0.0533 | |
| 9 | psaE; photosystem I protein E | asr4319 | photosystem I subunit IV | 0.69 | 0.0040 | |
| CO2 Fixation and Carbon Dioxide Concentration Mechanism (7 Proteins) | ||||||
| 10 | RbcL [EC:4.1.1.39] | alr1524* | CO2 fixation ribulosebisophosphate carboxylase large chain | 1.67 | 0.0535 | |
| 11 | Rca | alr1533* | CO2 fixation RuBisCO activase | 1.79 | 0.0112 | |
| 12 | CcmK | all0868 | CO2 fixation Carbon dioxide-concentrating mechanism protein | 1.82 | 0.0103 | |
| 13 | RbcS [EC:4.1.1.39] | alr1526* | CO2 fixation Ribulose bisphosphate carboxylase small chain | 0.68 | 0.0148 | |
| 14 | CcmK | alr0318 | CO2 fixation Carbon dioxide-concentrating mechanism protein | 0.76 | 0.0533 | |
| 15 | CmpA bicarbonate transport bicarbonate-binding protein | alr2877 | Part of the ABC transporter complex Cmp ABCD involved in bicarbonate transport, binds bicarbonate with high affinity | found only in control sample | 0.0228 | |
| 16 | transketolase [EC:2.2.1.1] | alr3344 | Pentose phosphate pathway Carbon fixation in photosynthetic organisms | 0.79 | 0.0669 | |
| Cell Division (Two Proteins) | ||||||
| 17 | FtsH | all3642 | cell division protein | 0.76 | 0.0588 | |
| 18 | MinD | alr3456 | septum site-determining protein | 0.67 | 0.0008 | |
| Regulatory Proteins and Proteases (9 Proteins) | ||||||
| 19 | ClpP EC:3.4.21.92 | alr3683 | ATP-dependent protease ClpP proteolytic subunit | 1.85 | 0.0378 | |
| 20 | Serine proteinase | alr2758 | modification, protein turnover, chaperones | 1.82 | 0.0062 | |
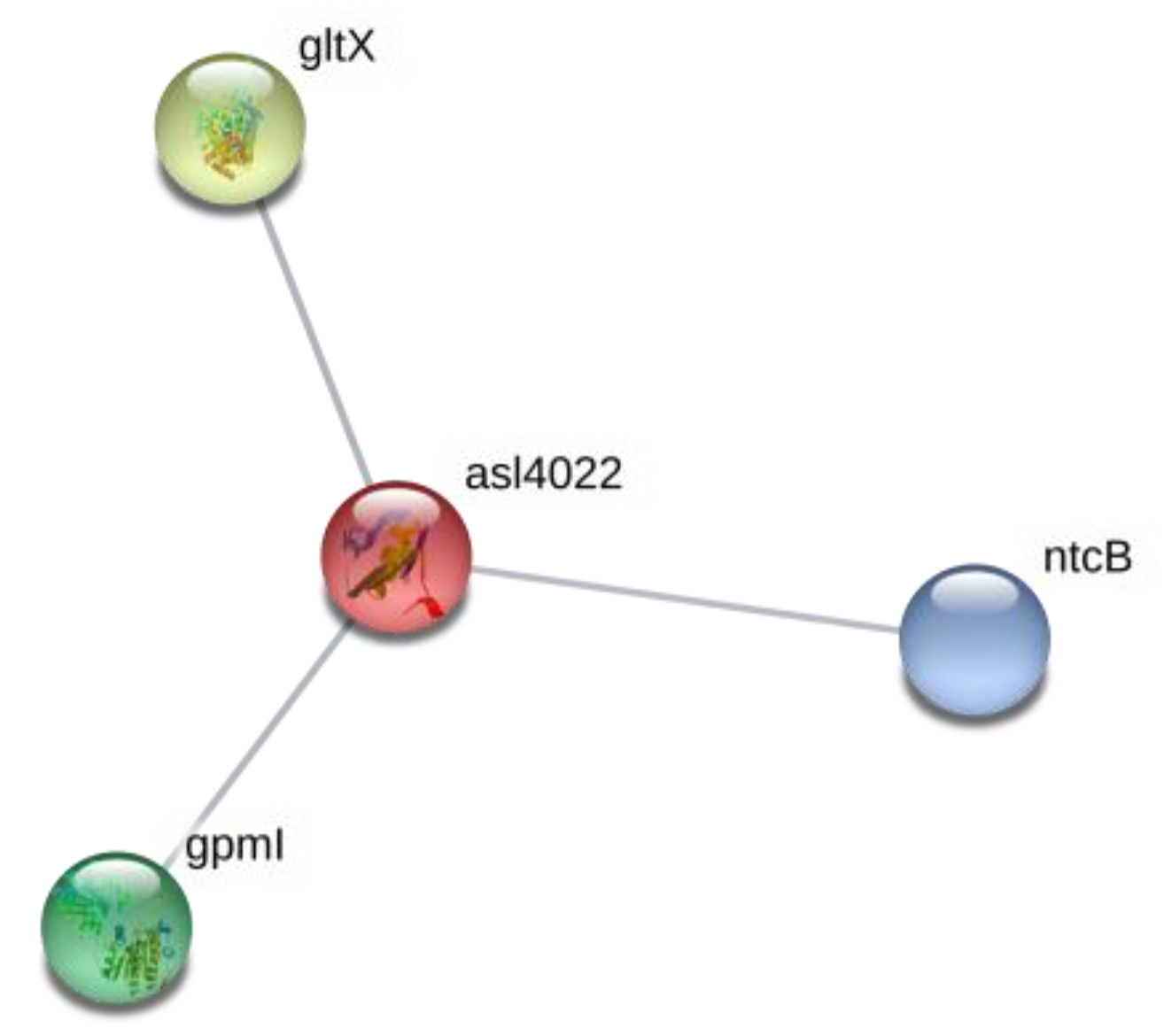
| 21 | RbpD|RNA-binding protein | asl4022 | 2.7 | 0.0339 | ||
| 22 | mRNA-binding protein | alr4831 | 1.72 | 0.0630 | ||
| 23 | Cytochrome P450 | all1361 | Cytochrome P450 monooxygenases is a heme-dependent enzyme that introduces oxygen into a wide range of molecules | 1.49 | 0.0177 | |
| 24 | YggE, uncharacterized conserved protein | all0089* | It contains kinase-interacting SIMPL domain | found only in control sample | 0.0162 | |
| 25 | MBL fold metallo-hydrolase | all0268 | https://www.ncbi.nlm.nih.gov/Structure/cdd/cl23716 | found only in control sample | 0.0037 | |
| 26 | RpaA DNA-binding response regulator, OmpR family | all0129* | two-component system, OmpR family, response regulator | 0.83 | 0.0089 | |
| 27 | DNA-binding protein HU, hanA | asr3935 | histone-like DNA-binding protein HU | 0.70 | 0.0007 | |
| Transporters (Two Proteins) | ||||||
| 28 | ABC transporter ATP-binding protein | alr2372 | ABC-2-type transport system ATP-binding protein | 1.49 | 0.0408 | |
| 29 | ABC transporter ATP-binding protein | alr1554* | ATP-binding cassette, subfamily B | 3.23 | 0.0375 | |
| Stress Response (6 Proteins) | ||||||
| 31 | thioredoxin 1 | all1866 | Chaperones and folding catalysts, reduces disulfide bonds in other proteins | found only in control sample | 0.0489 | |
| 32 | RecA | all3272 | recombinase A | 3.7 | 0.0417 | |
| 33 | Glutathione reductase (NADPH) [EC:1.8.1.7] | all4968 | Glutathione metabolism | 1.49 | 0.0202 | |
| 34 | leucylaminopeptidase [EC:3.4.11.1] | alr0237 | Glutathione metabolism | 1.49 | 0.0469 | |
| 35 | aldo/ketoreductase | all2316 | Positive role of AKR in detoxification of reactive carbonyl species (RCS) produced under oxidative stress | 2.04 | 0.0143 | |
| 36 | polyribonucleotidenucleotidyltransferase [EC:2.7.7.8] | all4396 | RNA degradation | 1.28 | 0.0632 | |
| Translation (7 Proteins) | ||||||
| 37 | IF-2 | alr3832 | translation initiation factor | 1.23 | 0.0395 | |
| 38 | 50S ribosomal protein L16 | all4208 | large subunit ribosomal protein L16 | 1.54 | 0.0142 | |
| 39 | RpsP protein | asr1953 | small subunit ribosomal protein S16 | 3.33 | 0.0007 | |
| 40 | DNA-directed RNA polymerase subunit omega [EC:2.7.7.6] | asr4648 | Promotes RNA polymerase assembly | 1.61 | 0.0073 | |
| 41 | isoleucyl-tRNAsynthetase [EC:6.1.1.5] | alr1073 | Aminoacyl-tRNA biosynthesis | 1.47 | 0.0313 | |
| 42 | protein S13 | all4193* | small subunit ribosomal | 0.46 | 0.0255 | |
| 43 | protein S10 | all4336 | small subunit ribosomal | 0.68 | 0.0201 | |
| Amino Acid Synthesis and Metabolism (10 Proteins) | ||||||
| 44 | valine-pyruvate aminotransferase [EC:2.6.1.66] | alr2811* | Valine, leucine and isoleucine biosynthesis | 0.625 | 0.0887 | |
| 45 | ilvG, acetolactate synthase I/II/III large subunit [EC:2.2.1.6] | all4613* | Valine, leucine and isoleucine biosynthesis | 0.51 | 0.0315 | |
| 46 | argininosuccinate synthase [EC:6.3.4.5] | alr4798 | Arginine biosynthesis Alanine, aspartate and glutamate metabolism | 0.72 | 0.0704 | |
| 47 | phosphoserine phosphatase [EC:3.1.3.3] | alr3338 | Glycine, serine and threonine metabolism | found only in control sample | 0.0191 | |
| 48 | ornithine carbamoyltransferase [EC:2.1.3.3] | alr4907 | Arginine biosynthesis | found only in control sample | 0.0015 | |
| 49 | NodM, glutamine-fructose-6-phosphatetransaminase (isomerizing) [EC:2.6.1.16] | alr3464 | Alanine, aspartate and glutamate metabolism | 1.43 | 0.0113 | |
| 50 | Urea subunit alpha [EC:3.5.1.5] | alr3670 | Arginine biosynthesis Purine metabolism | 1.59 | 0.0648 | |
| 51 | murE UDP-N-acetylmuramoyl-L-alanyl-D-glutamate-2,6-diaminopimelateligase [EC:6.3.2.13] | | all1663 | Lysine biosynthesis Peptidoglycan biosynthesis | 1.27 | 0.0623 | |
| 52 | RHH-type transcriptional regulator, proline utilization regulon repressor/proline dehydrogenase/delta 1-pyrroline-5-carboxylate dehydrogenase [EC:1.5.5.21.2.1.88] | alr0540 | Alanine, aspartate and glutamate metabolism Arginine and proline metabolism | 1.79 | 0.0362 | |
| 53 | cysteine synthase [EC:2.5.1.47] | all2521* | Cysteine and methionine metabolism Sulfur metabolism | 1.72 | 0.0266 | |
| Nucleotide Synthesis (4 Proteins) | ||||||
| 54 | phosphoribosylformyl- glycinamidine cyclo-ligase [EC:6.3.3.1] | alr3525 | Purine metabolism | 0.67 | 0.0605 | |
| 55 | Phosphoribosylamine- glycine ligase [EC:6.3.4.13] | alr3510 | Purine metabolism | found only in control sample | 0.0005 | |
| 56 | nucleoside-diphosphate kinase [EC:2.7.4.6] | alr3402 | Purine metabolism Pyrimidine metabolism | 1.92 | 0.0212 | |
| 57 | uracil phosphoribosyltransferase [EC:2.4.2.9] | all2063 | Pyrimidine metabolism | 2.04 | 0.0528 | |
| Starch and Sucrose Metabolism (Two Proteins) | ||||||
| 58 | 4-alpha-glucanotransferase [EC:2.4.1.25] | alr3871 | Starch and sucrose metabolism | 1.69 | 0.0008 | |
| 59 | fructokinase [EC:2.7.1.4] | alr0517 | Fructose and mannose metabolism Starch and sucrose metabolism | 1.69 | 0.0837 | |
| Sulfur Metabolism (One Protein) | ||||||
| 60 | phosphoadenosinephosphosulfate reductase [EC:1.8.4.8 1.8.4.10] | all4464 | Sulfur metabolism | 0.65 | 0.0151 | |
| Fatty Acid Biosynthesis (One Protein) | ||||||
| 61 | 3-oxoacyl-[acyl-carrier protein] reductase [EC:1.1.1.100] | alr1894 | Fatty acid biosynthesis Biotin metabolism | 2.86 | 0.0316 | |
| Energy Metabolism (Two Proteins) | ||||||
| 62 | NAD(P)H-quinone oxidoreductase subunit J [EC:7.1.1.2] | all3840 | Oxidative phosphorylation | found only in BMAA-treated sample | 0.0940 | |
| 63 | inorganic pyrophosphatase [EC:3.6.1.1] | all3570* | Oxidative phosphorylation | 0.77 | 0.0806 | |
| Protein | The Fold Changes between the BMAA-Treated Samples and Control Sample * | |||
|---|---|---|---|---|
| Nitrogen Starvation Growth Conditions (Previous Study [23]) | Nitrogen-Replete Growth Conditions (Present Study) | |||
| Downregulation | Upregulation | Downregulation | Upregulation | |
| PII | 0.55 | 1.75 | ||
| rbcL | 0.67 | 1.67 | ||
| ccmK | 1.82 | |||
| ccmM | 0.64 | |||
| Urease subunit alpha | 0.82 | 1.59 | ||
| thioredoxin I | Control | |||
| thioredoxin reductase | 2.22 | |||
| S10 | 0.068 | 0.68 | ||
| RecA | 3.03 | 3.7 | ||
| nodM | 2.2 | 1.43 | ||
| Alr4505 | 6.67 | 3.57 | ||
| All1411 | 4.55 | 2.7 | ||
| Alr3297 | 0.67 | 1.29 | ||
| Asl4369 | Control | Control | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koksharova, O.A.; Butenko, I.O.; Pobeguts, O.V.; Safronova, N.A.; Govorun, V.M. Proteomic Insights into Starvation of Nitrogen-Replete Cells of Nostoc sp. PCC 7120 under β-N-Methylamino-L-Alanine (BMAA) Treatment. Toxins 2020, 12, 372. https://doi.org/10.3390/toxins12060372
Koksharova OA, Butenko IO, Pobeguts OV, Safronova NA, Govorun VM. Proteomic Insights into Starvation of Nitrogen-Replete Cells of Nostoc sp. PCC 7120 under β-N-Methylamino-L-Alanine (BMAA) Treatment. Toxins. 2020; 12(6):372. https://doi.org/10.3390/toxins12060372
Chicago/Turabian StyleKoksharova, Olga A., Ivan O. Butenko, Olga V. Pobeguts, Nina A. Safronova, and Vadim M. Govorun. 2020. "Proteomic Insights into Starvation of Nitrogen-Replete Cells of Nostoc sp. PCC 7120 under β-N-Methylamino-L-Alanine (BMAA) Treatment" Toxins 12, no. 6: 372. https://doi.org/10.3390/toxins12060372
APA StyleKoksharova, O. A., Butenko, I. O., Pobeguts, O. V., Safronova, N. A., & Govorun, V. M. (2020). Proteomic Insights into Starvation of Nitrogen-Replete Cells of Nostoc sp. PCC 7120 under β-N-Methylamino-L-Alanine (BMAA) Treatment. Toxins, 12(6), 372. https://doi.org/10.3390/toxins12060372