Monitoring Phenolic Compounds in Rice during the Growing Season in Relation to Fungal and Mycotoxin Contamination




Abstract
1. Introduction
2. Results and Discussion
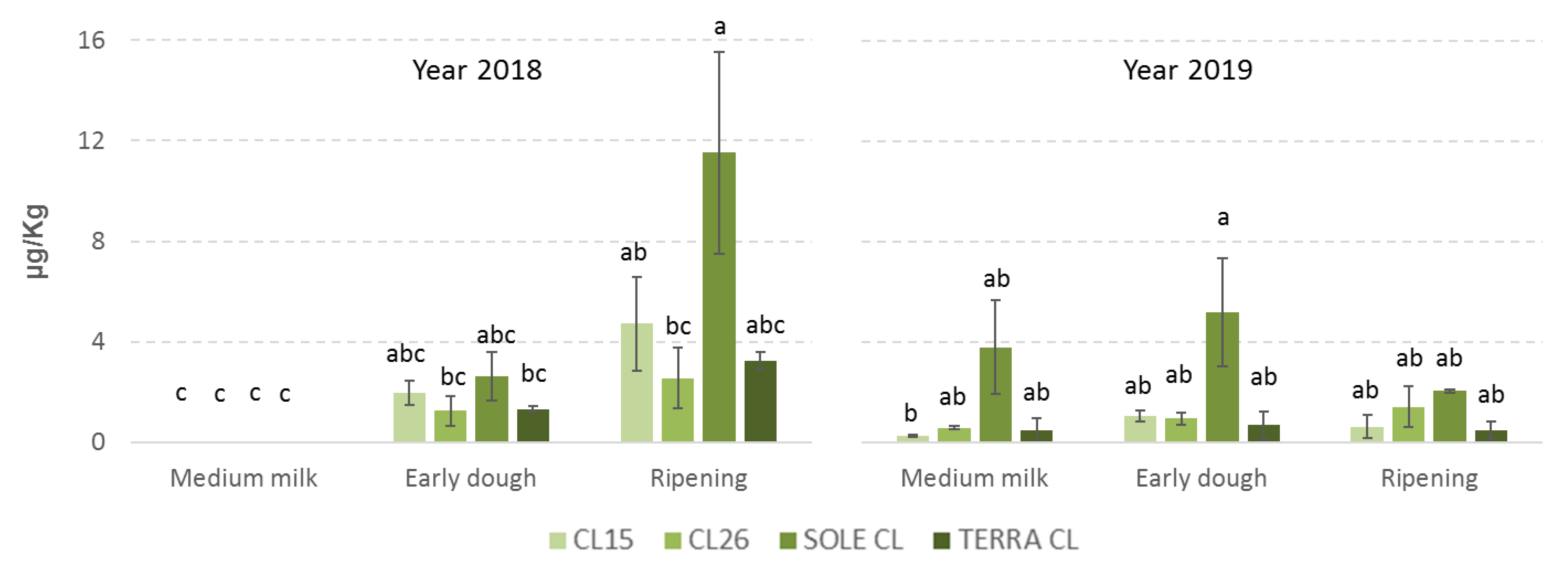
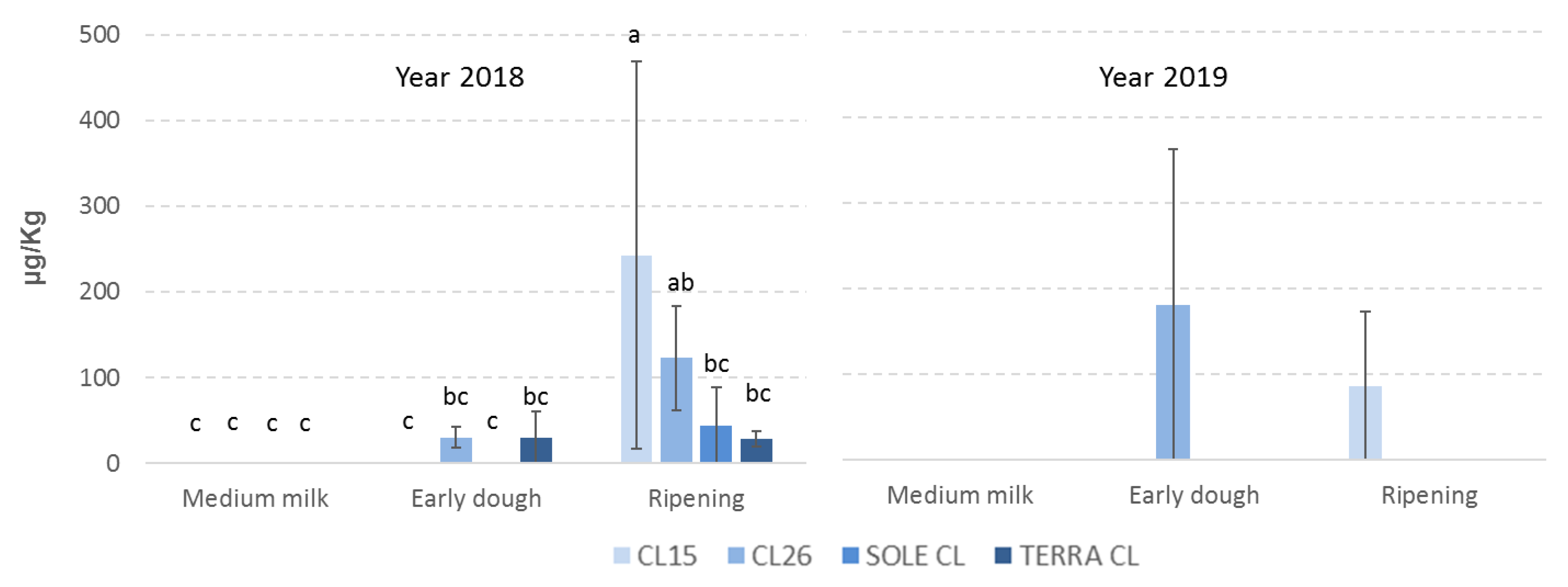
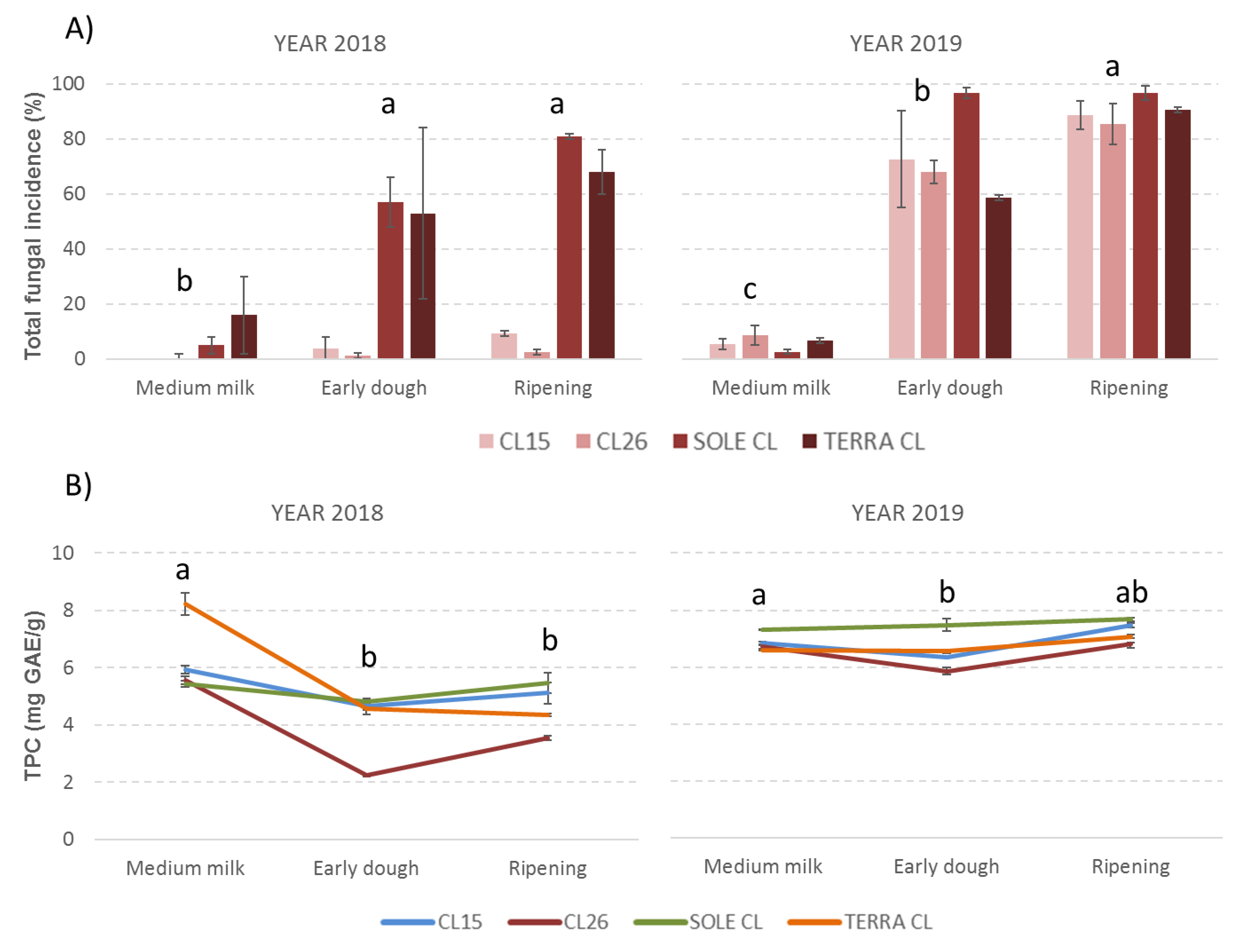
2.1. Fungal Incidence and Mycotoxin Contamination
2.2. Total Phenolic Content and Main Phenolic Composition
2.3. Correlations between Phenolic Acids, Fungal Contamination and Mycotoxin Content
3. Conclusions
4. Materials and Methods
4.1. Field Samples
4.2. Monitoring of Mycotoxigenic Fungi
4.3. Monitoring of Mycotoxins
4.4. Determination of Total Phenolic Content (TPC)
4.5. Determination of Phenolic Acids
4.6. Data Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shao, Y.; Xu, F.; Sun, X.; Bao, J.; Beta, T. Phenolic acids, anthocyanins, and antioxidant capacity in rice (Oryza sativa L.) grains at four stages of development after flowering. Food Chem. 2014, 143, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Bao, J. Polyphenols in whole rice grain: Genetic diversity and health benefits. Food Chem. 2015, 180, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Ente Nazionale Risi. 2018. Available online: www.enterisi.it (accessed on 3 March 2020).
- Nalley, L.L.; Tsiboe, F.; Durand-Morat, A.; Shew, A.M.; Thoma, G. Economic and Environmental Impact of Rice Blast Pathogen (Magnaporthe oryzae) Alleviation in the United States. PLoS ONE 2016, 11, e0167295. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Solanki, I.S.; Bashyal, B.M.; Singh, Y.; Srivastava, K. Bakanae of rice—An emerging disease in Asia. J. Anim. Plant Sci. 2015, 25, 1499–1514. [Google Scholar]
- Piotti, E.; Rigano, M.M.; Rodino, D.; Rodolfi, M.; Castiglione, S.; Am, P.; Sala, F. Genetic Structure of Pyricularia grisea (Cooke) Sacc. Isolates from Italian Paddy Fields. J. Phytopathol. 2005, 153, 80–86. [Google Scholar] [CrossRef]
- Bregaglio, S.; Donatelli, M.; Confalonieri, R. Fungal infections of rice, wheat, and grape in Europe in 2030–2050. Agron. Sustain. Dev. 2013, 33, 767–776. [Google Scholar] [CrossRef]
- Bertuzzi, T.; Romani, M.; Rastelli, S.; Giorni, P. Mycotoxins and Related Fungi in Italian Paddy Rice during the Growing Season and Storage. Toxins 2019, 11, 151. [Google Scholar] [CrossRef]
- Giorni, P.; Rolla, U.; Romani, M.; Mulazzi, A.; Bertuzzi, T. Efficacy of Azoxystrobin on Mycotoxins and Related Fungi in Italian Paddy Rice. Toxins 2019, 11, 310. [Google Scholar] [CrossRef]
- Esekhiagbe, M.; Uzuazokaro Agatemor, M.M.; Agatemor, C. Phenolic content and antimicrobial potentials of Xylopia aethiopica and Myristica argentea. J. Chem. Chem. Eng. 2009, 28, 159–162. [Google Scholar] [CrossRef]
- Djabali, S.; Barak, M. Impact of polyphenolic extracts on resistance to fungal contamination in dry bean grains. Afr. J. Biotechnol. 2013, 12, 2978–2985. [Google Scholar]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef] [PubMed]
- De Ascensao, A.R.; Dubery, I.A. Soluble and wall bound phenolic polymers in Musa acuminata roots exposed to elicitors from Fusarium oxysporum f. sp. cubense. Phytochemistry 2003, 63, 679–686. [Google Scholar] [CrossRef]
- Field, B.; Jordán, F.; Osbourn, A.; Field, B. First encounters? deployment of defence-related natural products by plants. New Phytol. 2006, 172, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Durazzo, A.; Lucarini, M.; Souto, E.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A concise overview on the chemistry, occurrence, and human health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef]
- Da Silva, B.; Souza, M.M.; Badiale-Furlong, E. Antioxidant and antifungal activity of phenolic compounds and their relation to aflatoxin B1 occurrence in soybeans (Glycine max L.). J. Sci. Food Agric. 2019, 100, 1256–1264. [Google Scholar] [CrossRef]
- Giordano, D.; Beta, T.; Reyneri, A.; Blandino, M. Changes in the Phenolic Acid Content and Antioxidant Activity during Kernel Development of Corn (Zea mays L.) and Relationship with Mycotoxin Contamination. Cereal Chem. J. 2017, 94, 315–324. [Google Scholar] [CrossRef]
- Walter, M.; Marchesan, E.; Massoni, P.F.S.; Da Silva, L.P.; Sartori, G.M.S.; Ferreira, R.B. Antioxidant properties of rice grains with light brown, red and black pericarp colors and the effect of processing. Food Res. Int. 2013, 50, 698–703. [Google Scholar] [CrossRef]
- Park, S.-Y.; Ha, S.-H.; Lim, S.-H.; Jung, J.Y.; Lee, S.M.; Yeo, Y.; Kim, J.K. Determination of phenolic acids in Korean rice (Oryza sativa L.) cultivars using gas chromatography-time-of-flight mass spectrometry. Food Sci. Biotechnol. 2012, 21, 1141–1148. [Google Scholar] [CrossRef]
- Pyrri, I.; Kapsanaki-Gotsi, E. Functional relations of airborne fungi to meteorological and pollution factors in a Mediterranean urban environment. Fungal Ecol. 2017, 30, 48–54. [Google Scholar] [CrossRef]
- Kang, D.-Y.; Cheon, K.-S.; Oh, J.; Oh, H.; Kim, S.; Kim, N.; Lee, E.; Choi, I.; Baek, J.; Kim, K.-H.; et al. Rice Genome Resequencing Reveals a Major Quantitative Trait Locus for Resistance to Bakanae Disease Caused by Fusarium fujikuroi. Int. J. Mol. Sci. 2019, 20, 2598. [Google Scholar] [CrossRef]
- Sandhu, G.S.; Dhaliwal, N. Incidence of Bakanae disease of basmati rice in south-western part of Punjab and its management. Int. J. Plant Prot. 2016, 9, 353–357. [Google Scholar] [CrossRef]
- Leggieri, M.C.; Van Der Fels-Klerx, H.; Battilani, P. Cross-validation of predictive models for deoxynivalenol in wheat at harvest. World Mycotoxin J. 2013, 6, 389–397. [Google Scholar] [CrossRef]
- Giorni, P.; Bertuzzi, T.; Battilani, P. Impact of Fungi Co-occurrence on Mycotoxin Contamination in Maize during the Growing Season. Front. Microbiol. 2019, 10, 1265. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, C.; Giorni, P.; Cirlini, M.; Reverberi, M.; Gregori, R.; Ludovici, M.; Camera, E.; Fanelli, C.; Battilani, P.; Scala, V. Maize lipids play a pivotal role in the fumonisin accumulation. World Mycotoxin J. 2015, 8, 87–97. [Google Scholar] [CrossRef]
- Schenck, J.; Müller, C.E.; Djurle, A.; Jensen, D.F.; O’Brien, M.; Johansen, A.; Rasmussen, P.H.; Spörndly, R. Occurrence of filamentous fungi and mycotoxins in wrapped forages in Sweden and Norway and their relation to chemical composition and management. Grass Forage Sci. 2019, 74, 613–625. [Google Scholar] [CrossRef]
- Hjelkrem, A.-G.R.; Aamot, H.U.; Brodal, G.; Strand, E.C.; Torp, T.; Edwards, S.; Dill-Macky, R.; Hofgaard, I.S. HT-2 and T-2 toxins in Norwegian oat grains related to weather conditions at different growth stages. Eur. J. Plant Pathol. 2018, 151, 501–514. [Google Scholar] [CrossRef]
- Marín, S.; Hodžić, I.; Ramos, A.J.; Sanchis, V. Predicting the growth/no-growth boundary and ochratoxin A production by Aspergillus carbonarius in pistachio nuts. Food Microbiol. 2008, 25, 683–689. [Google Scholar] [CrossRef]
- Battilani, P.; Giorni, P.; Bertuzzi, T.; Formenti, S.; Pietri, A. Black aspergilli and ochratoxin A in grapes in Italy. Int. J. Food Microbiol. 2006, 111, S53–S60. [Google Scholar] [CrossRef]
- Stanciu, O.; Juan, C.; Berrada, H.; Miere, D.; Loghin, F.; Mañes, J. Study on Trichothecene and Zearalenone Presence in Romanian Wheat Relative to Weather Conditions. Toxins 2019, 11, 163. [Google Scholar] [CrossRef]
- Latiff, N.A.; Mohd Din, A.R.J.; Zainul Alam, S.A.; Hanapi, S.Z.; Sarmidi, M.R. Quantification of polyphenol content, antioxidant properties and LC-MS/MS analysis in Malaysian indigenous rice cultivars (Oryza sativa L.). Agric. Nat. Resour. 2019, 53, 402–409. [Google Scholar]
- Shittu, H.O.; Aisagbonhi, E.; Obiazikwor, O.H. Plants’ innate defence mechanisms against phytopathogens. J. Microbiol. Biotechnol. Food Sci. 2019, 9, 314–319. [Google Scholar] [CrossRef]
- Mckeehen, J.D.; Busch, R.H.; Fulcher, R.G. Evaluation of wheat (Triticum aestivum L.) phenolic acids during grain development and their contribution to Fusarium resistance. J. Agric. Food Chem. 1999, 47, 1476–1482. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.G.; Gonçalves, L.M.; Prietto, L.; Hackbart, H.S.; Furlong, E.B. Antioxidant activity and enzyme inhibition of phenolic acids from fermented rice bran with fungus Rizhopus oryzae. Food Chem. 2014, 146, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Bashir, K.; Ishimaru, Y.; Shimo, H.; Kakei, Y.; Senoura, T.; Takahashi, R.; Sato, Y.; Sato, Y.; Uozumi, N.; Nakanishi, H.; et al. Rice phenolics efflux transporter 2 (PEZ2) plays an important role in solubilizing apoplasmic iron. Soil Sci. Plant Nutr. 2011, 57, 803–812. [Google Scholar] [CrossRef]
- Ricelli, A.; De Angelis, M.; Primitivo, L.; Righi, G.; Sappino, C.; Antonioletti, R. Role of Some Food-Grade Synthesized Flavonoids on the Control of Ochratoxin A in Aspergillus carbonarius. Molecules 2019, 24, 2553. [Google Scholar] [CrossRef] [PubMed]
- Morcia, C.; Tumino, G.; Ghizzoni, R.; Bara, A.; Salhi, N.; Terzi, V. In Vitro Evaluation of Sub-Lethal Concentrations of Plant-Derived Antifungal Compounds on FUSARIA Growth and Mycotoxin Production. Molecules 2017, 22, 1271. [Google Scholar] [CrossRef]
- Shen, Y.; Jin, L.; Xiao, P.; Lu, Y.; Bao, J. Total phenolics, flavonoids, antioxidant capacity in rice grain and their relations to grain color, size and weight. J. Cereal Sci. 2009, 49, 106–111. [Google Scholar] [CrossRef]
- Clewer, A.G.; Scarisbrick, D.H. Practical Statistics and Experimental Design for Plant and Crop Science; John Wiley & Sons: Chichester, UK, 2001. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Factor A: Year | |||||||||
| 2018 | 2019 | ||||||||
| Fusarium spp. (%) | n.s. | 9.73 ± 8.42 | 8.39 ± 7.07 | ||||||
| Aspergillus spp. (%) | n.s. | 0.70 ± 2.00 | 0.72 ± 1.71 | ||||||
| Penicillium spp. (%) | n.s. | 1.36 ± 4.28 | 0.78 ± 1.51 | ||||||
| Other fungi (%) | ** | 30.45 ± 32.70 | B | 46.94 ± 31.06 | A | ||||
| STC (µg/kg) | n.s. | 2.48 ± 3.50 | 1.47 ± 1.60 | ||||||
| DON (µg/kg) | * | 38.35 ± 99.30 | A | 22.33 ± 72.94 | B | ||||
| TPC (mg GAE/g) | ** | 2.725 ± 0.585 | A | 2.299 ± 0.191 | B | ||||
| Factor B: Rice Variety | |||||||||
| CL15 | CL26 | SOLE CL | TERRA CL | ||||||
| Fusarium spp. (%) | n.s. | 10.67 ± 9.57 | 5.38 ± 4.73 | 9.60 ± 7.77 | 9.47 ± 6.30 | ||||
| Aspergillus spp. (%) | ** | 0.13 ± 0.50 | B | 2.10 ± 2.63 | A | 0.67 ± 2.02 | B | 0.13 ± 0.50 | B |
| Penicillium spp. (%) | n.s. | 0.93 ± 1.91 | 0.46 ± 0.84 | 0.40 ± 0.80 | 2.13 ± 5.08 | ||||
| Other fungi (%) | * | 38.00 ± 27.07 | AB | 38.31 ± 29.41 | AB | 48.27 ± 35.54 | A | 37.87 ± 30.13 | B |
| STC (µg/kg) | ** | 1.44 ± 1.63 | B | 0.97 ± 0.77 | B | 4.20 ± 3.78 | A | 1.04 ± 1.02 | B |
| DON (µg/kg) | n.s. | 54.91 ± 121.45 | 51.58 ± 106.82 | 7.33 ± 21.95 | 9.75 ± 17.24 | ||||
| TPC (mg GAE/g) | n.s. | 2.426 ± 0.304 | 2.363 ± 0.434 | 2.547 ± 0.208 | 2.492 ± 0.661 | ||||
| Factor C: Sampling Time | |||||||||
| Medium Milk (BBCH 75) | Early Dough (BBCH 83) | Ripening (BBCH 89) | |||||||
| Fusarium spp. (%) | ** | 2.20 ± 3.16 | C | 10.11 ± 6.60 | B | 14.74 ± 6.43 | A | ||
| Aspergillus spp. (%) | ** | 0.00 ± 0.00 | B | 0.32 ± 0.73 | B | 1.86 ± 2.77 | A | ||
| Penicillium spp. (%) | n.s. | 1.00 ± 4.36 | 0.53 ± 1.43 | 1.47 ± 1.82 | |||||
| Other fungi (%) | ** | 2.70 ± 3.18 | C | 53.68 ± 19.73 | B | 67.68 ± 11.36 | A | ||
| STC (µg/kg) | ** | 0.64 ± 1.39 | B | 1.93 ± 1.68 | A | 3.38 ± 3.84 | A | ||
| DON (µg/kg) | ** | 0.00 ± 0.00 | B | 30.20 ± 90.40 | AB | 61.78 ± 120.41 | A | ||
| TPC (mg GAE/g) | ** | 2.633 ± 0.574 | A | 2.238 ± 0.219 | B | 2.501 ± 0.343 | A | ||
| Factors Interactions | |||||||||
| A × B | A × C | B × C | A × B × C | ||||||
| Fusarium spp. (%) | n.s. | n.s | n.s. | n.s. | |||||
| Aspergillus spp. (%) | n.s. | ** | ** | n.s. | |||||
| Penicillium spp. (%) | n.s. | n.s | n.s. | n.s. | |||||
| Other fungi (%) | n.s. | ** | n.s. | n.s. | |||||
| STC (µg/kg) | n.s. | ** | n.s. | n.s. | |||||
| DON (µg/kg) | n.s. | * | n.s. | n.s. | |||||
| TPC (mg GAE/g) | ** | ** | ** | ** | |||||
| 2018 | 2019 | ||||||
|---|---|---|---|---|---|---|---|
| Rice Variety | Phenolic Compound | Medium Milk (BBCH 75) mg/Kg | Early Dough (BBCH 83) mg/Kg | Ripening (BBCH 89) mg/Kg | Medium Milk (BBCH 75) mg/Kg | Early Dough (BBCH 83) mg/Kg | Ripening (BBCH 89) mg/Kg |
| CL15 | Chlorogenic acid | 9.69 ± 8.43 | 0.23 ± 0.05 | <0.01 | 53.41 ± 21.30 | 109.45 ± 34.93 | 2.74 ± 0.27 |
| Caffeic acid | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Syringic acid | 1.16 ± 0.56 | 0.43 ± 0.06 | 0.15 ± 0.10 | <0.01 | <0.01 | <0.01 | |
| 4-hydroxybenzoic acid | 11.83 ± 3.97 | 3.61 ± 0.27 | 2.13 ± 1.20 | 3.38 ± 0.19 | 1.75 ± 0.16 | 2.34 ± 0.23 | |
| p-Coumaric acid | 6.62 ± 1.71 | 1.39 ± 0.24 | 0.59 ± 0.17 | 2.31 ± 0.29 | 1.06 ± 0.27 | 1.24 ± 0.15 | |
| Ferulic acid | 4.13 ± 0.65 | 2.30 ± 0.02 | 2.59 ± 0.58 | 0.97 ± 0.40 | 0.80 ± 0.20 | 2.05 ± 0.31 | |
| Protocatecuic acid | 0.46 ± 0.01 | 0.27 ± 0.11 | 1.16 ± 0.99 | 0.14 ± 0.07 | 0.12 ± 0.04 | 0.92 ± 0.75 | |
| Gallic acid | 0.02 ± 0.02 | <0.01 | <0.01 | 0.28 ± 0.06 | 0.24 ± 0.02 | 0.27 ± 0.05 | |
| CL26 | Chlorogenic acid | 2.92 ± 0.59 | 0.20 ± 0.02 | 0.17 ± 0.03 | 152.35 ± 109.30 | 4.53 ± 4.54 | 15.88 ± 4.34 |
| Caffeic acid | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Syringic acid | 0.64 ± 0.19 | 0.49 ± 0.05 | <0.01 | <0.01 | <0.01 | 0.39 ± 0.33 | |
| 4-hydroxybenzoic acid | 6.09 ± 0.72 | 4.82 ± 0.20 | 5.05 ± 0.31 | 20.02 ± 5.68 | 0.75 ± 0.90 | 2.94 ± 0.39 | |
| p-Coumaric acid | 2.38 ± 0.38 | 1.25 ± 0.23 | 0.81 ± 0.14 | 1.91 ± 0.32 | 0.27±0.23 | 1.34 ± 0.34 | |
| Ferulic acid | 2.16 ± 0.16 | 1.69 ± 0.02 | 2.53 ± 0.18 | 0.23 ± 0.17 | 0.29 ± 0.25 | 1.50 ± 0.09 | |
| Protocatecuic acid | 0.26 ± 0.01 | <0.01 | <0.01 | 0.14 ± 0.10 | 0.02 ± 0.02 | 0.38 ± 0.01 | |
| Gallic acid | <0.01 | <0.01 | <0.01 | 0.26 ± 0.08 | 0.26 ± 0.07 | 0.17 ± 0.04 | |
| SOLE CL | Chlorogenic acid | 1.76 ± 1.52 | 0.35 ± 0.24 | 0.78 ± 0.57 | 102.78 ± 57.19 | 82.79 ± 61.39 | 13.17 ± 3.62 |
| Caffeic acid | 1.31 ± 0.99 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Syringic acid | 0.32 ± 0.05 | <0.01 | 0.72 ± 0.02 | <0.01 | <0.01 | <0.01 | |
| 4-hydroxybenzoic acid | 9.90 ± 2.27 | 5.78 ± 1.24 | 3.03 ± 0.07 | 3.57 ± 0.12 | 1.71 ± 0.23 | 2.42 ± 0.27 | |
| p-Coumaric acid | 5.37 ± 1.93 | 1.98 ± 1.98 | 0.81 ± 0.14 | 2.49 ± 0.04 | 1.63 ± 0.38 | 1.39 ± 0.35 | |
| Ferulic acid | 3.93 ± 1.71 | 2.77 ± 0.99 | 5.92 ± 3.69 | 1.97 ± 1.03 | 1.88 ± 0.21 | 2.16 ± 0.35 | |
| Protocatecuic acid | 0.33 ± 0.04 | 0.20 ± 0.01 | 0.17 ± 0.10 | 0.18 ± 0.08 | 0.20 ± 0.08 | 0.44 ± 0.06 | |
| Gallic acid | <0.01 | 0.08 ± 0.02 | <0.01 | 0.19 ± 0.01 | 0.21 ± 0.01 | 0.17 ± 0.01 | |
| TERRA CL | Chlorogenic acid | 14.46 ± 14.24 | 0.65 ± 0.11 | 0.05 ± 0.01 | 141.61 ± 31.00 | 75.74 ± 15.03 | 30.71 ± 8.03 |
| Caffeic acid | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| Syringic acid | 0.62 ± 0.14 | <0.01 | <0.01 | <0.01 | <0.01 | 0.29 ± 0.23 | |
| 4-hydroxybenzoic acid | 21.82 ± 5.93 | 3.80 ± 0.12 | 4.41 ± 0.55 | 2.84 ± 0.16 | 1.33 ± 0.10 | 2.54 ± 0.12 | |
| p-Coumaric acid | 8.45 ± 0.42 | 1.17 ± 0.38 | 0.98 ± 0.05 | 1.43 ± 0.17 | 0.69 ± 0.13 | 0.88 ± 0.03 | |
| Ferulic acid | 4.07 ± 1.32 | 2.38 ± 0.71 | 3.77 ± 1.79 | 1.05 ± 0.15 | 0.88 ± 0.08 | 2.14 ± 0.47 | |
| Protocatecuic acid | 1.04 ± 0.04 | 0.61 ± 0.32 | 0.46 ± 0.01 | 0.28 ± 0.04 | 0.20 ± 0.11 | 0.59 ± 0.02 | |
| Gallic acid | <0.01 | 0.01 ± 0.01 | <0.01 | 0.23 ± 0.02 | 0.18 ± 0.01 | 0.18 ± 0.01 | |
| Phenolic Acid | Chlorogenic Acid | Syringic Acid | 4-hydroxybenzoic Acid | p-Coumaric Acid | Ferulic Acid | Protocatecuic Acid | Gallic Acid |
|---|---|---|---|---|---|---|---|
| Chlorogenic acid | 1 | −0.294 * | −0.253 | 0.182 | −0.565 ** | −0.198 | 0.626 ** |
| Syringic acid | 1 | −0.350 ** | 0.190 | 0.447 ** | 0.159 | −0.486 ** | |
| 4-hydroxybenzoic acid | 1 | 0.653 ** | 0.387 ** | 0.118 | −0.481 ** | ||
| p-Coumaric acid | 1 | 0.270 * | 0.153 | −0.080 | |||
| Ferulic acid | 1 | 0.547 ** | −0.664 ** | ||||
| Protocatecuic acid | 1 | −0.296 * | |||||
| Gallic acid | 1 |
| Phenolic Acid | Fusarium spp. Incidence | Aspergillus spp. Incidence | Penicillium spp. Incidence | Total Fungi Incidence | STC | DON |
|---|---|---|---|---|---|---|
| Chlorogenic acid | −0.331 * | −0.285 * | −0.243 | −0.141 | −0.281 | −0.576 ** |
| Syringic acid | −0.156 | 0.171 | 0.003 | −0.151 | −0.230 | −0.055 |
| 4-hydroxybenzoic acid | −0.348 ** | −0.019 | −0.054 | −0.581 ** | −0.314 * | −0.059 |
| p-Coumaric acid | −0.497 ** | −0.269 * | −0.140 | −0.532 ** | −0.427 ** | −0.466 ** |
| Ferulic acid | 0.203 | 0.116 | 0.145 | 0.039 | 0.101 | 0.206 |
| Protocatecuic acid | 0.237 | −0.026 | 0.355 ** | 0.156 | −0.255 | −0.021 |
| Gallic acid | −0.068 | −0.148 | 0.043 | 0.138 | −0.102 | −0.183 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giorni, P.; Rastelli, S.; Fregonara, S.; Bertuzzi, T. Monitoring Phenolic Compounds in Rice during the Growing Season in Relation to Fungal and Mycotoxin Contamination. Toxins 2020, 12, 341. https://doi.org/10.3390/toxins12050341
Giorni P, Rastelli S, Fregonara S, Bertuzzi T. Monitoring Phenolic Compounds in Rice during the Growing Season in Relation to Fungal and Mycotoxin Contamination. Toxins. 2020; 12(5):341. https://doi.org/10.3390/toxins12050341
Chicago/Turabian StyleGiorni, Paola, Silvia Rastelli, Sofia Fregonara, and Terenzio Bertuzzi. 2020. "Monitoring Phenolic Compounds in Rice during the Growing Season in Relation to Fungal and Mycotoxin Contamination" Toxins 12, no. 5: 341. https://doi.org/10.3390/toxins12050341
APA StyleGiorni, P., Rastelli, S., Fregonara, S., & Bertuzzi, T. (2020). Monitoring Phenolic Compounds in Rice during the Growing Season in Relation to Fungal and Mycotoxin Contamination. Toxins, 12(5), 341. https://doi.org/10.3390/toxins12050341