Assessing the Effect of Mycotoxin Combinations: Which Mathematical Model Is (the Most) Appropriate?




Abstract
1. Introduction
2. Mathematical Models for Assessing Mycotoxin Interactions
2.1. Simple Addition of Effects
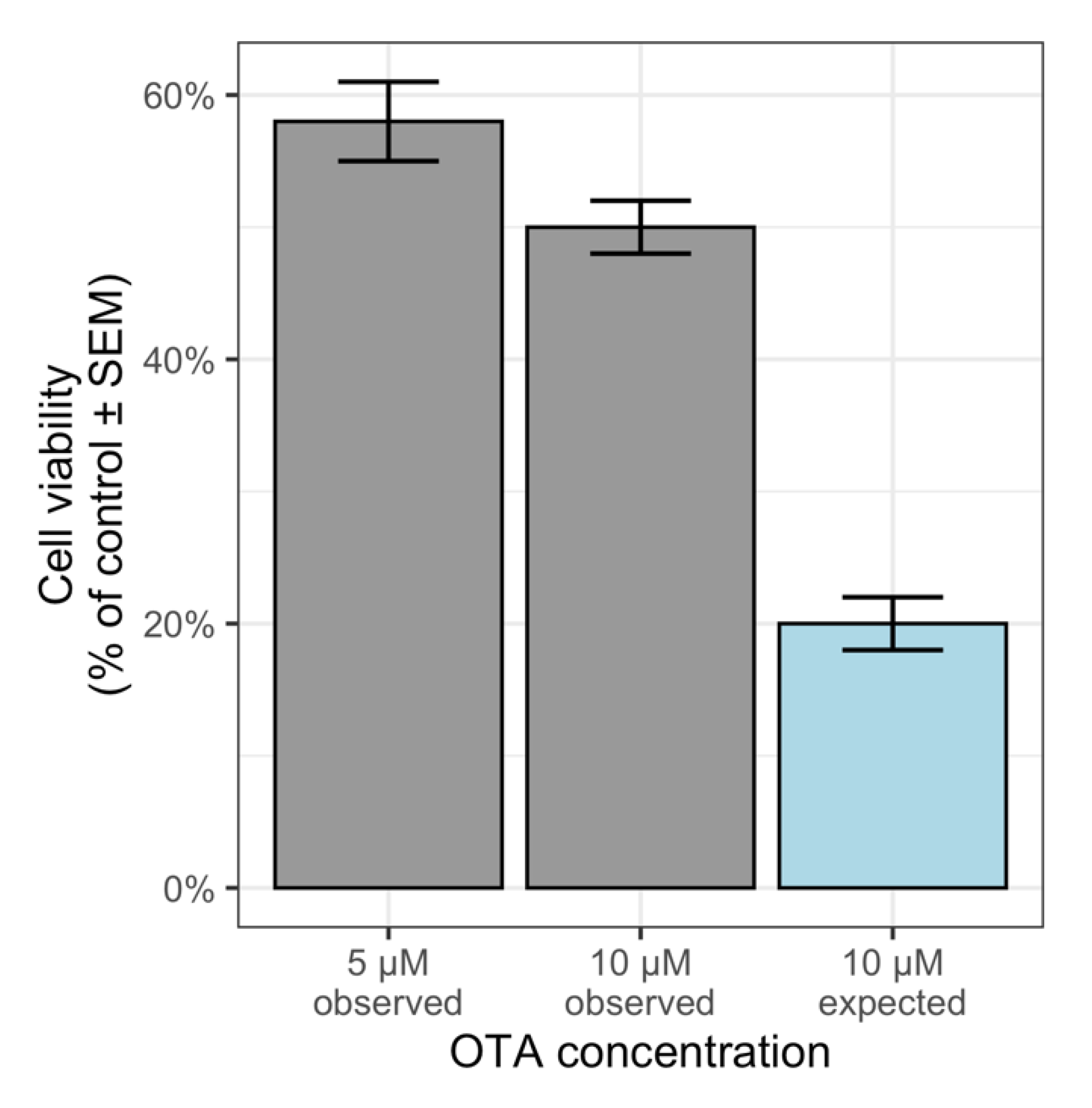
2.2. Factorial Analysis of Variance
2.3. Bliss Independence Criterion
2.4. Loewe’s Additivity Law
2.5. Response Surface
2.6. Highest Single Agent (HSA) Model
2.7. Combination Index and Isobologram Analysis
2.8. Chou and Talalay’s Median Effect Approach
2.9. MixLow Method
2.10. CISNE (Code for the Identification of Synergism Numerically Efficient)
2.11. Other methods
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pitt, J.I.; David Miller, J. A Concise History of Mycotoxin Research. J. Agric. Food Chem. 2017, 65, 7021–7033. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, N.; Keller, N.P. Mycotoxins in conversation with bacteria and fungi. Front. Microbiol. 2019, 10, 403. [Google Scholar] [CrossRef]
- Fox, E.M.; Howlett, B.J. Secondary metabolism: Regulation and role in fungal biology. Curr. Opin. Microbiol. 2008, 11, 481–487. [Google Scholar] [CrossRef]
- Van Egmond, H.P.; Schothorst, R.C.; Jonker, M.A. Regulations relating to mycotoxins in food. Anal. Bioanal. Chem. 2007, 389, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Assunção, R.; Silva, M.J.; Alvito, P. Challenges in risk assessment of multiple mycotoxins in food. World Mycotoxin J. 2016, 9, 791–811. [Google Scholar] [CrossRef]
- Abdallah, M.F.; Girgin, G.; Baydar, T.; Krska, R.; Sulyok, M. Occurrence of multiple mycotoxins and other fungal metabolites in animal feed and maize samples from Egypt using LC-MS/MS. J. Sci. Food Agric. 2017, 97, 4419–4428. [Google Scholar] [CrossRef] [PubMed]
- Malachová, A.; Sulyok, M.; Beltrán, E.; Berthiller, F.; Krska, R. Optimization and validation of a quantitative liquid chromatography-tandem mass spectrometric method covering 295 bacterial and fungal metabolites including all regulated mycotoxins in four model food matrices. J. Chromatogr. A 2014, 1362, 145–156. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging Mycotoxins: Beyond Traditionally Determined Food Contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [Google Scholar] [CrossRef]
- Kovač, M.; Šubarić, D.; Bulaić, M.; Kovač, T.; Šarkanj, B. Yesterday masked, today modified; what do mycotoxins bring next? Arh. Hig. Rada Toksikol. 2018, 69, 196–214. [Google Scholar] [CrossRef]
- Ibáñez-Vea, M.; González-Peñas, E.; Lizarraga, E.; López de Cerain, A. Co-occurrence of aflatoxins, ochratoxin A and zearalenone in barley from a northern region of Spain. Food Chem. 2012, 132, 35–42. [Google Scholar] [CrossRef]
- Santini, A.; Meca, G.; Uhlig, S.; Ritieni, A. Fusaproliferin, beauvericin and enniatins: Occurrence in food-A review. World Mycotoxin J. 2012, 5, 71–81. [Google Scholar] [CrossRef]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O.; et al. Current situation of mycotoxin contamination and co-occurrence in animal feed focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [PubMed]
- Vaclavikova, M.; Malachova, A.; Veprikova, Z.; Dzuman, Z.; Zachariasova, M.; Hajslova, J. “Emerging” mycotoxins in cereals processing chains: Changes of enniatins during beer and bread making. Food Chem. 2013, 136, 750–757. [Google Scholar] [CrossRef]
- Kademi, H.I.; Baba, I.A.; Saad, F.T. Modelling the dynamics of toxicity associated with aflatoxins in foods and feeds. Toxicol. Rep. 2017, 4, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marín, S. Predicting mycotoxins in foods: A review. Food Microbiol. 2009, 26, 757–769. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Yoon, Y.; Kim, D.M.; Kim, D.S.; Park, K.H.; Chun, H.S. Mathematical models to predict kinetic behavior and aflatoxin production of Aspergillus flavus under various temperature and water activity conditions. Food Sci. Biotechnol. 2014, 23, 975–982. [Google Scholar] [CrossRef]
- Smith, M.C.; Madec, S.; Coton, E.; Hymery, N. Natural Co-occurrence of mycotoxins in foods and feeds and their in vitro combined toxicological effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Schatzmayr, G.; Taranu, I.; Marin, D.; Puel, O.; Oswald, I.P. Mycotoxins co-contamination: Methodological aspects and biological relevance of combined toxicity studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 3489–3507. [Google Scholar] [CrossRef]
- Grenier, B.; Oswald, I. Mycotoxin co-contamination of food and feed: Meta-analysis of publications describing toxicological interactions. World Mycotoxin J. 2011, 4, 285–313. [Google Scholar] [CrossRef]
- Šegvić Klarić, M. Adverse effects of combined mycotoxins. Arh. Hig. Rada Toksikol. 2012, 63, 519–530. [Google Scholar] [CrossRef]
- Šegvić Klarić, M.; Rašić, D.; Peraica, M. Deleterious effects of mycotoxin combinations involving ochratoxin A. Toxins 2013, 5, 1965–1987. [Google Scholar]
- Tekin, E.; Beppler, C.; White, C.; Mao, Z.; Savage, V.M.; Yeh, P.J. Enhanced identification of synergistic and antagonistic emergent interactions among three or more drugs. J. R. Soc. Interface 2016, 13, 18–20. [Google Scholar] [CrossRef] [PubMed]
- Foucquier, J.; Guedj, M. Analysis of drug combinations: Current methodological landscape. Pharmacol. Res. Perspect. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Weber, F.; Freudinger, R.; Schwerdt, G.; Gekle, M. A rapid screening method to test apoptotic synergisms of ochratoxin A with other nephrotoxic substances. Toxicol. Vitr. 2005, 19, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Caudle, R.M.; Williams, G.M. The misuse of analysis of variance to detect synergy in combination drug studies. Pain 1993, 55, 313–317. [Google Scholar] [CrossRef]
- Bliss, C.I. The toxicity of poisons applied jointly. Ann. Appl. Biol. 1939, 26, 585–615. [Google Scholar] [CrossRef]
- Loewe, S.; Muischnek, H. Über Kombinationswirkungen. Arch. Für Exp. Pathol. Und Pharmakologie 1926, 114, 313–326. [Google Scholar] [CrossRef]
- Jonker, M.J.; Svendsen, C.; Bedaux, J.J.M.; Bongers, M.; Kammenga, J.E. Significance testing of synergistic/antagonistic, dose level–dependent, or dose ratio–dependent effects in mixture dose–response analysis. Environ. Toxicol. Chem. 2005, 24, 2701. [Google Scholar] [CrossRef]
- Berenbaum, M.C. Criteria for analyzing interactions between biologically active agents. Adv. Cancer Res. 1981, 35, 269–335. [Google Scholar]
- Chou, T.-C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef]
- Boik, J.C.; Newman, R.A.; Boik, R.J. Quantifying synergism/antagonism using nonlinear mixed-effects modeling: A simulation study. Stat. Med. 2008, 27, 1040–1061. [Google Scholar] [CrossRef] [PubMed]
- Berenbaum, M.C. What is synergy? Pharmacol. Rev. 1989, 41, 93–141. [Google Scholar]
- García-Fuente, A.; Vázquez, F.; Viéitez, J.M.; García Alonso, F.J.; Martín, J.I.; Ferrer, J. CISNE: An accurate description of dose-effect and synergism in combination therapies. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Šegvić Klarić, M.; Daraboš, D.; Rozgaj, R.; Kašuba, V.; Pepeljnjak, S. Beauvericin and ochratoxin A genotoxicity evaluated using the alkaline comet assay: Single and combined genotoxic action. Arch. Toxicol. 2010, 84, 641–650. [Google Scholar] [CrossRef]
- Huang, X.; Gao, Y.; Li, S.; Wu, C.; Wang, J.; Zheng, N. Modulation of Mucin (MUC2, MUC5AC AND MUC5B) mRNA expression and protein production and secretion in Caco-2/HT29-MTX co-cultures following exposure to individual and combined aflatoxin M1 and ochratoxin A. Toxins 2019, 11, 132. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Liu, Y.; Zhang, G. Interaction of aflatoxin B1 and fumonisin B1 in HepG2 cell apoptosis. Food Biosci. 2017, 20, 131–140. [Google Scholar] [CrossRef]
- Zheng, N.; Gao, Y.N.; Liu, J.; Wang, H.W.; Wang, J.Q. Individual and combined cytotoxicity assessment of zearalenone with ochratoxin A or α-zearalenol by full factorial design. Food Sci. Biotechnol. 2018, 27, 251–259. [Google Scholar] [CrossRef]
- Smith, M.-C.; Gheux, A.; Coton, M.; Madec, S.; Hymery, N.; Coton, E. In vitro co-culture models to evaluate acute cytotoxicity of individual and combined mycotoxin exposures on Caco-2, THP-1 and HepaRG human cell lines. Chem. Biol. Interact. 2018, 281, 51–59. [Google Scholar] [CrossRef]
- Smith, M.C.; Hymery, N.; Troadec, S.; Pawtowski, A.; Coton, E.; Madec, S. Hepatotoxicity of fusariotoxins, alone and in combination, towards the HepaRG human hepatocyte cell line. Food Chem. Toxicol. 2017, 109, 439–451. [Google Scholar] [CrossRef]
- Li, Y.; Wang, T.Q.; Wu, J.; Zhang, X.L.; Xu, Y.Y.; Qian, Y.Z. Multi-parameter analysis of combined hepatotoxicity induced by mycotoxin mixtures in HepG2 cells. World Mycotoxin J. 2018, 11, 225–235. [Google Scholar] [CrossRef]
- Vejdovszky, K.; Warth, B.; Sulyok, M.; Marko, D. Non-synergistic cytotoxic effects of Fusarium and Alternaria toxin combinations in Caco-2 cells. Toxicol. Lett. 2016, 241, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tallarida, R.J. Quantitative methods for assessing drug synergism. Genes Cancer 2011, 2, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Geary, N. Understanding synergy. Am. J. Physiol. Metab. 2013, 304, E237–E253. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, B.; He, X.; Cheng, W.H.; Xu, W.; Luo, Y.; Liang, R.; Luo, H.; Huang, K. Analysis of individual and combined effects of ochratoxin a and zearalenone on HepG2 and KK-1 cells with mathematical models. Toxins 2014, 6, 1177–1192. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.Y.; Cedergreen, N.; Yiannikouris, A.; Swamy, H.V.L.N.; Karrow, N.A. Assessing interactions of binary mixtures of Penicillium mycotoxins (PMs) by using a bovine macrophage cell line (BoMacs). Toxicol. Appl. Pharmacol. 2017, 318, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Assunção, R.; Pinhão, M.; Loureiro, S.; Alvito, P.; Silva, M.J. A multi-endpoint approach to the combined toxic effects of patulin and ochratoxin a in human intestinal cells. Toxicol. Lett. 2019, 313, 120–129. [Google Scholar] [CrossRef]
- Vejdovszky, K.; Sack, M.; Jarolim, K.; Aichinger, G.; Somoza, M.M.; Marko, D. In vitro combinatory effects of the Alternaria mycotoxins alternariol and altertoxin II and potentially involved miRNAs. Toxicol. Lett. 2017, 267, 45–52. [Google Scholar] [CrossRef]
- Solhaug, A.; Karlsøen, L.M.; Holme, J.A.; Kristoffersen, A.B.; Eriksen, G.S. Immunomodulatory effects of individual and combined mycotoxins in the THP-1 cell line. Toxicol. Vitr. 2016, 36, 120–132. [Google Scholar] [CrossRef]
- Tallarida, R.J. Revisiting the isobole and related quantitative methods for assessing drug synergism. J. Pharmacol. Exp. Ther. 2012, 342, 2–8. [Google Scholar] [CrossRef]
- Anastasiadi, M.; Polizzi, K.; Lambert, R.J.W. An improved model for the analysis of combined antimicrobials: A replacement for the Chou-Talalay combination index method. J. Appl. Microbiol. 2018, 124, 97–107. [Google Scholar] [CrossRef]
- Vejdovszky, K.; Hahn, K.; Braun, D.; Warth, B.; Marko, D. Synergistic estrogenic effects of Fusarium and Alternaria mycotoxins in vitro. Arch. Toxicol. 2017, 91, 1447–1460. [Google Scholar] [CrossRef] [PubMed]
- Aupanun, S.; Phuektes, P.; Poapolathep, S.; Alassane-Kpembi, I.; Oswald, I.P.; Poapolathep, A. Individual and combined cytotoxicity of major trichothecenes type B, deoxynivalenol, nivalenol, and fusarenon-X on Jurkat human T cells. Toxicon 2019, 160, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Alassane-Kpembi, I.; Puel, O.; Pinton, P.; Cossalter, A.M.; Chou, T.C.; Oswald, I.P. Co-exposure to low doses of the food contaminants deoxynivalenol and nivalenol has a synergistic inflammatory effect on intestinal explants. Arch. Toxicol. 2017, 91, 2677–2687. [Google Scholar] [CrossRef] [PubMed]
- Ferreira Lopes, S.; Vacher, G.; Ciarlo, E.; Savova-Bianchi, D.; Roger, T.; Niculita-Hirzel, H. Primary and Immortalized Human Respiratory Cells Display Different Patterns of Cytotoxicity and Cytokine Release upon Exposure to Deoxynivalenol, Nivalenol and Fusarenon-X. Toxins 2017, 9, 337. [Google Scholar] [CrossRef]
- Zhou, H.; George, S.; Li, C.; Gurusamy, S.; Sun, X.; Gong, Z.; Qian, H. Combined toxicity of prevalent mycotoxins studied in fish cell line and zebrafish larvae revealed that type of interactions is dose-dependent. Aquat. Toxicol. 2017, 193, 60–71. [Google Scholar] [CrossRef]
- Zouaoui, N.; Mallebrera, B.; Berrada, H.; Abid-Essefi, S.; Bacha, H.; Ruiz, M.J. Cytotoxic effects induced by patulin, sterigmatocystin and beauvericin on CHO-K1 cells. Food Chem. Toxicol. 2016, 89, 92–103. [Google Scholar] [CrossRef]
- Juan-García, A.; Tolosa, J.; Juan, C.; Ruiz, M.-J. Cytotoxicity, genotoxicity and disturbance of cell cycle in HepG2 Cells exposed to OTA and BEA: Single and combined actions. Toxins 2019, 11, 341. [Google Scholar] [CrossRef]
- Lei, Y.; Guanghui, Z.; Xi, W.; Yingting, W.; Xialu, L.; Fangfang, Y.; Goldring, M.B.; Xiong, G.; Lammi, M.J. Cellular responses to T-2 toxin and/or deoxynivalenol that induce cartilage damage are not specific to chondrocytes. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, S.; Tan, Y.; Liu, N.; Wu, A. Individual and combined cytotoxic effects of co-occurring deoxynivalenol family mycotoxins on human gastric epithelial cells. Toxins 2017, 9, 96. [Google Scholar] [CrossRef]
- Zhou, H.; George, S.; Hay, C.; Lee, J.; Qian, H.; Sun, X. Individual and combined effects of aflatoxin B1, deoxynivalenol and zearalenone on HepG2 and RAW 264.7 cell lines. Food Chem. Toxicol. 2017, 103, 18–27. [Google Scholar] [CrossRef]
- Gao, Y.N.; Wang, J.Q.; Li, S.L.; Zhang, Y.D.; Zheng, N. Aflatoxin M1 cytotoxicity against human intestinal Caco-2 cells is enhanced in the presence of other mycotoxins. Food Chem. Toxicol. 2016, 96, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Pistol, G.C.; Bulgaru, C.V.; Taranu, I. Cytotoxic and inflammatory effects of individual and combined exposure of HepG2 cells to zearalenone and its metabolites. Naunyn. Schmiedebergs. Arch. Pharmacol. 2019, 392, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Juan-García, A.; Juan, C.; Manyes, L.; Ruiz, M.J. Binary and tertiary combination of alternariol, 3-acetyl-deoxynivalenol and 15-acetyl-deoxynivalenol on HepG2 cells: Toxic effects and evaluation of degradation products. Toxicol. Vitr. 2016, 34, 264–273. [Google Scholar] [CrossRef]
- Sobral, M.M.C.; Faria, M.A.; Cunha, S.C.; Ferreira, I.M.P.L.V.O. Toxicological interactions between mycotoxins from ubiquitous fungi: Impact on hepatic and intestinal human epithelial cells. Chemosphere 2018, 202, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Blanco, C.; Elmo, L.; Waldner, T.; Ruiz, M.J. Cytotoxic effects induced by patulin, deoxynivalenol and toxin T2 individually and in combination in hepatic cells (HepG2). Food Chem. Toxicol. 2018, 120, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Kong, M. Confidence Intervals of Interaction Index for Assessing Multiple Drug Interaction. Stat. Biopharm. Res. 2009, 1, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Shao, W.; Yu, F.; Xing, K.; Liu, H.; Zhang, F.; Goldring, M.B.; Lammi, M.J.; Guo, X. Individual and combined toxicity of T-2 toxin and deoxynivalenol on human C-28/I2 and rat primary chondrocytes. J. Appl. Toxicol. 2019, 39, 343–353. [Google Scholar] [CrossRef]
- Boik, J.C.; Narasimhan, B. An R package for assessing drug synergism/antagonism. J. Stat. Softw. 2010, 34, 1–18. [Google Scholar] [CrossRef][Green Version]
- CISNE Code for the Identification of Synergism Numerically Efficient. Available online: https://cisnecode.github.io (accessed on 29 February 2020).
- Ren, Z.; Deng, H.; Deng, Y.; Liang, Z.; Deng, J.; Zuo, Z.; Hu, Y.; Shen, L.; Yu, S.; Cao, S. Combined effects of deoxynivalenol and zearalenone on oxidative injury and apoptosis in porcine splenic lymphocytes in vitro. Exp. Toxicol. Pathol. 2017, 69, 612–617. [Google Scholar] [CrossRef]
- Gayathri, L.; Karthikeyan, B.S.; Rajalakshmi, M.; Dhanasekaran, D.; Li, A.P.; Akbarsha, M.A. Metabolism-dependent cytotoxicity of citrinin and ochratoxin A alone and in combination as assessed adopting integrated discrete multiple organ co-culture (IdMOC). Toxicol. Vitr. 2018, 46, 166–177. [Google Scholar] [CrossRef]
- Gong, L.; Zhu, H.; Li, T.; Ming, G.; Duan, X.; Wang, J.; Jiang, Y. Molecular signatures of cytotoxic effects in human embryonic kidney 293 cells treated with single and mixture of ochratoxin A and citrinin. Food Chem. Toxicol. 2019, 123, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Juan-García, A.; Juan, C.; Tolosa, J.; Ruiz, M.J. Effects of deoxynivalenol, 3-acetyl-deoxynivalenol and 15-acetyl-deoxynivalenol on parameters associated with oxidative stress in HepG2 cells. Mycotoxin Res. 2019, 35, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Juan-García, A.; Taroncher, M.; Font, G.; Ruiz, M.J. Micronucleus induction and cell cycle alterations produced by deoxynivalenol and its acetylated derivatives in individual and combined exposure on HepG2 cells. Food Chem. Toxicol. 2018, 118, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.C.; Madec, S.; Pawtowski, A.; Coton, E.; Hymery, N. Individual and combined toxicological effects of deoxynivalenol and zearalenone on human hepatocytes in in vitro chronic exposure conditions. Toxicol. Lett. 2017, 280, 238–246. [Google Scholar] [CrossRef]
- Springler, A.; Vrubel, G.J.; Mayer, E.; Schatzmayr, G.; Novak, B. Effect of Fusarium-derived metabolites on the barrier integrity of differentiated intestinal porcine epithelial cells (IPEC-J2). Toxins 2016, 8, 345. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Mycotoxin Combination | In Vitro Model | Mathematical Model Applied for the Endpoint | Endpoint Combined Effect | Reference |
|---|---|---|---|---|
| AFM1 + OTA | Caco-2/ human colon HT29-MTX co-cultures (100/0, 90/10, 75/25 and 0/100) | Simple addition of effects | Cell viability (Enhanced Cell Counting Kit-8, CCK-8): synergism in all cultures TEER: antagonism in all cultures, except additive effect in 90/10 co-culture Intestinal mucin MUC2 and MUC5B mRNA expression: synergistic effect in 75/25 and 0/100 cultures at 4 μg/mL additive effects at the low concentration (0.05 μg/mL) culture, antagonistic effects in 100/0 and 90/10 cultures at 4 μg/mL Intestinal mucin MUC5AC mRNA expression: antagonistic effect in 100/0 cultures, an additive effect in 0/100 cultures at two concentrations of the mixtures Intestinal mucin MUC5AC, MUC2 AND MUC5B on protein level: synergism at 0.05 and 4 μg/mL additive effect at 0.05 μg/mL in 75/25 and 90/10 cultures | [35] |
| AFB1 + FB1 | HepG2 cells | Simple addition of effects and factorial analysis (two-way ANOVA) | Cell cycle analysis (flow citometry assay): synergism on apoptosis at 10% and 30% of IC50 | [36] |
| ZEN (30 or 60 µM) + OTA (6 or 12 µM) ZEN (30 or 60 µM) + α-ZEL (15 or 30 µM) | HepG2 cells | Full factorial analysis: 3 × 3 two-way ANOVA matrix | Cytotoxicity (MTT test): synergism of ZEN (60 µM) + α-ZEL (15 or 30 µM) antagonism in all other combinations Oxidative stress parameters (MDA, GSH, Gpx, SOD): synergism of ZEN (60 µM) + α-ZEL (15 or 30 µM) antagonism in all other combinations | [37] |
| DON + ZEN | Bi- and tri-culture systems: A) Caco-2 and HepaRG; B) Caco-2 and THP-1; C) HepaRG and THP-1 D) Caco-2, HepaRG and THP-1 | Response additivity, CIRA) and Bliss independence criterion (independent joint action, CIIjA); IC10 (1:1) and IC30 (1:1) | Cytotoxicity (MTS test): additive effect for combination of IC10 in A–D (CIRA and CIIjA) synergism for combination of IC30 in A–C (CIRA and CIIjA) additive effect for combination of IC30 in D (CIRA and CIIjA) | [38] |
| DON + MON DON + FB1 DON + ZEN NIV + T-2 | HepaRG cells | Response additivity (CIRA) and Bliss independence criterion (independent joint action, CIIjA) | Cytotoxicity (MTS): synergism of DON + MON in all combinations except additive effect at highest concentration (1:1) (CIRA and CIIjA) synergism of DON + FB1 in all combinations (CIRA and CIIjA) except additive effect at highest concentration (1:1) (CIRA) synergism of DON + ZEN at low and medium concentrations (CIRA and CIIjA); additive effect (CIIJA) and antagonism at high concentrations (CRA) NIV + T-2 synergism at low concentrations (CIRA and CIIjA); additive effect or antagonism (CIIJA) and antagonism at medium and high concentrations (CIRA) | [39] |
| AFB1 + ZEN AFB1 + DON ZEN + DON AFB1 + ZEN + DON | HepG2 cells | Bliss independence criterion (IA) and Loewe additivity models (CA); CI-Isobologram method | Cell number (high content analysis by fluorescent labelling: IA and CA model: deviation from the obtained results; better consistency was achieved by CA model; CI model: antagonism at low fraction affected (0.05–0.15) changing to additive and synergistic effect as fraction affected increases for all combinations | [40] |
| TeA + ENN B; TeA + ZEN; TeA + DON; TeA + NIV; TeA + AURO; ENN B + ZEN; ENN B + DON; ENN B + NIV ENN B + AURO; ZEN + DON; ZEN + NIV; ZEN + AURO; DON + NIV; DON + AURO | Caco-2 cells | Bliss independence criterion combined with CI calculated by Chou (C) and Chou-Talalay (CT) method | Cytotxicity (WST-1 test): additive effects of binary mixtures at low concentrations calculated by Bliss independence criterion antagonism of binary mixtures ENN B, ZEN and DON as well as binary combinations of Fusarium toxins with TeA applied at cytotoxic concentrations as calculated by CI | [41] |
| ATX II + AOH | HepG2, HT29 cells and human corneal epithelial HCEC cells | Bliss independence criterion, constant ratio of 1:10 or 1:1 | Cytotoxicity (WST-1 test): dominant additive effect in all cell lines antagonism in specific doses of ratios 1:10 or 1:1 | [47] |
| AOH + DON AOH + ZEN ZEN + DON AOH + DON + ZEN | THP-1 monocytes differentiated into macrophages | Concentration addition (CA) and independent action (IA) model at equal effect concentration | CD14 expression: synergism of AOH + DON applied at low concentrations additive effects of binary and tertiary mixtures of AOH, ZEN and DON, as calculated by both CA and IA | [48] |
| CIT + OTA OTA + PAT OTA + MPA OTA + PA CIT + PAT CIT + MPA CIT + PA PAT + MPA PAT + PA MPA + PA | Bovine peritoneal macrophage BoMacs cells | CA and IA model; Penicillium toxins in IC25, ½ IC25 and ¼ IC25 | Cell proliferation (CyQUANT® GR dye): CIT + OTA synergism at ½ IC25 (CA, IA) OTA + PAT additive effects (CA, IA) OTA + MPA synergism at IC25, ½ IC25 and ¼ IC25 (CA) OTA + PA synergism at IC25 and ¼ IC25 (CA) - CIT + PAT antagonism at ½ IC25 (CA) CIT + MPA inconclusive (synergism CA, antagonism IA) CIT + PA antagonism at IC25, ½ IC25 (IA) PAT + MPA antagonism at IC25, ½ IC25 and ¼ IC25 (IA) PAT + PA synergism at ½ IC25; antagonism at IC25 MPA + PA inconclusive | [45] |
| OTA + PAT | Caco-2 cells | Concentration addition model (CA) and independent action (IA) model with Jonker’s generalization [28] | Cytotoxicity (MTT test): - additive effects (CA) synergism at high concentration of OTA and low of PAT (IA) antagonism at high concentration of PAT and low of OTA (IA) Gastrointestinal barrier integrity (TEER assay): synergism at low concentration and antagonism at high concentration; the change from synergism to antagonism at higher IC50 level (CA, IA) Genotoxicity (alkaline comet test): no dose-effect relationship of the single toxins; mathematical modelling was not applicable for the mixture | [46] |
| Mycotoxin Combination | In Vitro Model | Mathematical Model Applied for the Endpoint | Endpoint Combined Effect | Reference |
|---|---|---|---|---|
| AOH (50 nM-10 µM) + ZEN (10 pM-1nM) AOH (50 nM-10 µM) + α-ZEL (1 pM-1nM) | Human endometrial adenocarcinoma cell line, Ishikawa | Chou and Chou-Talalay method | Estrogenic effect (AIP test) according to C: 61% synergism, 10% additive effect, 32% antagonism for AOH + ZEN 86% synergism, 14% antagonism for AOH + α-ZEL Estrogenic effect (AIP assay) according to CT: ZEN or α-ZEL:AOH (1:250) dominant synergism Cytotoxicity (SRB assay): not possible to calculate CI | [51] |
| DON + T2 | Human chondrocytic C28/I2, human hepatic epithelial L-02 and human tubular epithelial HK-2 cells | CI-Isobologram according to Chou-Talalay method; CI at IC10–90 (1:1) | Cytotoxicity (MTT test): synergism at IC10 in HK2 antagonism in C28/12, L-02 (IC10–90) and in HK2 (IC25–90) | [58] |
| DON + 15-ADON (1:1) DON + FX (3:1) DON + NIV (3:1) 15-ADON + FX (3:1) 15-ADON + NIV (3:1) FX + NIV (1:1) | Human gastric epithelial GES-1 cells | CI-Isobologram according to Chou-Talalay method; CI at IC10–IC90 | Cytotoxicity (OD test): synergism of DON + 15-ADON, DON + NIV, FX + NIV at IC10–IC70; DON +FX at IC10 and IC30; 15-ADON + FX at IC10 additive effect of FX + NIV at IC90 antagonism of 15A-DON + NIV at IC10–IC90; 15-ADON + FX at IC30-IC90; DON + FX at IC50-IC90; DON +15-ADON, DON + NIV, FX + NIV at IC90 | [59] |
| AFB1 + DON AFB1 + ZEN DON + ZEN AFB1 + DON + ZEN | HepG2 and (murine leukemia virus-induced tumor RAW 264.7 cells | CI-Isobologram according to Chou-Talalay method; CI at IC25,50,70 (1:1 and 1:1:1) | Cytotoxicity (Resazurin test) in HepG2: synergism of DON + ZEN AFB1 + DON + ZEN at IC25–70 additive effects of AFB1 + DON at IC25–70 antagonism of AFB1 + ZEN at IC25–70 Cytotoxicity in RAW 264.7: synergism of AFB1 + DON at IC25; DON + ZEN, AFB1 + DON + ZEN at IC50,70 additive effects of AFB1 + DON at IC50,70, DON + ZEN, AFB1 + DON + ZEN at IC25 antagonism of AFB1 + ZEN at IC25–70 | [60] |
| AFM1 + OTA AFM1 + α-ZEL AFM1 + ZEN OTA + ZEN OTA + α-ZEL ZEN + α-ZEL AFM1 + OTA + α-ZEL AFM1 + ZEN + α-ZEL AFM1 + OTA + ZEN OTA + ZEN + α-ZEL AFM1 + OTA + α-ZEL + ZEN | Caco-2 cells | CI-Isobologram according to Chou-Talalay method; CI at IC25,50,75,90 (1:1, 1:1:1 and 1:1:1.1) | Cytotoxicity (MTT test): synergism of AFM1 + OTA at IC50; OTA + ZEN at IC25,50; OTA + α-ZEL at IC25; ZEN + α-ZEL at IC75,90; AFM1 + ZEN + α-ZEL, AFM1 + OTA + ZEN and OTA + ZEN + α-ZEL at IC25; four toxins combination at IC25,50 additive effects of AFM1 + OTA at IC25,75; AFM1 + ZEN at IC25; OTA + ZEN and ZEN + α-ZEL at IC50; AFM1 + OTA + α-ZEL at IC25,50; AFM1 + OTA + ZEN and OTA + ZEN + α-ZEL at IC50 antagonism at AFM1 + OTA at IC90; AFM1 + α-ZEL at IC25–90; AFM1 + ZEN at IC50-90; OTA + ZEN at IC75,90; OTA + α-ZEL at IC25; ZEN + α-ZEL at IC25; AFM1 + OTA + α-ZEL, AFM1 + OTA + ZEN, OTA + ZEN + α-ZEL and AFM1 + OTA + α-ZEL + ZEN at IC75,90; AFM1 + ZEN + α-ZEL at IC50-90 | [61] |
| ZEN + α-ZEL ZEN + ß-ZEL α-ZEL + ß-ZEL | HepG2 cells | CI-Isobologram according to Chou-Talalay method; CI at IL25-IL75 (1:1) | Cytotoxicity (NR test): synergistic effect in all combinations, except additive effect for ZEA + β ZEL at IL25 Expression of pro-inflammatory cytokines (IL-1ß, TNF-α, IL-8): synergism of all mixtures for IL-8 at IL50.75; ZEN + α-ZEL (IL50,75) and, ZEN + β-ZEL (IL75) for IL-1β and TNF-α antagonism of all mixtures for all cytokines at IL25 except for ZEN + α-ZEL (synergism); ZEN + β-ZEL (IL50) for ILβ; α-ZEL + β-ZEL for IL-1β and TNF-α at IL50,75 | [62] |
| 3-ADON + AOH 15-ADON + AOH 3-ADON + 15-ADON AOH + 3-ADON + 15-ADON | HepG2 cells | CI-Isobologram according to Chou-Talalay method; CI at IC25,50,75,90 (1:1) | Cytotoxicity (MTT test) upon 24, 48 and 72 h: dominant synergism, 3-ADON + AOH (24 and 48 h and IC25 72 h), 15-ADON + AOH (24 h), 3-ADON + 15-ADON and AOH + 3-ADON + 15-ADON (all treatments) additive effect of 3-ADON + AOH IC50-90 (72 h); 15-ADON + AOH at IC25,50 (48 h) and IC50-90 (72 h) antagonism of 15-ADON + AOH at IC75,90 (48 h) and IC25 (72 h) | [63] |
| AFB1 + DON AFB1 + OTA DON+OTA | Caco-2 and HepG2 cells | CI-Isobologram according to Chou-Talalay method; CI at IC10–IC90 (1:1) | Cytotoxicity (MTT test) in Caco-2 cells: synergism of DON+OTA at IC10–IC90; AFB1 + DON at IC60-90; AFB1 + OTA at IC75-90 antagonism of AFB1 + OTA at IC10-50; AFB1 + DON at IC10,30 Cytotoxicity in HepG2 cells: synergism of AFB1 + DON at IC10-90; additive effects of AFB1 + OTA at IC10,90; DON + OTA at IC60,90 antagonism of DON + OTA at IC10-50 | [64] |
| DON + PAT DON + T2 PAT + T2 DON + T2 + PAT | HepG2 cells | CI-Isobologram according to Chou-Talalay method; CI at IC10–IC90 (1:1) | Cytotoxicity (MTT test) upon 24, 48 and 72 h: no synergism dominant additive effect of DON + PAT; DON + T2 upon 72 h and at IC75,90 (24 h); PAT + T2 upon 72 h and at IC10, 50-90 (24 h); DON + T2 + PAT upon 72 h and at IC50-90 (24 h) and IC25–90 (48 h) antagonism of DON + T2 upon 48 h and at IC10-50 (48 h); PAT + T-2 upon 48 h and at IC25 (24 h); DON + T2 + PAT at IC10,25 (24 h) and IC10 (48 h) | [65] |
| DON + NIV (1:0.6) NIV + FX (3:1) DON + FX (1:0.2) DON + NIV + FX (1:0.6:0.2) | Jurkat human T cells | CI-Isobologram according to Chou-Talalay method; CI at IC10, IC20 and IC30 | Cytotoxicity (MTT test): DON + NIV additive effect (IC10) and antagonism (IC20,30) NIV + FX synergism DON + FX antagonism DON + NIV + FX antagonism | [52] |
| DON + NIV | Differentiated three-dimensional porcine jejunal explants | CI-Isobologram according to Chou-Talalay method; CI at equimolar concentrations (1:1) | mRNA expression of cytokines: synergism in activation of all the tested pro-inflammatory genes (IL-1α,β, IL-8, IL-17A, IL-22) | [53] |
| DON + NIV DON + FX NIV + FX DON + NIV + FX | Human alveolar adenocarcinoma (A549) and bronchial 16HBE14o- cells primary human bronchial (hAECB) and nasal (hAECN) cells | CI-Isobologram method derived from the median-effect according to Chou at IC10,30,50 (1:1) | Cytotoxicity (MTT test): in A549 cells synergism of DON + NIV and DON + FX at IC10 and additive effect at IC30; antagonism of NIV + FX at IC30 and DON + NIV or FX at IC50 in 16HBE14o- cells synergism of DON + FX and NIV + FX at IC10-50; antagonism of DON + NIV at IC10-50 in hAECB cells synergism of binary mixtures at IC10.30 and NIV + FX at IC50; additive effects of DON + NIV and DON + FX at IC50 in hAECN cells of binary mixtures at IC30,50 and DON + NIV and NIV + FX at IC10; antagonism of DON + FX at IC10 | [54] |
| DON + ZEN (1:7.5) NIV + T-2 (1:0.067) (ratio of IC50) | HepaRG cells | CI-Isobologram according to Chou-Talalay method | Cytotoxicity (MTS): synergism of DON + ZEN at all applied concentrations - synergism of NIV + T-2 at low concentrations antagonism of NIV + T-2 at medium concentrations | [39] |
| AFB1 + DON (1:1.44) AFB1 + ZEN (1:15.19) DON + ZEN (1:10.56) AFB1 + DON + ZEN (1:1.44:15.19) | Fibroblast cell line BF-2 from the caudal fin of Lepomis macrochirus | CI-Isobologram according to Chou-Talalay method; CI at IC10-IC50 | Cytotoxicity (resazurin): synergism of AFB1 + DON and AFB1 + ZEN and ternary mixture at IC10-30 additive effect of ternary mixture at IC40 antagonism of DON + ZEN and ternary mixture at IC50 | [55] |
| BEA + STC (1:5) BEA + PAT (3.2:1) PAT + STC + (1:5) BEA + PAT + STC (3.2:1:5) | Chinese hamster ovary CHO-K1 cells | CI-Isobologram according to Chou-Talalay method; CI at IC5-IC50 | Cytotoxicity (MTT test) upon 24, 48 and 72 h: synergism of BEA + STC at IC5,10 (24 h); BEA + PAT at IC5 (24 h); PAT + STC at IC5,10 (24-72 h); BEA + PAT + STC at IC5,10 (24 h) and IC10-50 (72 h) additive effect of BEA + STC at IC25,50 (24 h), IC50 (48 h) and IC10-50 (72 h); BEA + PAT at IC10-50 (24 h) and IC25,50 (72 h); PAT + STC at IC25 (24-72 h) and IC50 (24, 48 h); BEA + PAT + STC at IC5 (72 h) and IC25,50 (24, 48h) antagonism of BEA + STC at IC5-25 (48 h); BEA +PAT at IC5-50 (48 h) and IC5,10 (72 h); PAT + STC at IC50 (72 h); BEA + PAT + STC at IC5,10 (48 h) | [56] |
| BEA + OTA | HepG2 cells | CI-Isobologram according to Chou-Talalay method; CI at IC25-IC90 (1:1) and equimolar ration (1:10) | Cytotoxicity (MTT test) upon 24, 48 and 72 h: synergism upon 72 h at IC25-IC90; 48 h at IC25-IC75; and 1:10 upon 48 and 72 h additive effects upon 24 h at IC25-IC90; 48 h at IC90; and 1:10 upon 24 h | [57] |
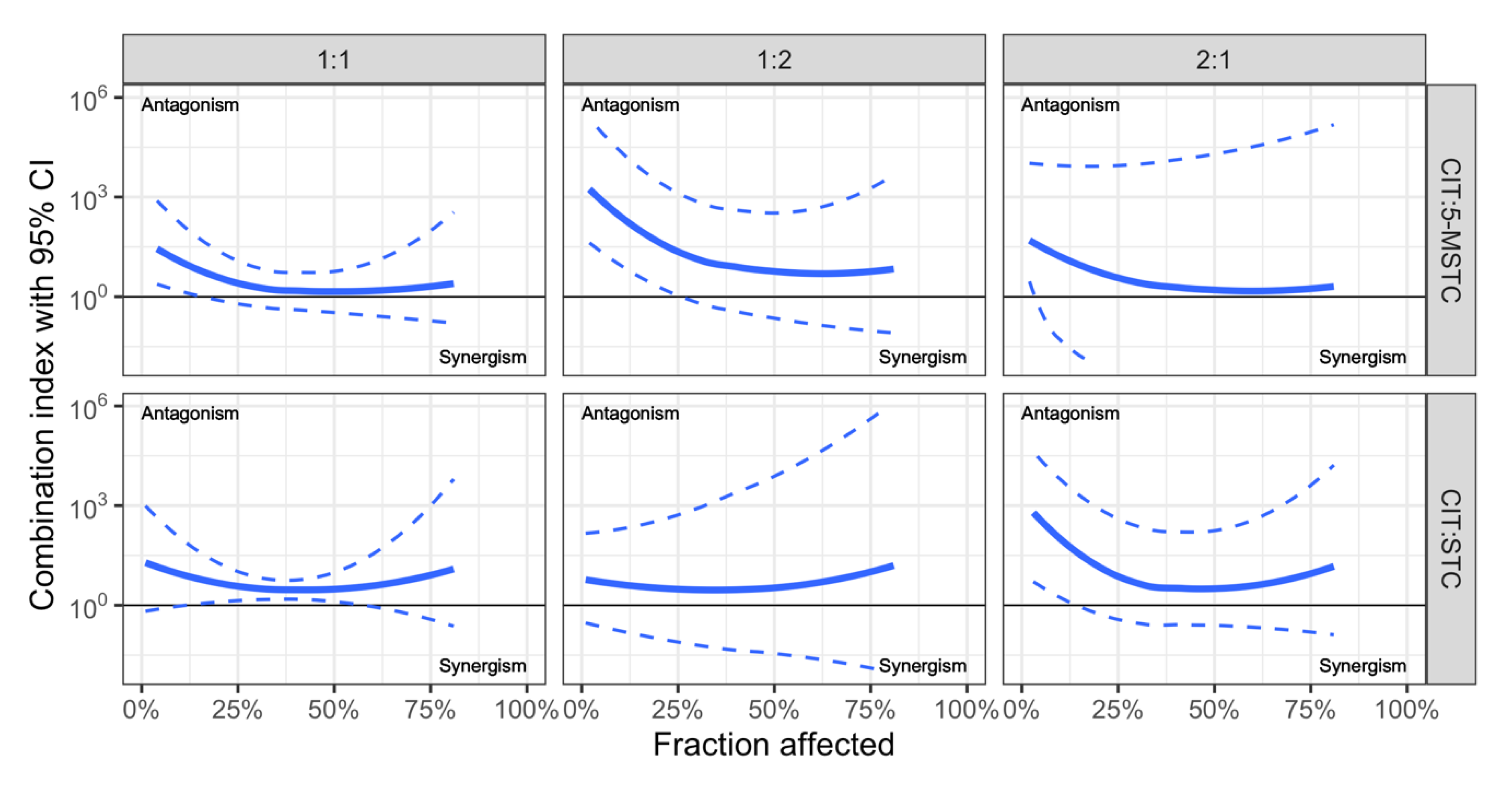
| CIT + STC CIT + M-STC | Human adenocarcinoma lung A549 cells | CI-Isobologram with correction for unparalleled dose-response curves, developed by Anastasiadi et al. [50]; “ray” desing with 1:1, 1:2 and 2:1 concentration ratios | Cytotoxicity (MTT test) additive effect antagonism exceptionally in low affected areas for CIT + 5-MSTC and 2:1 CIT + STC, also between IC25 and IC50 for CIT + STC | Personal unpublished data shown in Figure 2. |
| DON + T2 | Human C-28/I2 and newborn rat primary costal chondrocytes (RC) | MixLow method; combination ratios of DON and T-2 toxin (R1=1:1 R10= 10:1, R100=100:1 and R1000=1000:1). | Cytotoxicity (MTT test): synergism at fraction affected 0.5, 0.75, 0.9 of R10 concentrations in RC antagonism at fraction affected 0.25 of R100 in both cell types; fraction affected 0.5 of R100 in C-28/12; fraction 0.5 of R1000 in RC | [67] |
| Mycotoxin Combination | In Vitro Model | Statistical Analysis Applied for the Endpoint | Endpoint Combined Effect | Reference |
|---|---|---|---|---|
| DON + ZEN | Porcine splenic lymphocytes | ANOVA followed by the Tukey post hoc test (p < 0.05) | Antioxidant parameters (MDA, GSH, CAT, SOD, Gpx): synergism Apoptotic rate: synergism Expression of p53, Bcl-2, Bax, caspase-3, and caspase-8: synergism | [70] |
| DON + ZEN (at concentrations corresponding to the AED, TDI and ML) | HepaRG cells | Student’s t-test (p < 0.05) | Cytotoxicity (MTS test) upon 14, 28 and 42 days: at ML no antagonistic or synergistic effect Gene expression of CYP4F3B, CYP3A4, C/EBPα, HNF4α, aldolase B, transferrin, albumin and claudin-1 (qPCR): at AED majority of genes were ↑↑ after 14 days and ↓↓ after 28 days at TDI the gene expression upon 14 and 28 days were less different but more ↑↑ after 28 days at ML DON and DON+ZEA reduced the cell viability by more than 90%, no sufficient amounts of RNA DON + ZEN affected different genes than single DON and ZEA | [75] |
| DON + 3ADON (3:1) DON + 15-ADON (3:1) 3-ADON + 15-ADON (1:1) DON + 3-ADON + 15-ADON (3:1:1) (ratios of IC50) | HepG2 cells | ANOVA followed by the Tukey post hoc test (p ≤ 0.05) | Oxidative stress (ROS and MDA): binary mixtures significantly increased ROS vs. control and initial time binary and tertiary mixtures increased MDA vs. control (24, 48 and 72 h) Cell cycle distribution upon 48 h (flow cytometry): DON + 3-ADON ↓ G0/G1 and S, G0/G1 and S, G2/M phase ↑ at lower and ↓ at higher concentrations in respect to control DON + 15-ADON ↑ G0/G1 and G2/M at lower and ↓ at higher concentrations in respect to control 3-ADON + 15-ADON ↓ G0/G1 and S at all concentrations vs. control tertiary combination ↓ G0/G1, S and G2/M vs. control Micronuclei (MN): binary mixtures ↑ in MN at lower concentrations vs. control tertiary mixtures ↑ in MN at all concentrations vs. control | [73,74] |
| ENN A + A1 + B + B1 (1.5 or 3 µM) ENN A + A1 + B + B1 + DON (1.5 or 3 µM) BEA (2.5 µM) + DON (1.5 or 3 µM) Apicidin (0.438 µM) + DON (1.5 or 3 µM) AURO (5 µM) + DON (1.5 or 3 µM) | Porcine epithelial cells IPEC-J2 | ANOVA followed by the Dunnett’s t-test or Kruskall-Wallis test (p < 0.05) | TEER upon 24, 48 and 72 h: dominant additive effect DON had no effect on enniatin-induced TEER decrease BEA + DON did not significantly reduce TEER | [76] |
| OTA + CIT | Multiple organ co-culture (IdMOC) of HepG2 and 3T3 cells | Paired sample t-test (p < 0.05) | Luciferin-IPA metabolism assay: synergism at 20% IC50 (CTN forms a reactive metabolite that diffuses out of HepG2 to cause cytotoxicity to 3T3 cells synergistically with OTA) | [71] |
| OTA + CIT (equimolar concentrations 0–30 µM) | Human embryonic kidney HEK293 cells | No statistical analysis indicated/ effect of combination was compared to the effects of mycotoxins acting alone | Cytotoxicity (MTT test): synergism based on IC50 of single OTA (16 µM) and CIT (189 µM) vs. combination (7 µM) | [72] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kifer, D.; Jakšić, D.; Šegvić Klarić, M. Assessing the Effect of Mycotoxin Combinations: Which Mathematical Model Is (the Most) Appropriate? Toxins 2020, 12, 153. https://doi.org/10.3390/toxins12030153
Kifer D, Jakšić D, Šegvić Klarić M. Assessing the Effect of Mycotoxin Combinations: Which Mathematical Model Is (the Most) Appropriate? Toxins. 2020; 12(3):153. https://doi.org/10.3390/toxins12030153
Chicago/Turabian StyleKifer, Domagoj, Daniela Jakšić, and Maja Šegvić Klarić. 2020. "Assessing the Effect of Mycotoxin Combinations: Which Mathematical Model Is (the Most) Appropriate?" Toxins 12, no. 3: 153. https://doi.org/10.3390/toxins12030153
APA StyleKifer, D., Jakšić, D., & Šegvić Klarić, M. (2020). Assessing the Effect of Mycotoxin Combinations: Which Mathematical Model Is (the Most) Appropriate? Toxins, 12(3), 153. https://doi.org/10.3390/toxins12030153