In Vitro Immunological Cross-Reactivity of Thai Polyvalent and Monovalent Antivenoms with Asian Viper Venoms



 , and
, and
Abstract
1. Introduction
2. Results
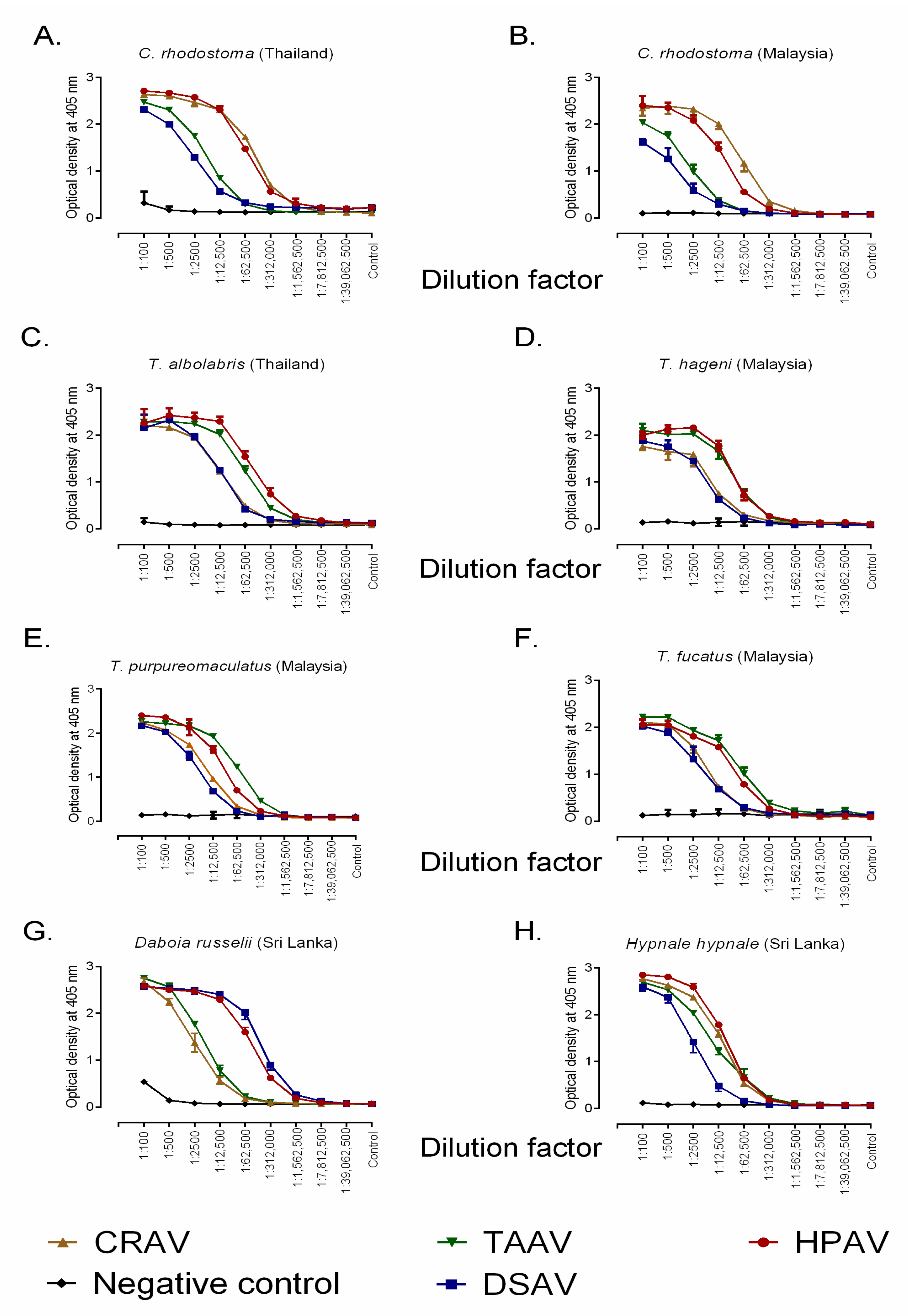
2.1. Immunological Cross-Reactivity Determined by End-Point Titration (EPT) ELISA
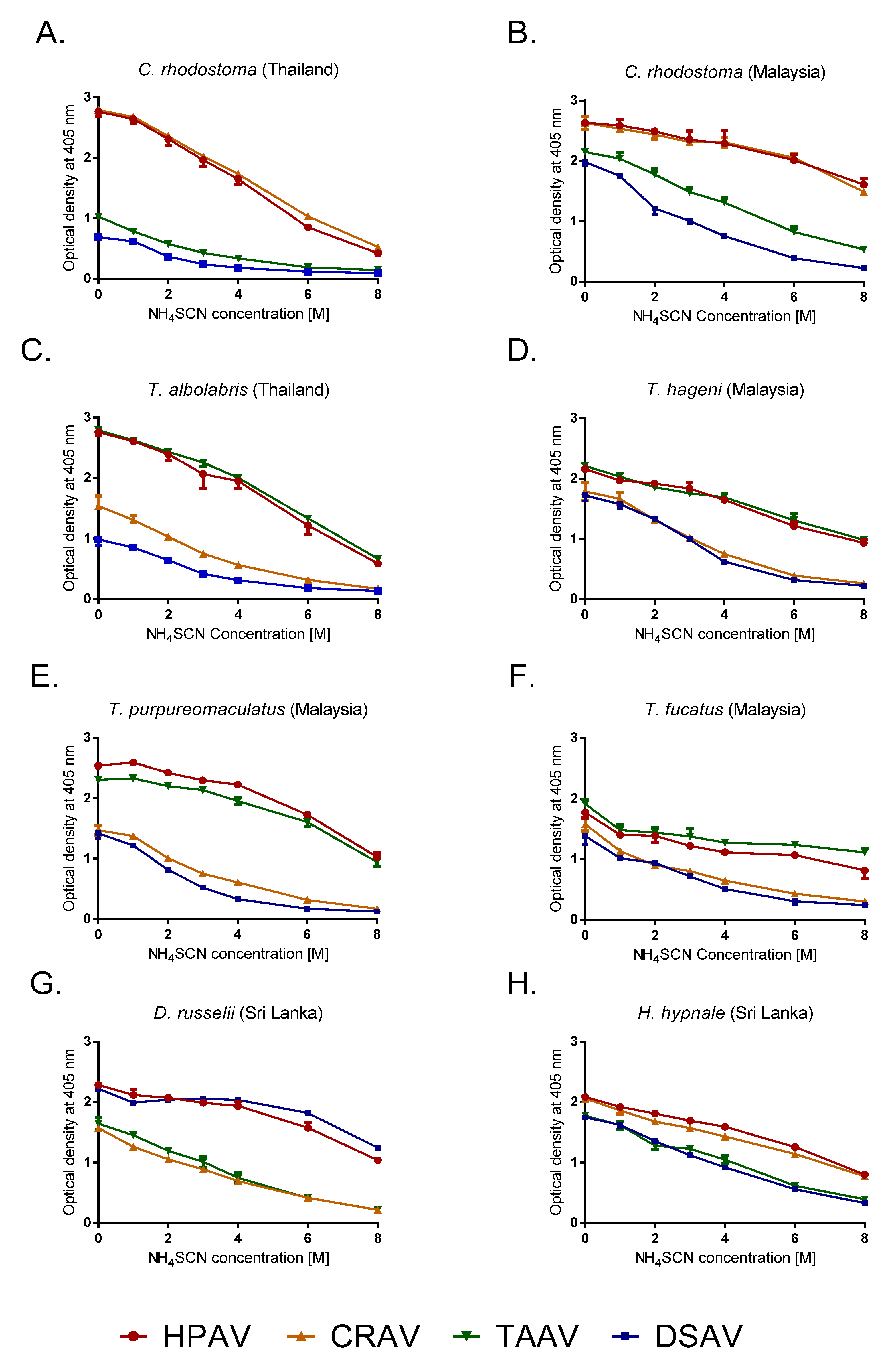
2.2. Immunological Avidity Determined by Chaotropic ELISA
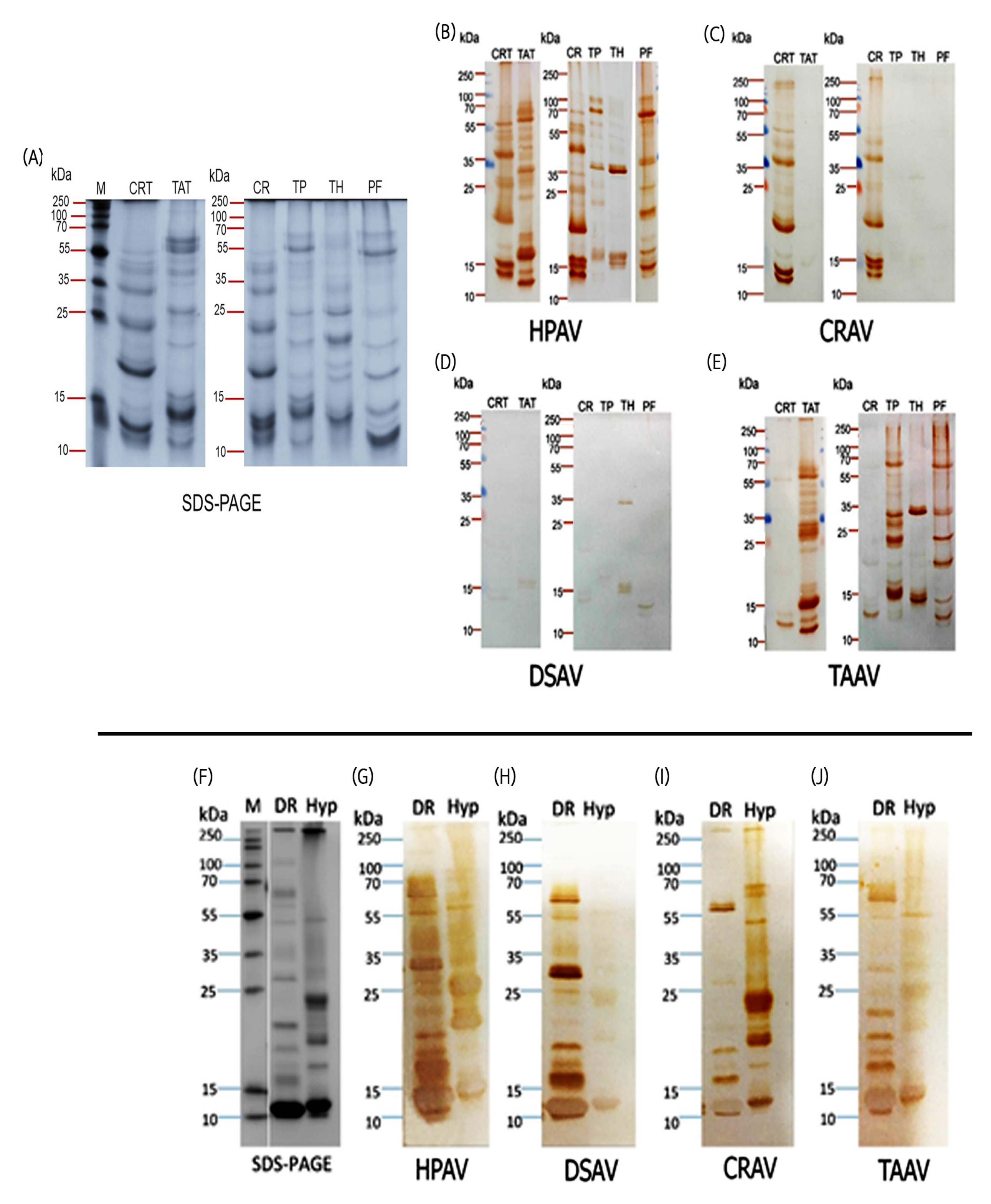
2.3. Immunological Cross-Reactivity Visualized by Immunoblotting: South-East Asian Pit Viper Venoms
2.4. Immunological Cross-Reactivity Visualized by Immunoblotting: Sri Lankan Viper Venoms
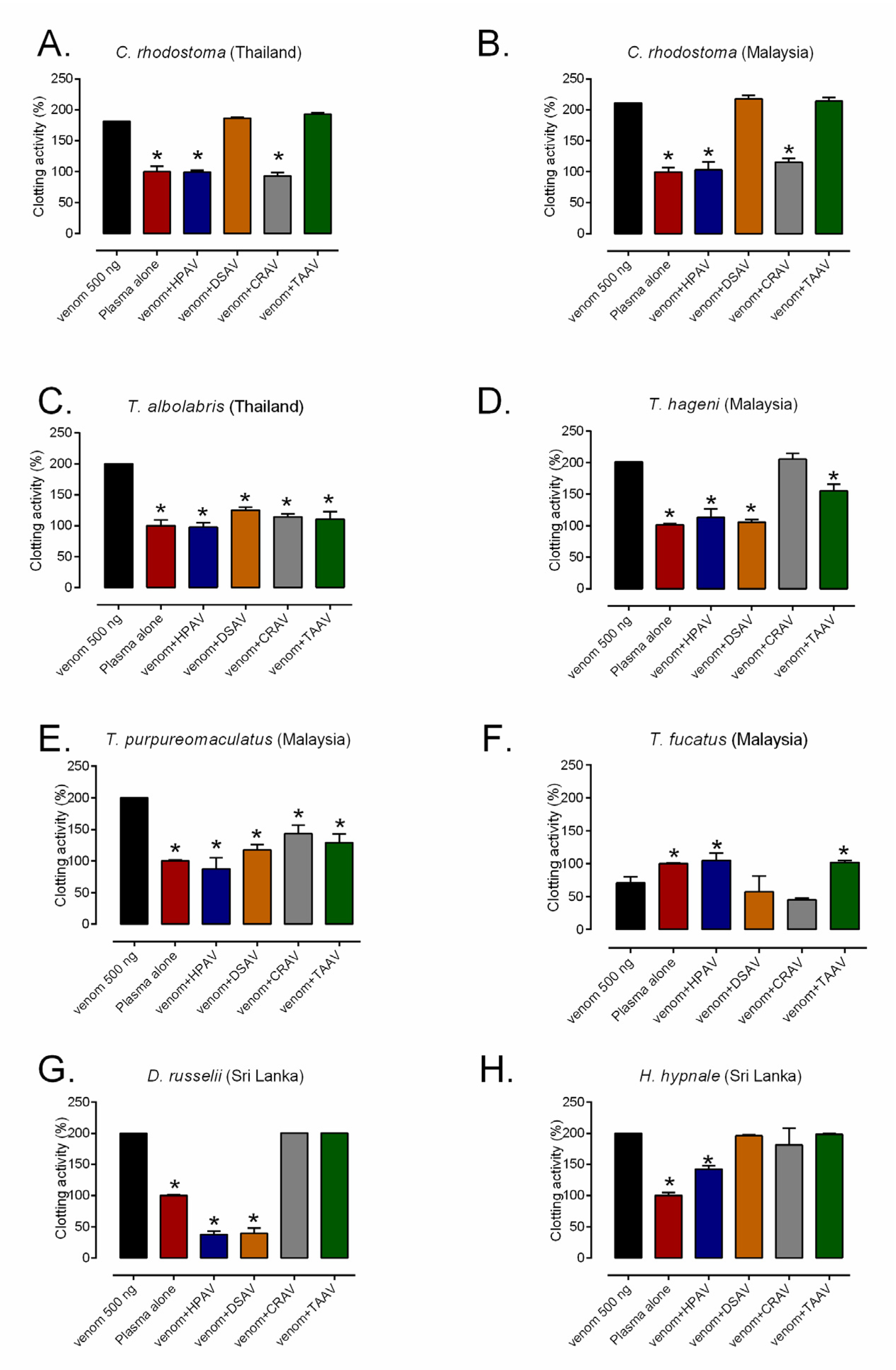
2.5. Coagulopathic Venom Activity and Antivenom Neutralization Measured by Plasma Clotting Assay
3. Discussion
4. Materials and Methods
4.1. Snake Venom and Antivenoms
4.2. Immunological Assays
4.2.1. End-Point Titration (EPT) ELISA
4.2.2. Relative Avidity ELISA
4.2.3. SDS-PAGE
4.2.4. Western Blotting
4.3. Plasma Coagulation Assay
4.4. Analysis of Results and Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gutierrez, J.M.; Williams, D.; Fan, H.W.; Warrell, D.A. Snakebite envenoming from a global perspective: Towards an integrated approach. Toxicon 2010, 56, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Primers 2017, 3, 17079. [Google Scholar] [CrossRef] [PubMed]
- WHO. Venomous snakes of the South-East Asia Region, their venoms and pathophysiology of human envenoming. In Guidelines for the Management of Snakebites, 2nd ed.; Regional Office for Southeast Asia: New Delhi, India, 2016; pp. 27–71. [Google Scholar]
- Chaisakul, J.; Alsolaiss, J.; Charoenpitakchai, M.; Wiwatwarayos, K.; Sookprasert, N.; Harrison, R.A.; Chaiyabutr, N.; Chanhome, L.; Tan, C.H.; Casewell, N.R. Evaluation of the geographical utility of Eastern Russell’s viper (Daboia siamensis) antivenom from Thailand and an assessment of its protective effects against venom-induced nephrotoxicity. PLoS Negl. Trop. Dis. 2019, 13, e0007338. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, M.A.; Isbister, G.K. Commercial monovalent antivenoms in Australia are polyvalent. Toxicon. 2009, 54, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, I.S.; Abubakar, S.B.; Habib, A.G.; Nasidi, A.; Durfa, N.; Yusuf, P.O.; Larnyang, S.; Garnvwa, J.; Sokomba, E.; Salako, L.; et al. Randomised controlled double-blind non-inferiority trial of two antivenoms for saw-scaled or carpet viper (Echis ocellatus) envenoming in Nigeria. PLoS Negl. Trop. Dis. 2010, 4, e767. [Google Scholar] [CrossRef]
- Silva, A.; Hodgson, W.C.; Isbister, G.K. Cross-Neutralisation of in vitro neurotoxicity of Asian and Australian snake neurotoxins and venoms by different antivenoms. Toxins 2016, 8, 302. [Google Scholar] [CrossRef]
- Tan, C.H.; Liew, J.L.; Tan, N.H.; Ismail, A.K.; Maharani, T.; Khomvilai, S.; Sitprija, V. Cross reactivity and lethality neutralization of venoms of Indonesian Trimeresurus complex species by Thai Green Pit Viper Antivenom. Toxicon 2017, 140, 32–37. [Google Scholar] [CrossRef]
- Casewell, N.R.; Cook, D.A.; Wagstaff, S.C.; Nasidi, A.; Durfa, N.; Wuster, W.; Harrison, R.A. Pre-clinical assays predict pan-African Echis viper efficacy for a species-specific antivenom. PLoS Negl. Trop. Dis. 2010, 4, e851. [Google Scholar] [CrossRef]
- Casewell, N.R.; Al-Abdulla, I.; Smith, D.; Coxon, R.; Landon, J. Immunological cross-reactivity and neutralisation of European viper venoms with the monospecific Vipera berus antivenom ViperaTAb. Toxins 2014, 6, 2471–2482. [Google Scholar] [CrossRef]
- Senji Laxme, R.R.; Khochare, S.; de Souza, H.F.; Ahuja, B.; Suranse, V.; Martin, G.; Whitaker, R.; Sunagar, K. Beyond the ‘big four’: Venom profiling of the medically important yet neglected Indian snakes reveals disturbing antivenom deficiencies. PLoS Negl. Trop. Dis. 2019, 13, e0007899. [Google Scholar] [CrossRef] [PubMed]
- Visser, L.E.; Kyei-Faried, S.; Belcher, D.W.; Geelhoed, D.W.; van Leeuwen, J.S.; van Roosmalen, J. Failure of a new antivenom to treat Echis ocellatus snake bite in rural Ghana: The importance of quality surveillance. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Jackson, T.N.W.; Laustsen, A.H.; Sunagar, K. Causes and consequences of snake venom variation. Trends Pharmacol. Sci. 2020, 41, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Pla, D.; Sanz, L.; Quesada-Bernat, S.; Villalta, M.; Baal, J.; Chowdhury, M.A.W.; Leon, G.; Gutierrez, J.M.; Kuch, U.; Calvete, J.J. Phylovenomics of Daboia russelii across the Indian subcontinent. Bioactivities and comparative in vivo neutralization and in vitro third-generation antivenomics of antivenoms against venoms from India, Bangladesh and Sri Lanka. J. Proteom. 2019, 207, 103443. [Google Scholar] [CrossRef]
- Tasoulis, T.; Isbister, G.K. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef]
- Kornhauser, R.; Isbister, G.K.; O’Leary, M.A.; Mirtschin, P.; Dunstan, N.; Hodgson, W.C. Cross-neutralisation of the neurotoxic effects of Egyptian cobra venom with commercial tiger snake antivenom. Basic Clin. Pharmacol. Toxicol. 2013, 112, 138–143. [Google Scholar] [CrossRef]
- Ainsworth, S.; Slagboom, J.; Alomran, N.; Pla, D.; Alhamdi, Y.; King, S.I.; Bolton, F.M.S.; Gutierrez, J.M.; Vonk, F.J.; Toh, C.H.; et al. The paraspecific neutralisation of snake venom induced coagulopathy by antivenoms. Commun. Biol. 2018, 1, 34. [Google Scholar] [CrossRef]
- Leong, P.K.; Tan, C.H.; Sim, S.M.; Fung, S.Y.; Sumana, K.; Sitprija, V.; Tan, N.H. Cross neutralization of common Southeast Asian viperid venoms by a Thai polyvalent snake antivenom (Hemato Polyvalent Snake Antivenom). Acta Trop. 2014, 132, 7–14. [Google Scholar] [CrossRef]
- Debono, J.; Bos, M.H.A.; Frank, N.; Fry, B. Clinical implications of differential antivenom efficacy in neutralising coagulotoxicity produced by venoms from species within the arboreal viperid snake genus Trimeresurus. Toxicol. Lett. 2019, 316, 35–48. [Google Scholar] [CrossRef]
- Harrison, R.A.; Oluoch, G.O.; Ainsworth, S.; Alsolaiss, J.; Bolton, F.; Arias, A.S.; Gutierrez, J.M.; Rowley, P.; Kalya, S.; Ozwara, H.; et al. Preclinical antivenom-efficacy testing reveals potentially disturbing deficiencies of snakebite treatment capability in East Africa. PLoS Negl. Trop. Dis. 2017, 11, e0005969. [Google Scholar] [CrossRef]
- Yang, J.Y.; Hui, H.; Lee, A.C. Severe coagulopathy associated with white-lipped green pit viper bite. Hong Kong Med. J. 2007, 13, 392–395. [Google Scholar] [PubMed]
- Morais, V. Antivenom therapy: Efficacy of premedication for the prevention of adverse reactions. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 7. [Google Scholar] [CrossRef] [PubMed]
- Jamaiah, I.; Rohela, M.; Ng, T.K.; Ch’ng, K.B.; Teh, Y.S.; Nurulhuda, A.L.; Suhaili, N. Retrospective prevalence of snakebites from Hospital Kuala Lumpur (HKL) (1999–2003). Southeast. Asian J. Trop. Med. Public Health 2006, 37, 200–205. [Google Scholar] [PubMed]
- Jamaiah, I.; Rohela, M.; Roshalina, R.; Undan, R.C. Prevalence of snake bites in Kangar District Hospital, Perlis, west Malaysia: A retrospective study (January 1999–December 2000). Southeast. Asian J. Trop. Med. Public Health 2004, 35, 962–965. [Google Scholar] [PubMed]
- McGain, F.; Limbo, A.; Williams, D.J.; Didei, G.; Winkel, K.D. Snakebite mortality at Port Moresby General Hospital, Papua New Guinea, 1992–2001. Med. J. Aust. 2004, 181, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Chean, C.C. Borneo PET: Rural Challenge. In Proceedings of the ASEAN Marine Animal & Snake Envenomation (Poisoning) Management, 4th International Symposium, Penang, Malaysia, 27 November–1 December 2016. [Google Scholar]
- Atieka, S.H.A.; Maharani, T.; Dradjat, R.S.; Setijowati, N. Risk Factors evaluation of snakebite outcome andcomplication: Study from Bondowoso General Hospital, East Java, Indonesia. In Proceedings of the ASEAN Marine Animal & Sanke Envenomation (Poisioning) Management, 4th International Symposium, Penang, Malaysia, 27 November–1 December 2016. [Google Scholar]
- Das, I.A.N.; Liat, L.B. Venomous terestrial snakes of Malaysia: Their Indentity and Biology. In Clinical Toxinology in Asia Pacific and Africa; Gopalakrishnakone, P., Ravindra Fernando, A.F., Gnanathasan, C.A., Habib, A.G., Yang, C.-C., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 53–69. [Google Scholar]
- Ismail, A.K. Snakebite and Envenomation Management in Malaysia. In Clinical Toxinology in Asia Pacific and Africa, Toxinology; Gopalakrishnakone, P., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 72–102. [Google Scholar]
- Liew, J.L.; Tan, N.H.; Tan, C.H. Proteomics and preclinical antivenom neutralization of the mangrove pit viper (Trimeresurus purpureomaculatus, Malaysia) and white-lipped pit viper (Trimeresurus albolabris, Thailand) venoms. Acta Trop. 2020. [Google Scholar] [CrossRef] [PubMed]
- Mong, R.; Tan, H.H. Snakebite by the Shore pit viper (Trimeresurus purpureomaculatus) treated with polyvalent antivenom. Wilderness Environ. Med. 2016, 27, 266–270. [Google Scholar] [CrossRef]
- Slagboom, J.; Kool, J.; Harrison, R.A.; Casewell, N.R. Haemotoxic snake venoms: Their functional activity, impact on snakebite victims and pharmaceutical promise. Br. J. Haematol. 2017, 177, 947–959. [Google Scholar] [CrossRef]
- White, J. Snake venoms and coagulopathy. Toxicon Off. J. Int. Soc. Toxinol. 2005, 45, 951–967. [Google Scholar] [CrossRef]
- Villalta, M.; Sanchez, A.; Herrera, M.; Vargas, M.; Segura, A.; Cerdas, M.; Estrada, R.; Gawarammana, I.; Keyler, D.E.; McWhorter, K.; et al. Development of a new polyspecific antivenom for snakebite envenoming in Sri Lanka: Analysis of its preclinical efficacy as compared to a currently available antivenom. Toxicon Off. J. Int. Soc. Toxinol. 2016, 122, 152–159. [Google Scholar] [CrossRef]
- Tan, C.H.; Leong, P.K.; Fung, S.Y.; Sim, S.M.; Ponnudurai, G.; Ariaratnam, C.; Khomvilai, S.; Sitprija, V.; Tan, N.H. Cross neutralization of Hypnale hypnale (hump-nosed pit viper) venom by polyvalent and monovalent Malayan pit viper antivenoms in vitro and in a rodent model. Acta Trop. 2011, 117, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Still, K.B.M.; Nandlal, R.S.S.; Slagboom, J.; Somsen, G.W.; Casewell, N.R.; Kool, J. Multipurpose HTS coagulation analysis: Assay development and assessment of coagulopathic snake venoms. Toxins 2017, 9, 382. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Venoms | Antivenom | |||
|---|---|---|---|---|
| Hematopolyvalent (HPAV) | D. siamensis (DSAV) | C. rhodostoma (CRAV) | T. albolabris (TAAV) | |
| C. rhodostoma (Thailand) ** | 2.57 ± 0.03 | 1.29 ± 0.02 | 2.46 ± 0.08 | 1.75 ± 0.05 |
| C. rhodostoma (Malaysia) | 2.07 ± 0.11 | 0.58 ± 0.14 | 2.32 ± 0.18 | 0.98 ± 0.15 |
| T. albolabris (Thailand) ** | 2.37 ± 0.10 | 1.95 ± 0.07 | 1.94 ± 0.02 | 2.24 ± 0.05 |
| T. hageni (Malaysia) | 2.16 ± 0.04 | 1.44 ± 0.06 | 1.65 ± 0.20 | 2.03 ± 0.06 |
| T. purpureomaculatus (Malaysia) | 2.13 ± 0.11 | 1.48 ± 0.08 | 1.73 ± 0.05 | 2.16 ± 0.07 |
| T. fucatus (Malaysia) | 1.81 ± 0.05 | 1.32 ± 0.26 | 1.57 ± 0.12 | 1.93 ± 0.01 |
| D. russelii (Sri Lanka) | 2.47 ± 0.04 | 2.50 ± 0.09 | 1.37 ± 0.19 | 1.77 ± 0.03 |
| H. hypnale (Sri Lanka) | 2.59 ± 0.08 | 1.42 ± 0.22 | 2.38 ± 0.02 | 2.03 ± 0.01 |
| Venoms | Antivenom | |||
|---|---|---|---|---|
| Hematopolyvalent (HPAV) | D. siamensis (DSAV) | C. rhodostoma (CRAV) | T. albolabris (TAAV) | |
| C. rhodostoma (Thailand) ** | 29.0 ± 4.0 | 75.0 ± 0.1 | 27.0 ± 1.1 | 65.1 ± 4.5 |
| C. rhodostoma (Malaysia) | 16.9 ± 4.0 | 61.9 ± 1.8 | 12.9 ± 5.5 | 39.0 ± 3.5 |
| T. albolabris (Thailand) ** | 13.6 ± 1.0 | 60.2 ± 2.0 | 33.3 ± 5.1 | 19.5 ± 4.6 |
| T. hageni (Malaysia) | 23.7 ± 0.4 | 63.6 ± 2.6 | 57.8 ± 1.4 | 23.3 ± 1.6 |
| T. purpureomaculatus (Malaysia) | 12.2 ± 1.7 | 76.7 ± 1.0 | 58.7 ± 4.0 | 15.2 ± 1.5 |
| T. fucatus (Malaysia) | 34.9 ± 6.3 | 63.2 ± 3.2 | 59.0 ± 1.9 | 31.8 ± 1.2 |
| D. russelii (Sri Lanka) | 15.2 ± 4.9 | 8.2 ± 1.3 | 55.8 ± 1.9 | 54.5 ± 3.3 |
| H. hypnale (Sri Lanka) | 23.7 ± 0.8 | 47.2 ± 3.3 | 30.4 ± 0.4 | 40.9 ± 4.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaisakul, J.; Rusmili, M.R.A.; Alsolaiss, J.; Albulescu, L.-O.; Harrison, R.A.; Othman, I.; Casewell, N.R. In Vitro Immunological Cross-Reactivity of Thai Polyvalent and Monovalent Antivenoms with Asian Viper Venoms. Toxins 2020, 12, 766. https://doi.org/10.3390/toxins12120766
Chaisakul J, Rusmili MRA, Alsolaiss J, Albulescu L-O, Harrison RA, Othman I, Casewell NR. In Vitro Immunological Cross-Reactivity of Thai Polyvalent and Monovalent Antivenoms with Asian Viper Venoms. Toxins. 2020; 12(12):766. https://doi.org/10.3390/toxins12120766
Chicago/Turabian StyleChaisakul, Janeyuth, Muhamad Rusdi Ahmad Rusmili, Jaffer Alsolaiss, Laura-Oana Albulescu, Robert A. Harrison, Iekhsan Othman, and Nicholas R. Casewell. 2020. "In Vitro Immunological Cross-Reactivity of Thai Polyvalent and Monovalent Antivenoms with Asian Viper Venoms" Toxins 12, no. 12: 766. https://doi.org/10.3390/toxins12120766
APA StyleChaisakul, J., Rusmili, M. R. A., Alsolaiss, J., Albulescu, L.-O., Harrison, R. A., Othman, I., & Casewell, N. R. (2020). In Vitro Immunological Cross-Reactivity of Thai Polyvalent and Monovalent Antivenoms with Asian Viper Venoms. Toxins, 12(12), 766. https://doi.org/10.3390/toxins12120766