Use of Integrative Interactomics for Improvement of Farm Animal Health and Welfare: An Example with Fescue Toxicosis

{kind=link}
{kind=link}
Abstract
1. Introduction
2. Technological Advances Increase Throughput for Toxicology Screening
3. Maintaining Animal Productivity and Wellness in Adverse Environments, Toxicity Included
4. Fescue Toxicosis
4.1. Tall Fescue and Epichloë Coenophiala
4.2. Ergot Alkaloids: Presence, Biosynthesis, and Monoaminergic Activities
4.3. Ergot Alkaloid Metabolism in Ruminant Animals
4.4. Biomarkers of Ergot Alkaloid Exposure and Effect
4.5. Adverse Effects of Toxic Tall Fescue Grazing and Ergot Alkaloids on Livestock
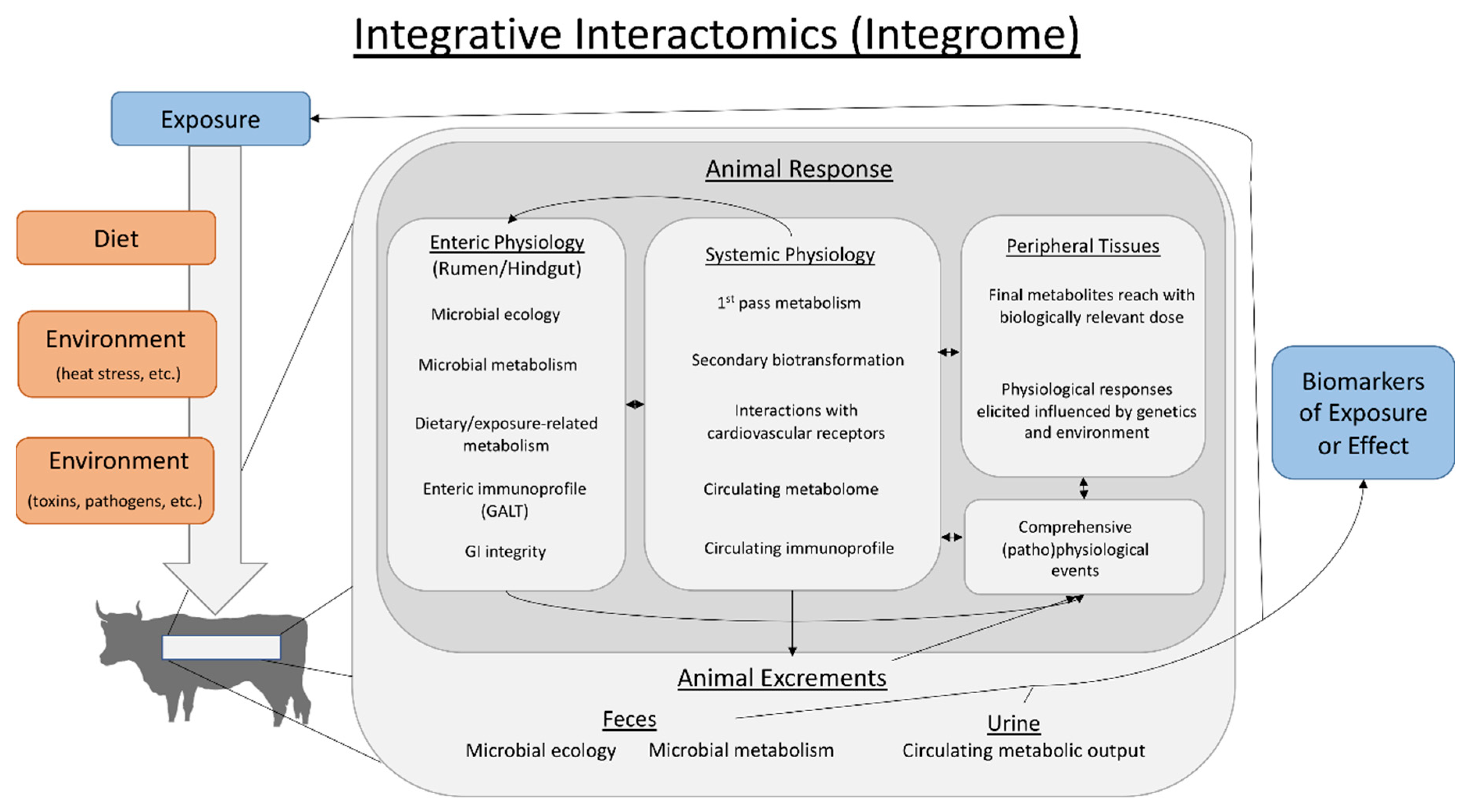
5. The Case for Integrative Interactomics in Fescue Toxicosis Studies
5.1. Effects of Epichloë Coenophiala Infection on the Plant
5.2. Effects of E. coenophiala-Infected Tall Fescue on the Animal
5.2.1. Toxic Tall Fescue Effects along the Bovine Alimentary Tract
5.2.2. Metabolic Effects of Toxic Tall Fescue Grazing
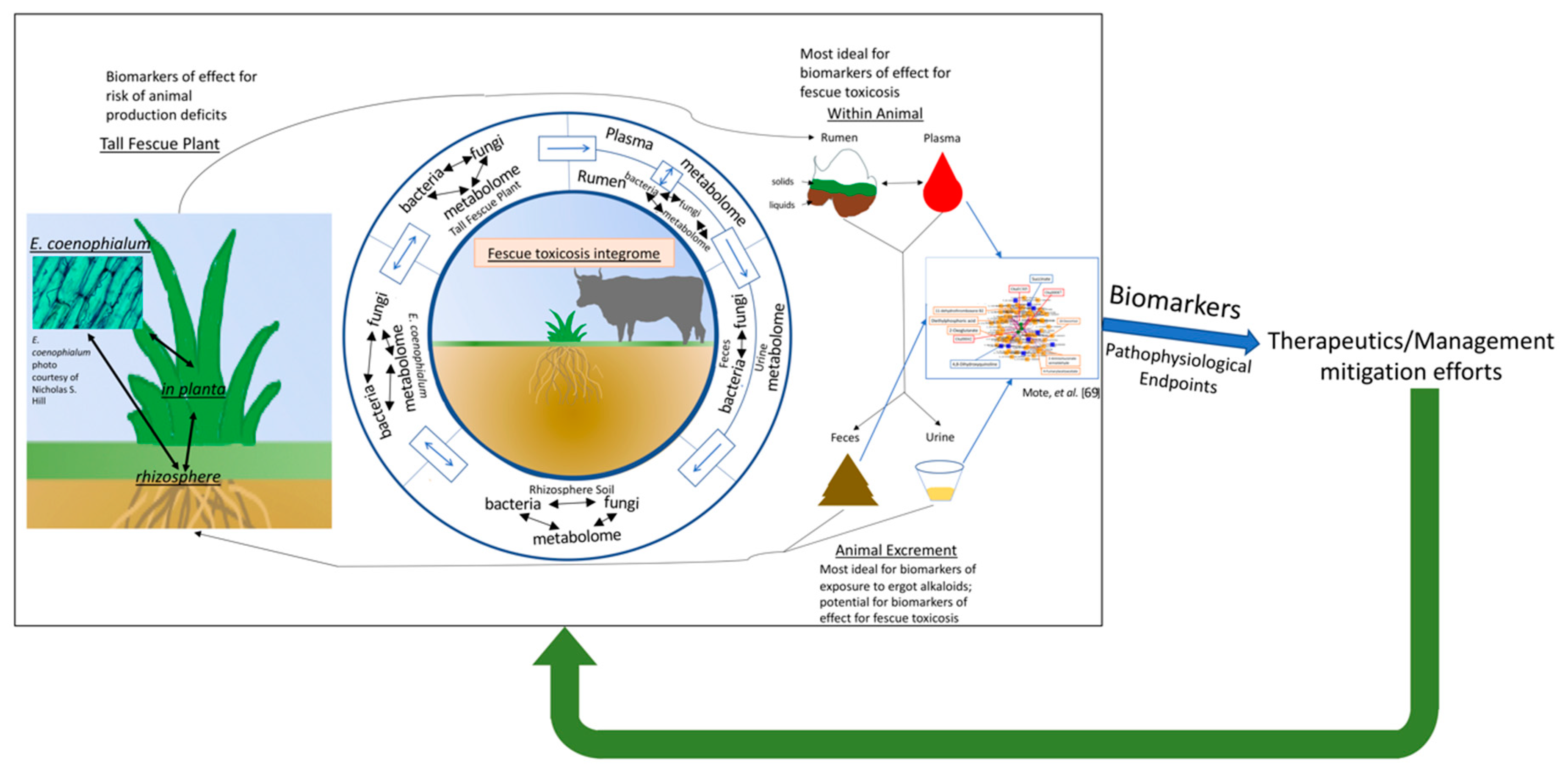
5.2.3. Understanding the Fescue Toxicosis Integrome
6. Where Is This Approach for Fescue Toxicosis Research Headed?
7. Applicability of Integrative Interactomics across Agriculture
8. Conclusions
Funding
Conflicts of Interest
References
- United Nations, DESA. World Population Prospects 2019; United Nations, Department of Economic and Social Affairs: New York, NY, USA, 2019. [Google Scholar]
- USDA OCE. USDA Agricultural Projections to 2028; USDA Office of the Chief Economist Outlook Board: Washington, DC, USA, 2019; p. 108.
- Li, J.Y.; You, Z.; Wang, Q.; Zhou, Z.J.; Qiu, Y.; Luo, R.; Ge, X.Y. The epidemic of 2019-novel-coronavirus (2019-nCoV) pneumonia and insights for emerging infectious diseases in the future. Microbes Infect. 2020, 22, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Capalbo, S.M.; Antle, J.M.; Seavert, C. Next generation data systems and knowledge products to support agricultural producers and science-based policy decision making. Agric. Syst. 2017, 155, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Unamba, C.I.; Nag, A.; Sharma, R.K. Next Generation Sequencing Technologies: The Doorway to the Unexplored Genomics of Non-Model Plants. Front. Plant Sci. 2015, 6, 1074. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2019, 10, 115–132. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Global Rumen Census, C.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef]
- Loor, J.J.; Elolimy, A.A.; McCann, J.C. Dietary impacts on rumen microbiota in beef and dairy production. Anim. Front. 2016, 6, 22–29. [Google Scholar] [CrossRef]
- Wolff, S.M.; Ellison, M.J.; Hao, Y.; Cockrum, R.R.; Austin, K.J.; Baraboo, M.; Burch, K.; Lee, H.J.; Maurer, T.; Patil, R.; et al. Diet shifts provoke complex and variable changes in the metabolic networks of the ruminal microbiome. Microbiome 2017, 5, 60. [Google Scholar] [CrossRef]
- Zehavi, T.; Probst, M.; Mizrahi, I. Insights Into Culturomics of the Rumen Microbiome. Front. Microbiol. 2018, 9, 1999. [Google Scholar] [CrossRef]
- Blum, B.C.; Mousavi, F.; Emili, A. Single-platform ‘multi-omic’ profiling: Unified mass spectrometry and computational workflows for integrative proteomics-metabolomics analysis. Mol. Omics 2018, 14, 307–319. [Google Scholar] [CrossRef]
- Griffiths, W.J. Metabolomics, Metabonomics, and Metabolic Profiling; Royal Society of Chemistry: London, UK, 2008. [Google Scholar]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2008, 5, 3–21. [Google Scholar] [CrossRef]
- Cevallos-Cevallos, J.M.; Reyes-De-Corcuera, J.I.; Etxeberria, E.; Danyluk, M.D.; Rodrick, G.E. Metabolomic analysis in food science: A review. Trends Food Sci. Technol. 2009, 20, 557–566. [Google Scholar] [CrossRef]
- Yu, T.; Jones, D.P. Improving peak detection in high-resolution LC/MS metabolomics data using preexisting knowledge and machine learning approach. Bioinformatics 2014, 30, 2941–2948. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.M.; Walker, D.I.; Liang, Y.; Uppal, K.; Soltow, Q.A.; Tran, V.; Strobel, F.; Quyyumi, A.A.; Ziegler, T.R.; Pennell, K.D.; et al. Reference Standardization for Mass Spectrometry and High-resolution Metabolomics Applications to Exposome Research. Toxicol. Sci. 2015, 148, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Park, Y.; Johnson, J.M.; Jones, D.P. apLCMS--adaptive processing of high-resolution LC/MS data. Bioinformatics 2009, 25, 1930–1936. [Google Scholar] [CrossRef] [PubMed]
- Uppal, K.; Soltow, Q.A.; Strobel, F.H.; Pittard, W.S.; Gernert, K.M.; Yu, T.; Jones, D.P. xMSanalyzer: Automated pipeline for improved feature detection and downstream analysis of large-scale, non-targeted metabolomics data. BMC Bioinform. 2013, 14, 15. [Google Scholar] [CrossRef]
- Mallon, T.M.; Krahl, P.K.; Haines, K.M., Jr.; Walker, D.I.; Thatcher, T.; Woeller, C.F.; Thakar, J.; Hopke, P.K.; Gaydos, J.C.; Smith, M.R.; et al. Use of Biomarkers to Assess Environmental Exposures and Health Outcomes in Deployed Troops. J. Occup. Environ. Med. 2019, 61 (Suppl. 12), S1–S4. [Google Scholar] [CrossRef]
- Niedzwiecki, M.M.; Walker, D.I.; Vermeulen, R.; Chadeau-Hyam, M.; Jones, D.P.; Miller, G.W. The Exposome: Molecules to Populations. Annu. Rev. Pharmacol. Toxicol. 2019, 59, 107–127. [Google Scholar] [CrossRef]
- Uppal, K.; Ma, C.; Go, Y.M.; Jones, D.P.; Wren, J. xMWAS: A data-driven integration and differential network analysis tool. Bioinformatics 2018, 34, 701–702. [Google Scholar] [CrossRef]
- Koster, T.; Marondedze, C.; Meyer, K.; Staiger, D. RNA-Binding Proteins Revisited—The Emerging Arabidopsis mRNA Interactome. Trends Plant Sci. 2017, 22, 512–526. [Google Scholar] [CrossRef]
- Maccarrone, G.; Bonfiglio, J.J.; Silberstein, S.; Turck, C.W.; Martins-de-Souza, D. Characterization of a Protein Interactome by Co-Immunoprecipitation and Shotgun Mass Spectrometry. In Multiplex Biomarker Techniques: Methods and Applications; Guest, P.C., Ed.; Springer: New York, NY, USA, 2017; pp. 223–234. [Google Scholar] [CrossRef]
- Ghadie, M.A.; Coulombe-Huntington, J.; Xia, Y. Interactome evolution: Insights from genome-wide analyses of protein-protein interactions. Curr. Opin. Struct. Biol. 2018, 50, 42–48. [Google Scholar] [CrossRef]
- Shukla, E.; Chauhan, R. Host-HIV-1 Interactome: A Quest for Novel Therapeutic Intervention. Cells 2019, 8, 1155. [Google Scholar] [CrossRef] [PubMed]
- Park, D.I.; Turck, C.W. Interactome Studies of Psychiatric Disorders. In Reviews on Biomarker Studies in Psychiatric and Neurodegenerative Disorders; Guest, P.C., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 163–173. [Google Scholar] [CrossRef]
- Subramanian, I.; Verma, S.; Kumar, S.; Jere, A.; Anamika, K. Multi-omics Data Integration, Interpretation, and Its Application. Bioinform. Biol. Insights 2020, 14. [Google Scholar] [CrossRef] [PubMed]
- Ulfenborg, B. Vertical and horizontal integration of multi-omics data with miodin. BMC Bioinform. 2019, 20, 649. [Google Scholar] [CrossRef] [PubMed]
- Bersanelli, M.; Mosca, E.; Remondini, D.; Giampieri, E.; Sala, C.; Castellani, G.; Milanesi, L. Methods for the integration of multi-omics data: Mathematical aspects. BMC Bioinform. 2016, 17, S15. [Google Scholar] [CrossRef]
- Canzler, S.; Schor, J.; Busch, W.; Schubert, K.; Rolle-Kampczyk, U.E.; Seitz, H.; Kamp, H.; von Bergen, M.; Buesen, R.; Hackermüller, J. Prospects and challenges of multi-omics data integration in toxicology. Arch. Toxicol. 2020, 94, 371–388. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, H.; White, J.C.; Chen, X.; Li, H.; Qu, X.; Ji, R. Metabolomics reveals that engineered nanomaterial exposure in soil alters both soil rhizosphere metabolite profiles and maize metabolic pathways. Environ. Sci. Nano 2019, 6, 1716–1727. [Google Scholar] [CrossRef]
- Walker, D.I.; Lane, K.J.; Liu, K.; Uppal, K.; Patton, A.P.; Durant, J.L.; Jones, D.P.; Brugge, D.; Pennell, K.D. Metabolomic assessment of exposure to near-highway ultrafine particles. J. Expo. Sci. Environ. Epidemiol. 2019, 29, 469–483. [Google Scholar] [CrossRef]
- Houten, S.M.; Chen, J.; Belpoggi, F.; Manservisi, F.; Sanchez-Guijo, A.; Wudy, S.A.; Teitelbaum, S.L. Changes in the Metabolome in Response to Low-Dose Exposure to Environmental Chemicals Used in Personal Care Products during Different Windows of Susceptibility. PLoS ONE 2016, 11, e0159919. [Google Scholar] [CrossRef]
- Van Meter, R.J.; Glinski, D.A.; Purucker, S.T.; Henderson, W.M. Influence of exposure to pesticide mixtures on the metabolomic profile in post-metamorphic green frogs (Lithobates clamitans). Sci. Total Environ. 2018, 624, 1348–1359. [Google Scholar] [CrossRef]
- Grison, S.; Fave, G.; Maillot, M.; Manens, L.; Delissen, O.; Blanchardon, E.; Dublineau, I.; Aigueperse, J.; Bohand, S.; Martin, J.C.; et al. Metabolomics reveals dose effects of low-dose chronic exposure to uranium in rats: Identification of candidate biomarkers in urine samples. Metabolomics 2016, 12, 154. [Google Scholar] [CrossRef]
- Golder, H.M.; Thomson, J.M.; Denman, S.E.; McSweeney, C.S.; Lean, I.J. Genetic Markers Are Associated with the Ruminal Microbiome and Metabolome in Grain and Sugar Challenged Dairy Heifers. Front. Genet. 2018, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Ma, M.P.; Diao, Q.Y.; Tu, Y. Saponin-Induced Shifts in the Rumen Microbiome and Metabolome of Young Cattle. Front. Microbiol. 2019, 10, 356. [Google Scholar] [CrossRef] [PubMed]
- Myer, P.R. Bovine Genome-Microbiome Interactions: Metagenomic Frontier for the Selection of Efficient Productivity in Cattle Systems. Msystems 2019, 4, e00103–e00119. [Google Scholar] [CrossRef]
- Patterson, D.S.P.; Shreeve, B.J.; Roberts, B.A.; Berrett, S.; Brush, P.J.; Glancy, E.M.; Krogh, P. Effect on calves of barley naturally contaminated with ochratoxin A and groundnut meal contaminated with low concentrations of aflatoxin R1. Res. Vet. Sci. 1981, 31, 213–218. [Google Scholar] [CrossRef]
- Cook, W.O.; Richard, J.L.; Osweiler, G.D.; Trampel, D.W. Clinical and pathologic changes in acute bovine aflatoxicosis: Rumen motility and tissue and fluid concentrations of aflatoxins B1 and M1. Am. J. Vet. Res. 1986, 47, 1817–1825. [Google Scholar]
- Placinta, C.M.; D’Mello, J.P.F.; Macdonald, A.M.C. A review of worldwide contamination of cereal grains and animal feed with Fusarium mycotoxins. Anim. Feed Sci. Technol. 1999, 78, 21–37. [Google Scholar] [CrossRef]
- Klotz, J.L. Activities and Effects of Ergot Alkaloids on Livestock Physiology and Production. Toxins 2015, 7, 2801–2821. [Google Scholar] [CrossRef]
- Li, F.; Li, C.; Chen, Y.; Liu, J.; Zhang, C.; Irving, B.; Fitzsimmons, C.; Plastow, G.; Guan, L.L. Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle. Microbiome 2019, 7, 92. [Google Scholar] [CrossRef]
- Weimer, P.J.; Stevenson, D.M.; Mantovani, H.C.; Man, S.L. Host specificity of the ruminal bacterial community in the dairy cow following near-total exchange of ruminal contents. J. Dairy Sci. 2010, 93, 5902–5912. [Google Scholar] [CrossRef]
- Delgado, B.; Bach, A.; Guasch, I.; Gonzalez, C.; Elcoso, G.; Pryce, J.E.; Gonzalez-Recio, O. Whole rumen metagenome sequencing allows classifying and predicting feed efficiency and intake levels in cattle. Sci. Rep. 2019, 9, 11. [Google Scholar] [CrossRef]
- Lima, J.; Auffret, M.D.; Stewart, R.D.; Dewhurst, R.J.; Duthie, C.A.; Snelling, T.J.; Walker, A.W.; Freeman, T.C.; Watson, M.; Roehe, R. Identification of Rumen Microbial Genes Involved in Pathways Linked to Appetite, Growth, and Feed Conversion Efficiency in Cattle. Front. Genet. 2019, 10, 701. [Google Scholar] [CrossRef]
- Jewell, K.A.; McCormick, C.A.; Odt, C.L.; Weimer, P.J.; Suen, G. Ruminal Bacterial Community Composition in Dairy Cows Is Dynamic over the Course of Two Lactations and Correlates with Feed Efficiency. Appl. Environ. Microbiol. 2015, 81, 4697–4710. [Google Scholar] [CrossRef] [PubMed]
- Belanche, A.; Kingston-Smith, A.H.; Griffith, G.W.; Newbold, C.J. A Multi-Kingdom Study Reveals the Plasticity of the Rumen Microbiota in Response to a Shift From Non-grazing to Grazing Diets in Sheep. Front. Microbiol. 2019, 10, 122. [Google Scholar] [CrossRef] [PubMed]
- Khafipour, E.; Li, S.; Tun, H.M.; Derakhshani, H.; Moossavi, S.; Plaizier, J.C. Effects of grain feeding on microbiota in the digestive tract of cattle. Anim. Front. 2016, 6, 13–19. [Google Scholar] [CrossRef]
- Mao, S.; Zhang, M.; Liu, J.; Zhu, W. Characterising the bacterial microbiota across the gastrointestinal tracts of dairy cattle: Membership and potential function. Sci. Rep. 2015, 5, 16116. [Google Scholar] [CrossRef]
- de Oliveira, M.N.; Jewell, K.A.; Freitas, F.S.; Benjamin, L.A.; Totola, M.R.; Borges, A.C.; Moraes, C.A.; Suen, G. Characterizing the microbiota across the gastrointestinal tract of a Brazilian Nelore steer. Vet. Microbiol. 2013, 164, 307–314. [Google Scholar] [CrossRef]
- O’Hara, E.; Kenny, D.A.; McGovern, E.; Byrne, C.J.; McCabe, M.S.; Guan, L.L.; Waters, S.M. Investigating temporal microbial dynamics in the rumen of beef calves raised on two farms during early life. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef]
- Artegoitia, V.M.; Foote, A.P.; Lewis, R.M.; Freetly, H.C. Rumen Fluid Metabolomics Analysis Associated with Feed Efficiency on Crossbred Steers. Sci. Rep. 2017, 7, 2864. [Google Scholar] [CrossRef]
- Boudonck, K.J.; Mitchell, M.W.; Wulff, J.; Ryals, J.A. Characterization of the biochemical variability of bovine milk using metabolomics. Metabolomics 2009, 5, 375–386. [Google Scholar] [CrossRef]
- Klein, M.S.; Almstetter, M.F.; Schlamberger, G.; Nurnberger, N.; Dettmer, K.; Oefner, P.J.; Meyer, H.H.; Wiedemann, S.; Gronwald, W. Nuclear magnetic resonance and mass spectrometry-based milk metabolomics in dairy cows during early and late lactation. J. Dairy Sci. 2010, 93, 1539–1550. [Google Scholar] [CrossRef]
- Goldansaz, S.A.; Guo, A.C.; Sajed, T.; Steele, M.A.; Plastow, G.S.; Wishart, D.S. Livestock metabolomics and the livestock metabolome: A systematic review. PLoS ONE 2017, 12, e0177675. [Google Scholar] [CrossRef] [PubMed]
- Leng, R.A. Biofilm compartmentalisation of the rumen microbiome: Modification of fermentation and degradation of dietary toxins. Anim. Prod. Sci. 2017, 57, 2188–2203. [Google Scholar] [CrossRef]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef] [PubMed]
- Ogunade, I.; Jiang, Y.; Pech Cervantes, A. DI/LC-MS/MS-Based Metabolome Analysis of Plasma Reveals the Effects of Sequestering Agents on the Metabolic Status of Dairy Cows Challenged with Aflatoxin B1. Toxins 2019, 11, 693. [Google Scholar] [CrossRef]
- Ogunade, I.; Jiang, Y.; Adeyemi, J.; Oliveira, A.; Vyas, D.; Adesogan, A. Biomarker of Aflatoxin Ingestion: (1)H NMR-Based Plasma Metabolomics of Dairy Cows Fed Aflatoxin B(1) with or without Sequestering Agents. Toxins 2018, 10, 545. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Zheng, N.; Guo, L.; Song, X.; Zhao, S.; Wang, J. Biological System Responses of Dairy Cows to Aflatoxin B1 Exposure Revealed with Metabolomic Changes in Multiple Biofluids. Toxins 2019, 11, 77. [Google Scholar] [CrossRef]
- Piotrowska, M.; Slizewska, K.; Nowak, A.; Zielonka, L.; Zakowska, Z.; Gajecka, M.; Gajecki, M. The effect of experimental fusarium mycotoxicosis on microbiota diversity in porcine ascending colon contents. Toxins 2014, 6, 2064–2081. [Google Scholar] [CrossRef]
- Shaffer, M.; Armstrong, A.J.S.; Phelan, V.V.; Reisdorph, N.; Lozupone, C.A. Microbiome and metabolome data integration provides insight into health and disease. Transl. Res. 2017, 189, 51–64. [Google Scholar] [CrossRef]
- Lu, K.; Abo, R.P.; Schlieper, K.A.; Graffam, M.E.; Levine, S.; Wishnok, J.S.; Swenberg, J.A.; Tannenbaum, S.R.; Fox, J.G. Arsenic exposure perturbs the gut microbiome and its metabolic profile in mice: An integrated metagenomics and metabolomics analysis. Environ. Health Perspect. 2014, 122, 284–291. [Google Scholar] [CrossRef]
- Yang, Y.; Misra, B.B.; Liang, L.; Bi, D.; Weng, W.; Wu, W.; Cai, S.; Qin, H.; Goel, A.; Li, X.; et al. Integrated microbiome and metabolome analysis reveals a novel interplay between commensal bacteria and metabolites in colorectal cancer. Theranostics 2019, 9, 4101–4114. [Google Scholar] [CrossRef]
- Tang, Z.-Z.; Chen, G.; Hong, Q.; Huang, S.; Smith, H.M.; Shah, R.D.; Scholz, M.; Ferguson, J.F. Multi-Omic Analysis of the Microbiome and Metabolome in Healthy Subjects Reveals Microbiome-Dependent Relationships Between Diet and Metabolites. Front. Genet. 2019, 10, 454. [Google Scholar] [CrossRef] [PubMed]
- Mote, R.S.; Hill, N.S.; Skarlupka, J.H.; Turner, Z.B.; Sanders, Z.P.; Jones, D.P.; Suen, G.; Filipov, N.M. Response of Beef Cattle Fecal Microbiota to Grazing on Toxic Tall Fescue. Appl. Environ. Microbiol. 2019, 85, e00032-19. [Google Scholar] [CrossRef] [PubMed]
- Mote, R.S.; Hill, N.S.; Uppal, K.; Tran, V.T.; Jones, D.P.; Filipov, N.M. Metabolomics of fescue toxicosis in grazing beef steers. Food Chem. Toxicol. 2017, 105, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Mote, R.S.; Hill, N.S.; Skarlupka, J.H.; Tran, V.T.; Walker, D.I.; Turner, Z.B.; Sanders, Z.P.; Jones, D.P.; Suen, G.; Filipov, N.M. Toxic tall fescue grazing increases susceptibility of the Angus steer fecal microbiota and plasma/urine metabolome to environmental effects. Sci. Rep. 2020, 10, 2497. [Google Scholar] [CrossRef]
- Melchior, E.A.; Myer, P.R. Fescue toxicosis and its influence on the rumen microbiome: Mitigation of production losses through clover isoflavones. J. Appl. Anim. Res. 2018, 46, 1280–1288. [Google Scholar] [CrossRef]
- Melchior, E.A.; Smith, J.K.; Schneider, L.G.; Mulliniks, J.T.; Bates, G.E.; McFarlane, Z.D.; Flythe, M.D.; Klotz, J.L.; Goodman, J.P.; Ji, H.; et al. Effects of red clover isoflavones on tall fescue seed fermentation and microbial populations in vitro. PLoS ONE 2018, 13, e0201866. [Google Scholar] [CrossRef]
- Clay, K.; Schardl, C. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat. 2002, 160 (Suppl. 4), S99–S127. [Google Scholar] [CrossRef]
- Christensen, M.J.; Bennett, R.J.; Ansari, H.A.; Koga, H.; Johnson, R.D.; Bryan, G.T.; Simpson, W.R.; Koolaard, J.P.; Nickless, E.M.; Voisey, C.R. Epichloe endophytes grow by intercalary hyphal extension in elongating grass leaves. Fungal Genet. Biol. 2008, 45, 84–93. [Google Scholar] [CrossRef]
- Schardl, C.L.; Young, C.A.; Pan, J.; Florea, S.; Takach, J.E.; Panaccione, D.G.; Farman, M.L.; Webb, J.S.; Jaromczyk, J.; Charlton, N.D.; et al. Currencies of mutualisms: Sources of alkaloid genes in vertically transmitted epichloae. Toxins 2013, 5, 1064–1088. [Google Scholar] [CrossRef]
- Missaoui, A.; Hill, N. Use of accelerated aging as a surrogate phenotyping approach to improve endophyte survival during storage of tall fescue seed. Field Crop. Res. 2015, 183, 43–49. [Google Scholar] [CrossRef]
- Guerre, P. Ergot alkaloids produced by endophytic fungi of the genus Epichloe. Toxins 2015, 7, 773–790. [Google Scholar] [CrossRef] [PubMed]
- Young, C.A.; Charlton, N.D.; Takach, J.E.; Swoboda, G.A.; Trammell, M.A.; Huhman, D.V.; Hopkins, A.A. Characterization of Epichloe coenophiala within the US: Are all tall fescue endophytes created equal? Front. Chem. 2014, 2, 95. [Google Scholar] [CrossRef] [PubMed]
- Maruo, V.M.; Bracarense, A.P.; Metayer, J.P.; Vilarino, M.; Oswald, I.P.; Pinton, P. Ergot Alkaloids at Doses Close to EU Regulatory Limits Induce Alterations of the Liver and Intestine. Toxins 2018, 10, 183. [Google Scholar] [CrossRef]
- Philippe, G. Lolitrem B and Indole Diterpene Alkaloids Produced by Endophytic Fungi of the Genus Epichloe and Their Toxic Effects in Livestock. Toxins 2016, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Berde, B.; Stürmer, E. Introduction to the Pharmacology of Ergot Alkaloids and Related Compounds as a Basis of Their Therapeutic Application; Springer: Berlin/Heidelberg, Germany, 1978; pp. 1–28. [Google Scholar] [CrossRef]
- Moubarak, A.S.; Piper, E.L.; Johnson, Z.B.; Flieger, M. HPLC Method for Detection of Ergotamine, Ergosine, and Ergine after Intravenous Injection of a Single Dose. J. Agric. Food Chem. 1996, 44, 146–148. [Google Scholar] [CrossRef]
- Hill, N.S.; Thompson, F.N.; Stuedemann, J.A.; Dawe, D.L.; Hiatt, E.E., 3rd. Urinary alkaloid excretion as a diagnostic tool for fescue toxicosis in cattle. J. Vet. Diagn. Investig. 2000, 12, 210–217. [Google Scholar] [CrossRef]
- Stuedemann, J.A.; Hill, N.S.; Thompson, F.N.; Fayrer-Hosken, R.A.; Hay, W.P.; Dawe, D.L.; Seman, D.H.; Martin, S.A. Urinary and biliary excretion of ergot alkaloids from steers that grazed endophyte-infected tall fescue. J. Anim. Sci. 1998, 76, 2146–2154. [Google Scholar] [CrossRef]
- Hill, N.S.; Thompson, F.N.; Stuedemann, J.A.; Rottinghaus, G.W.; Ju, H.J.; Dawe, D.L.; Hiatt, E.E., 3rd. Ergot alkaloid transport across ruminant gastric tissues. J. Anim. Sci. 2001, 79, 542–549. [Google Scholar] [CrossRef]
- Realini, C.E.; Duckett, S.K.; Hill, N.S.; Hoveland, C.S.; Lyon, B.G.; Sackmann, J.R.; Gillis, M.H. Effect of endophyte type on carcass traits, meat quality, and fatty acid composition of beef cattle grazing tall fescue. J. Anim. Sci. 2005, 83, 430–439. [Google Scholar] [CrossRef]
- Zbib, N.; Repussard, C.; Tardieu, D.; Priymenko, N.; Domange, C.; Guerre, P. Toxicity of endophyte-infected ryegrass hay containing high ergovaline level in lactating ewes. J. Anim. Sci. 2015, 93, 4098–4109. [Google Scholar] [CrossRef]
- Westendorf, M.L.; Mitchell, G.E., Jr.; Tucker, R.E.; Bush, L.P.; Petroski, R.J.; Powell, R.G. In vitro and in vivo ruminal and physiological responses to endophyte-infected tall fescue. J Dairy Sci. 1993, 76, 555–563. [Google Scholar] [CrossRef]
- Ayers, A.W.; Hill, N.S.; Rottinghaus, G.E.; Stuedemann, J.A.; Thompson, F.N.; Purinton, P.T.; Seman, D.H.; Dawe, D.L.; Parks, A.H.; Ensley, D. Ruminal Metabolism and Transport of Tall Fescue Ergot Alkaloids. Crop Sci. 2009, 49, 2309–2316. [Google Scholar] [CrossRef]
- Strimbu, K.; Tavel, J.A. What are biomarkers? Curr. Opin. HIV AIDS 2010, 5, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Mayeux, R. Biomarkers: Potential uses and limitations. NeuroRx 2004, 1, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Hurley, W.L.; Convey, E.M.; Leung, K.; Edgerton, L.A.; Hemken, R.W. Bovine prolactin, TSH, T and T concentrations as affected by tall fescue summer toxicosis and temperature. J. Anim. Sci. 1980, 51, 374–379. [Google Scholar] [CrossRef]
- Karg, H.; Schams, D. Prolactin release in cattle. J. Reprod. Fertil. 1974, 39, 463–472. [Google Scholar] [CrossRef]
- Smith, V.G.; Hacker, R.R.; Brown, R.G. Effect of alterations in ambient temperature on serum prolactin concentration in steers. J. Anim. Sci. 1977, 44, 645–649. [Google Scholar] [CrossRef]
- Bryant, G.D.; Linzell, J.L.; Greenwood, F.C. Plasma prolactin in goats measured by radioimmunoassay: The effects of teat stimulation, mating behavior, stress, fasting and of oxytocin, insulin and glucose injections. Hormones 1970, 1, 26–35. [Google Scholar] [CrossRef]
- Yayou, K.; Ito, S.; Yamamoto, N.; Kitagawa, S.; Okamura, H. Relationships of stress responses with plasma oxytocin and prolactin in heifer calves. Physiol. Behav. 2010, 99, 362–369. [Google Scholar] [CrossRef]
- Cunningham, I. Tall fescue grass is poison for cattle. N. Z. J. Agric. 1948, 77, 519. [Google Scholar]
- Cunningham, I.J. A Note on the Cause of Tall Fescue Lameness in Cattle. Aust. Vet. J. 1949, 25, 27–28. [Google Scholar] [CrossRef]
- Jacobson, D.R.; Miller, W.M.; Seath, D.M.; Yates, S.G.; Tookey, H.L.; Wolff, I.A. Nature of Fescue Toxicity and Progress toward Identification of the Toxic Entity. J. Dairy Sci. 1963, 46, 416–422. [Google Scholar] [CrossRef]
- Paterson, J.; Forcherio, C.; Larson, B.; Samford, M.; Kerley, M. The effects of fescue toxicosis on beef cattle productivity. J. Anim. Sci. 1995, 73, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Strickland, J.R.; Oliver, J.W.; Cross, D.L. Fescue toxicosis and its impact on animal agriculture. Vet. Hum. Toxicol. 1993, 35, 454–464. [Google Scholar] [PubMed]
- Snider, M.A.; Harmon, D.L.; Klotz, J.L. Pharmacologic assessment of bovine ruminal and mesenteric vascular serotonin receptor populations. J. Anim. Sci. 2018, 96, 1570–1578. [Google Scholar] [CrossRef] [PubMed]
- Trotta, R.J.; Harmon, D.L.; Klotz, J.L. Interaction of ergovaline with serotonin receptor 5-HT2A in bovine ruminal and mesenteric vasculature. J. Anim. Sci. 2018, 96, 4912–4922. [Google Scholar] [CrossRef]
- McLeay, L.M.; Smith, B.L. Effects of ergotamine and ergovaline on the electromyographic activity of smooth muscle of the reticulum and rumen of sheep. Am. J. Vet. Res. 2006, 67, 707–714. [Google Scholar] [CrossRef]
- Foote, A.P.; Kristensen, N.B.; Klotz, J.L.; Kim, D.H.; Koontz, A.F.; McLeod, K.R.; Bush, L.P.; Schrick, F.N.; Harmon, D.L. Ergot alkaloids from endophyte-infected tall fescue decrease reticuloruminal epithelial blood flow and volatile fatty acid absorption from the washed reticulorumen. J. Anim. Sci. 2013, 91, 5366–5378. [Google Scholar] [CrossRef]
- Koontz, A.F.; Kim, D.H.; McLeod, K.R.; Klotz, J.L.; Harmon, D.L. Effect of fescue toxicosis on whole body energy and nitrogen balance, in situ degradation and ruminal passage rates in Holstein steers. Anim. Prod. Sci. 2015, 55, 988–998. [Google Scholar] [CrossRef]
- Koontz, A.F.; Bush, L.P.; Klotz, J.L.; McLeod, K.R.; Schrick, F.N.; Harmon, D.L. Evaluation of a ruminally dosed tall fescue seed extract as a model for fescue toxicosis in steers. J. Anim. Sci. 2012, 90, 914–921. [Google Scholar] [CrossRef]
- Koontz, A.F.; Kim, D.H.; Foote, A.P.; Bush, L.P.; Klotz, J.L.; McLeod, K.R.; Harmon, D.L. Alteration of fasting heat production during fescue toxicosis in Holstein steers. J. Anim. Sci. 2013, 91, 3881–3888. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.S. Physical constraints on voluntary intake of forages by ruminants. J. Anim. Sci. 1996, 74, 3063–3075. [Google Scholar] [CrossRef] [PubMed]
- Felitti, S.; Shields, K.; Ramsperger, M.; Tian, P.; Sawbridge, T.; Webster, T.; Logan, E.; Erwin, T.; Forster, J.; Edwards, D.; et al. Transcriptome analysis of Neotyphodium and Epichloe grass endophytes. Fungal Genet. Biol. 2006, 43, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; McCulley, R.L.; McNear, D.H., Jr. Tall fescue cultivar and fungal endophyte combinations influence plant growth and root exudate composition. Front. Plant. Sci. 2015, 6, 183. [Google Scholar] [CrossRef]
- Rojas, X.; Guo, J.; Leff, J.W.; McNear, D.H., Jr.; Fierer, N.; McCulley, R.L. Infection with a Shoot-Specific Fungal Endophyte (Epichloe) Alters Tall Fescue Soil Microbial Communities. Microb. Ecol. 2016, 72, 197–206. [Google Scholar] [CrossRef]
- Rasmussen, S.; Parsons, A.J.; Newman, J.A. Metabolomics analysis of the Lolium perenne–Neotyphodium lolii symbiosis: More than just alkaloids? Phytochem. Rev. 2009, 8, 535–550. [Google Scholar] [CrossRef]
- Bastias, D.A.; Martinez-Ghersa, M.A.; Ballare, C.L.; Gundel, P.E. Epichloe Fungal Endophytes and Plant Defenses: Not Just Alkaloids. Trends Plant Sci. 2017, 22, 939–948. [Google Scholar] [CrossRef]
- Miller, M.J.; McDole, J.R.; Newberry, R.D. Microanatomy of the intestinal lymphatic system. Ann. N. Y. Acad. Sci. 2010, 1207 (Suppl. 1), E21–E28. [Google Scholar] [CrossRef]
- Unthank, J.L.; Bohlen, H.G. Lymphatic pathways and role of valves in lymph propulsion from small intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 1988, 254, G389–G398. [Google Scholar] [CrossRef]
- Futrell, M.C.; Farnell, D.R.; Poe, W.E.; Watson, V.H.; Coats, R.E. Fungal Populations in the Rumen Associated with Fescue Toxicosis. J. Environ. Qual. 1974, 3, 140–143. [Google Scholar] [CrossRef]
- Eich, E.; Eichberg, D.; Muller, W.E. Clavines. New antibiotics with cytostatic activity. Biochem. Pharmacol. 1984, 33, 523–526. [Google Scholar] [CrossRef]
- Eich, E.; Eichberg, D.; Schwarz, G.; Clas, F.; Loos, M. Antimicrobial activitiy of clavines. Arzeimittelforschung 1985, 35, 1760–1762. [Google Scholar]
- Looper, M.L.; Edrington, T.S.; Flores, R.; Burke, J.M.; Callaway, T.R.; Aiken, G.E.; Schrick, F.N.; Rosenkrans, C.F., Jr. Influence of dietary endophyte (Neotyphodium coenophialum)-infected tall fescue (Festuca arundinacea) seed on fecal shedding of antibiotic resistance-selected Escherichia coli O157:H7 in ewes. J. Anim. Sci. 2007, 85, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Harlow, B.E.; Goodman, J.P.; Lynn, B.C.; Flythe, M.D.; Ji, H.; Aiken, G.E. Ruminal tryptophan-utilizing bacteria degrade ergovaline from tall fescue seed extract. J. Anim. Sci. 2017, 95, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Alrashedi, S. Effect of Endophyte-Infected Tall Fescue Toxins on Growth Performance and the Microbial Community in the Rumen and Feces in Pregnant Ewes. University of Arkansas. 2017. Available online: http://scholarworks.uark.edu/etd/2532 (accessed on 2 April 2020).
- Koester, L.; Poole, D.; Serão, N.; Schmitz-Esser, S. Beef cattle that respond differently to fescue toxicosis have distinct gastrointestinal tract microbiota. PLoS ONE 2020, 15, e0229192. [Google Scholar] [CrossRef]
- Strickland, J.; Aiken, G.E.; Spiers, D.; Fletcher, L.R.; Oliver, J. Physiological Basis of Fescue Toxicosis. In Tall Fescue for the Twenty-First Century; American Society of Agronomy: Madison, WI, USA, 2009; Volume 53, pp. 203–227. [Google Scholar] [CrossRef]
- Oliver, J.W.; Schultze, A.; Rohrbach, B.W.; Fribourg, H.; Ingle, T.; Waller, J. Alterations in hemograms and serum biochemical analytes of steers after prolonged consumption of endophyte-infected tall fescue. J. Anim. Sci. 2000, 78, 1029–1035. [Google Scholar] [CrossRef]
- Browning, R., Jr.; Gissendanner, S.J.; Wakefield, T., Jr. Ergotamine alters plasma concentrations of glucagon, insulin, cortisol, and triiodothyronine in cows. J. Anim. Sci. 2000, 78, 690–698. [Google Scholar] [CrossRef]
- Coufal-Majewski, S.; Stanford, K.; McAllister, T.; Blakley, B.; McKinnon, J.; Chaves, A.V.; Wang, Y. Impacts of Cereal Ergot in Food Animal Production. Front. Vet. Sci. 2016, 3, 15. [Google Scholar] [CrossRef]
- Schmidt, S.P.; Hoveland, C.S.; Clark, E.M.; Davis, N.D.; Smith, L.A.; Grimes, H.W.; Holliman, J.L. Association of an endophytic fungus with fescue toxicity in steers fed Kentucky 31 tall fescue seed or hay. J. Anim. Sci. 1982, 55, 1259–1263. [Google Scholar] [CrossRef]
- Patterson, J.; Looper, M.C.; Williamson, B.; Rosenkrans, C. Effects of fescue cultivar and heat shock protein haplotype on growth and fertility of crossbred beef heifers. Anim. Sci. Ark. Anim. Sci. 2011, 597, 57–59. [Google Scholar]
- Jackson, J.J.; Lindemann, M.D.; Boling, J.A.; Matthews, J.C. Summer-Long Grazing of High vs. Low Endophyte (Neotyphodium coenophialum)-Infected Tall Fescue by Growing Beef Steers Results in Distinct Temporal Blood Analyte Response Patterns, with Poor Correlation to Serum Prolactin Levels. Front. Vet. Sci. 2015, 2, 77. [Google Scholar] [CrossRef] [PubMed]
- Amar, J. Microbiota-Host Crosstalk: A Bridge Between Cardiovascular Risk Factors, Diet, and Cardiovascular Disease. Am. J. Hypertens. 2018, 31, 941–944. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, S.M.; Lagrou, K.; Duarte-Oliveira, C.; Maertens, J.A.; Cunha, C.; Carvalho, A. The microbiome-metabolome crosstalk in the pathogenesis of respiratory fungal diseases. Virulence 2017, 8, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Garcia, A.; Pineda-Quiroga, C.; Atxaerandio, R.; Perez, A.; Hernandez, I.; Garcia-Rodriguez, A.; Gonzalez-Recio, O. Comparison of Mothur and QIIME for the Analysis of Rumen Microbiota Composition Based on 16S rRNA Amplicon Sequences. Front. Microbiol. 2018, 9, 3010. [Google Scholar] [CrossRef] [PubMed]
- Prodan, A.; Tremaroli, V.; Brolin, H.; Zwinderman, A.H.; Nieuwdorp, M.; Levin, E. Comparing bioinformatic pipelines for microbial 16S rRNA amplicon sequencing. PLoS ONE 2020, 15, e0227434. [Google Scholar] [CrossRef]
- Lee, M.Y.; Hu, T. Computational Methods for the Discovery of Metabolic Markers of Complex Traits. Metabolites 2019, 9, 66. [Google Scholar] [CrossRef]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies-Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef]
- Carroll, A.J.; Zhang, P.; Whitehead, L.; Kaines, S.; Tcherkez, G.; Badger, M.R. PhenoMeter: A Metabolome Database Search Tool Using Statistical Similarity Matching of Metabolic Phenotypes for High-Confidence Detection of Functional Links. Front. Bioeng. Biotechnol. 2015, 3, 106. [Google Scholar] [CrossRef]
- Longnecker, K.; Futrelle, J.; Coburn, E.; Kido Soule, M.C.; Kujawinski, E.B. Environmental metabolomics: Databases and tools for data analysis. Mar. Chem. 2015, 177, 366–373. [Google Scholar] [CrossRef]
- Kumar, A.; Pathak, R.K.; Gupta, S.M.; Gaur, V.S.; Pandey, D. Systems Biology for Smart Crops and Agricultural Innovation: Filling the Gaps between Genotype and Phenotype for Complex Traits Linked with Robust Agricultural Productivity and Sustainability. OMICS 2015, 19, 581–601. [Google Scholar] [CrossRef]
- Shameer, K.; Naika, M.B.N.; Shafi, K.M.; Sowdhamini, R. Decoding systems biology of plant stress for sustainable agriculture development and optimized food production. Prog. Biophys. Mol. Biol. 2019, 145, 19–39. [Google Scholar] [CrossRef] [PubMed]
- de Menezes, A.B.; Lewis, E.; O’Donovan, M.; O’Neill, B.F.; Clipson, N.; Doyle, E.M. Microbiome analysis of dairy cows fed pasture or total mixed ration diets. FEMS Microbiol. Ecol. 2011, 78, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Pitta, D.W.; Kumar, S.; Vecchiarelli, B.; Shirley, D.J.; Bittinger, K.; Baker, L.D.; Ferguson, J.D.; Thomsen, N. Temporal dynamics in the ruminal microbiome of dairy cows during the transition period. J. Anim. Sci. 2014, 92, 4014–4022. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Zolla, L. Meat science: From proteomics to integrated omics towards system biology. J. Proteom. 2013, 78, 558–577. [Google Scholar] [CrossRef] [PubMed]
- Fecteau, M.E.; Pitta, D.W.; Vecchiarelli, B.; Indugu, N.; Kumar, S.; Gallagher, S.C.; Fyock, T.L.; Sweeney, R.W. Dysbiosis of the Fecal Microbiota in Cattle Infected with Mycobacterium avium subsp. paratuberculosis. PLoS ONE 2016, 11, e0160353. [Google Scholar] [CrossRef]
- Iheshiulor, O.O.M.; Esonu, B.O.; Chuwuka, O.K.; Omede, A.A.; Okoli, I.C.; Ogbuewu, I.P. Effects of Mycotoxins in Animal Nutrition: A Review. Asian J. Anim. Sci. 2011, 5, 19–33. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mote, R.S.; Filipov, N.M. Use of Integrative Interactomics for Improvement of Farm Animal Health and Welfare: An Example with Fescue Toxicosis. Toxins 2020, 12, 633. https://doi.org/10.3390/toxins12100633
Mote RS, Filipov NM. Use of Integrative Interactomics for Improvement of Farm Animal Health and Welfare: An Example with Fescue Toxicosis. Toxins. 2020; 12(10):633. https://doi.org/10.3390/toxins12100633
Chicago/Turabian StyleMote, Ryan S., and Nikolay M. Filipov. 2020. "Use of Integrative Interactomics for Improvement of Farm Animal Health and Welfare: An Example with Fescue Toxicosis" Toxins 12, no. 10: 633. https://doi.org/10.3390/toxins12100633
APA StyleMote, R. S., & Filipov, N. M. (2020). Use of Integrative Interactomics for Improvement of Farm Animal Health and Welfare: An Example with Fescue Toxicosis. Toxins, 12(10), 633. https://doi.org/10.3390/toxins12100633