Calcium-Induced Activity and Folding of a Repeat in Toxin Lipase from Antarctic Pseudomonas fluorescens Strain AMS8

 , and
, and
Abstract
1. Introduction
2. Results and Discussion
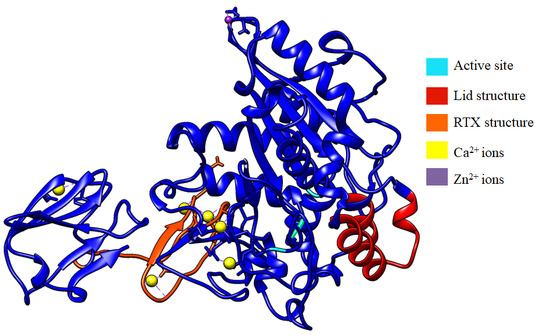
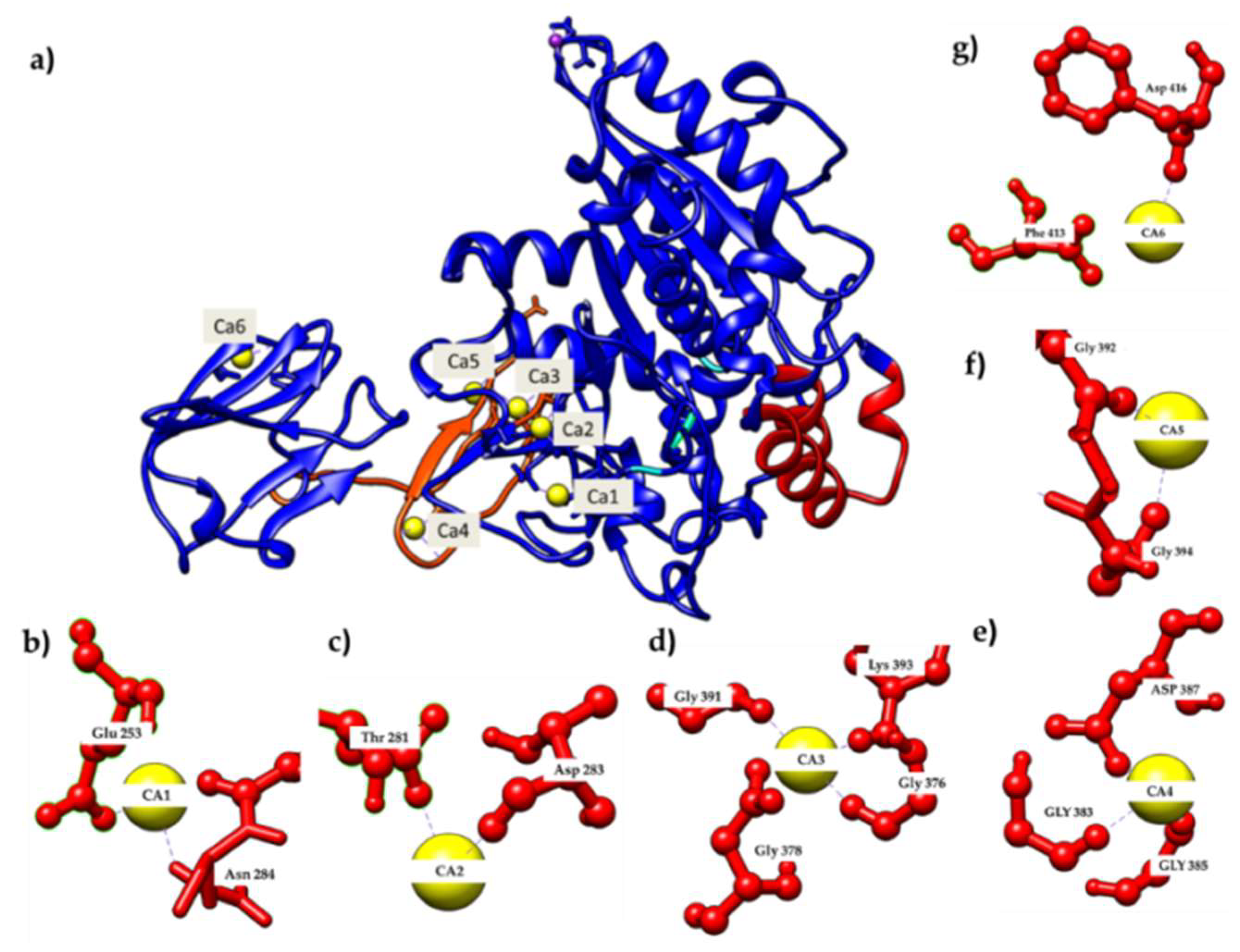
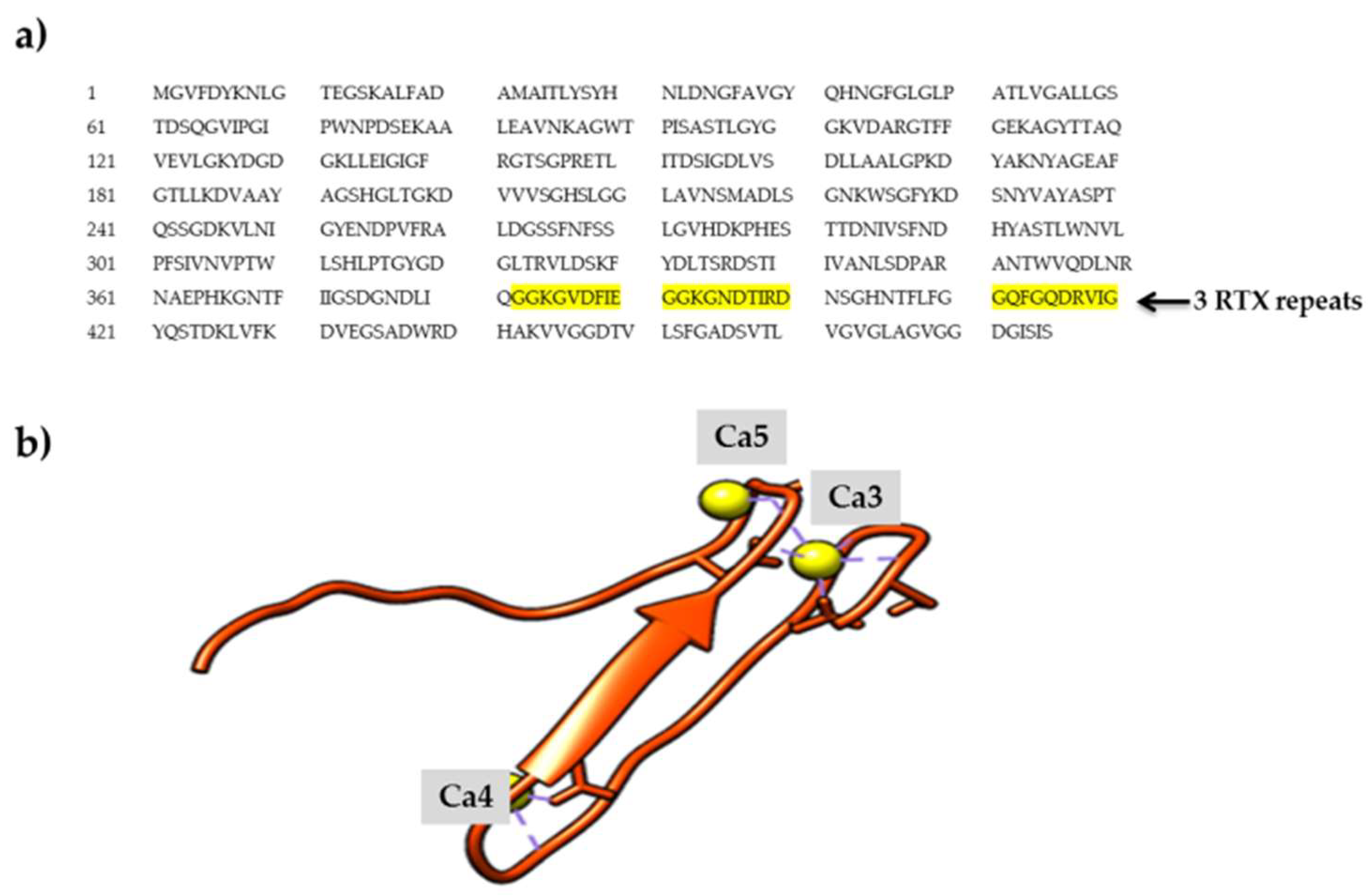
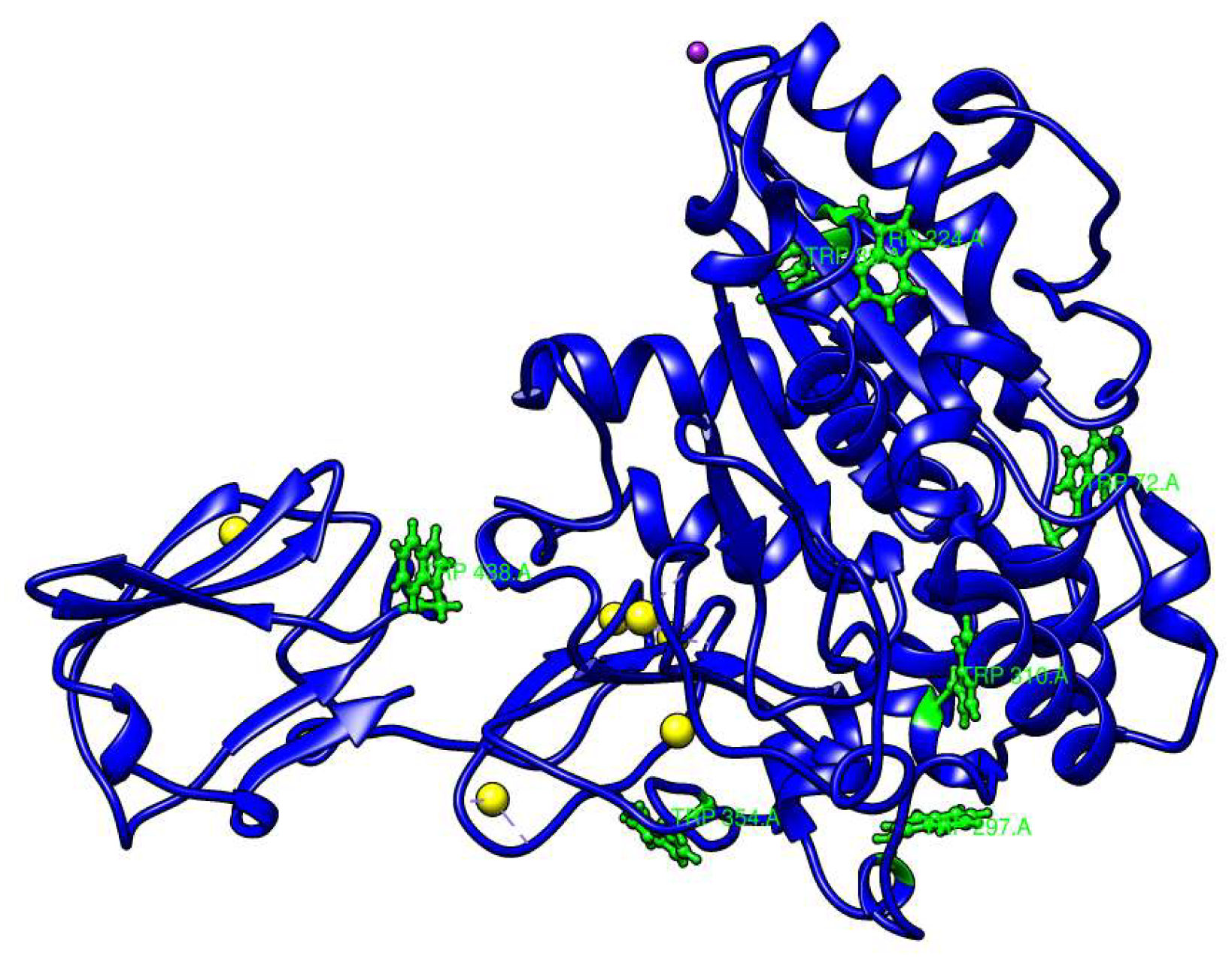
2.1. Calcium-Binding Site and RTX β-Roll Motif Repeat Structure of the AMS8 Lipase Predicted Model
2.2. Effect of Different Ca2+ Ion Concentrations on AMS8 Lipase Activity
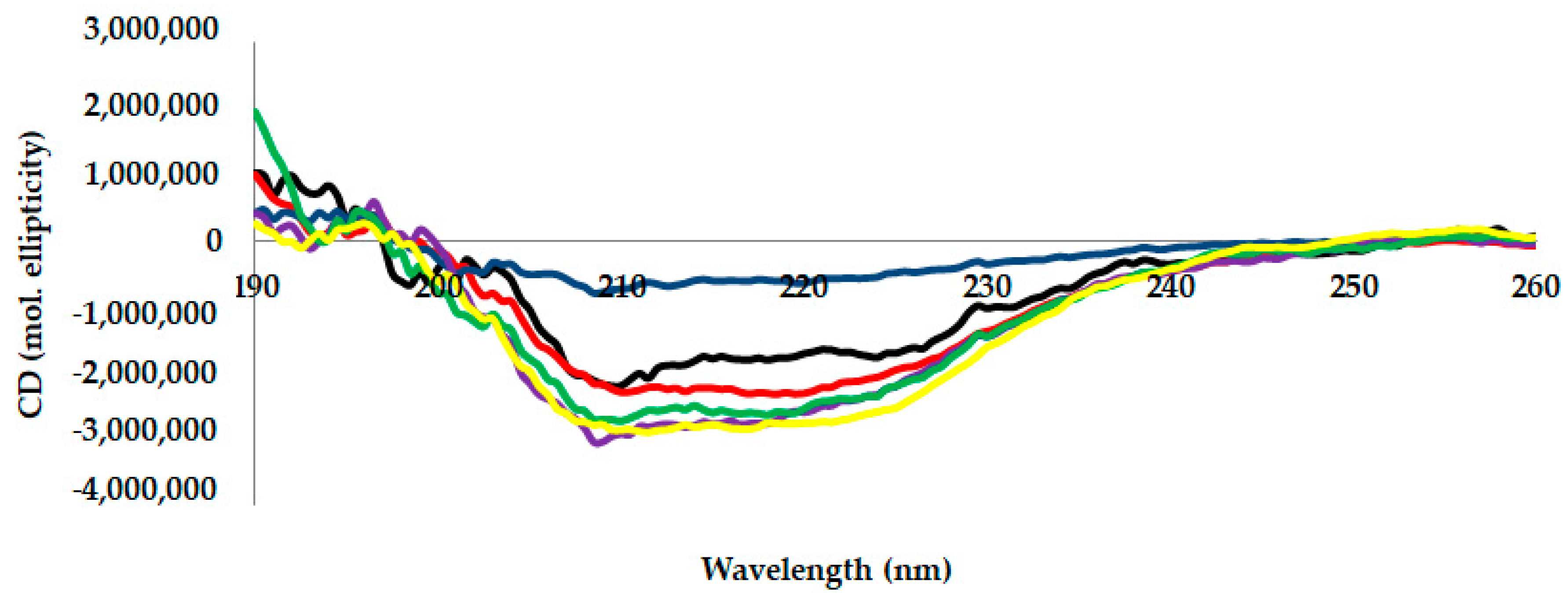
2.3. Circular Dichroism (CD) Analysis
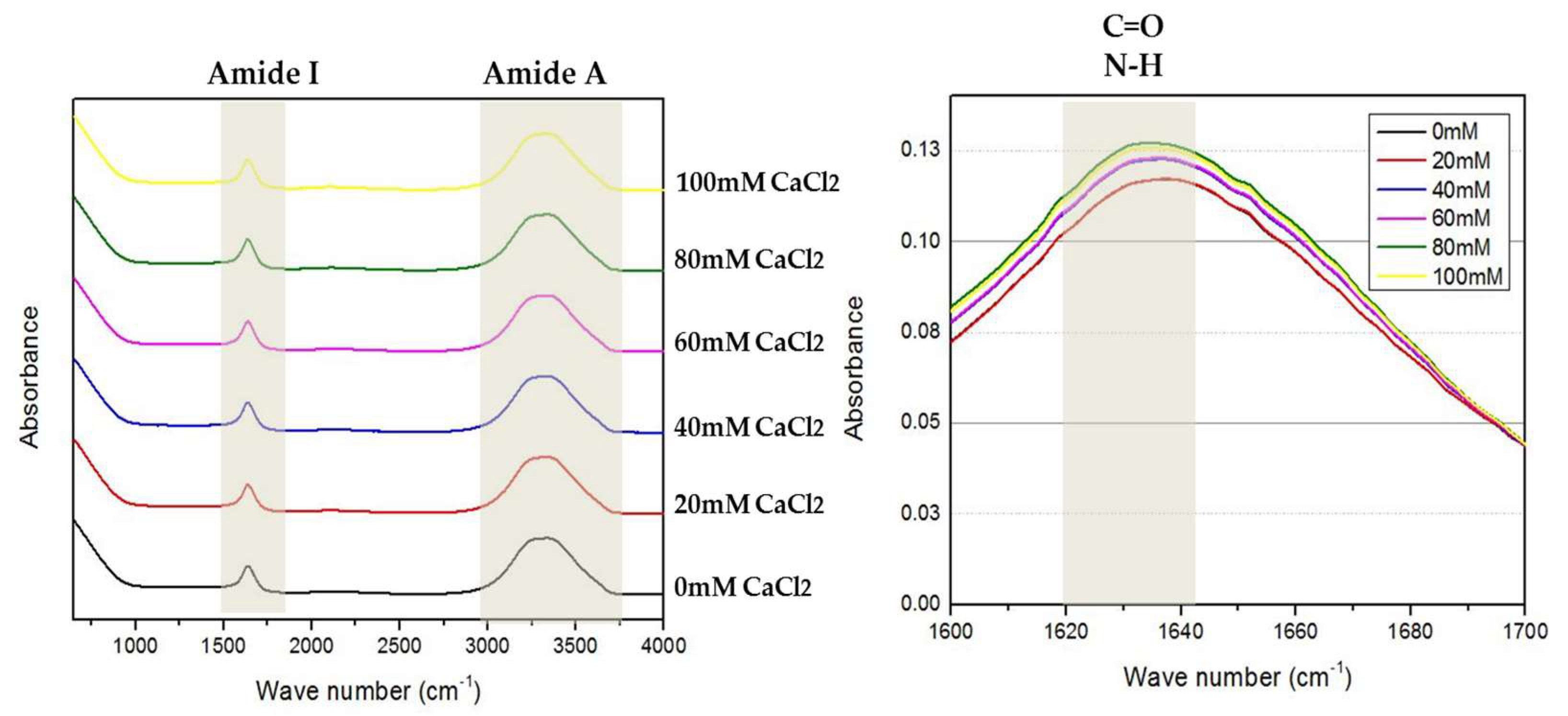
2.4. Fourier-Transform Infrared Spectroscopy (FTIR) Analysis
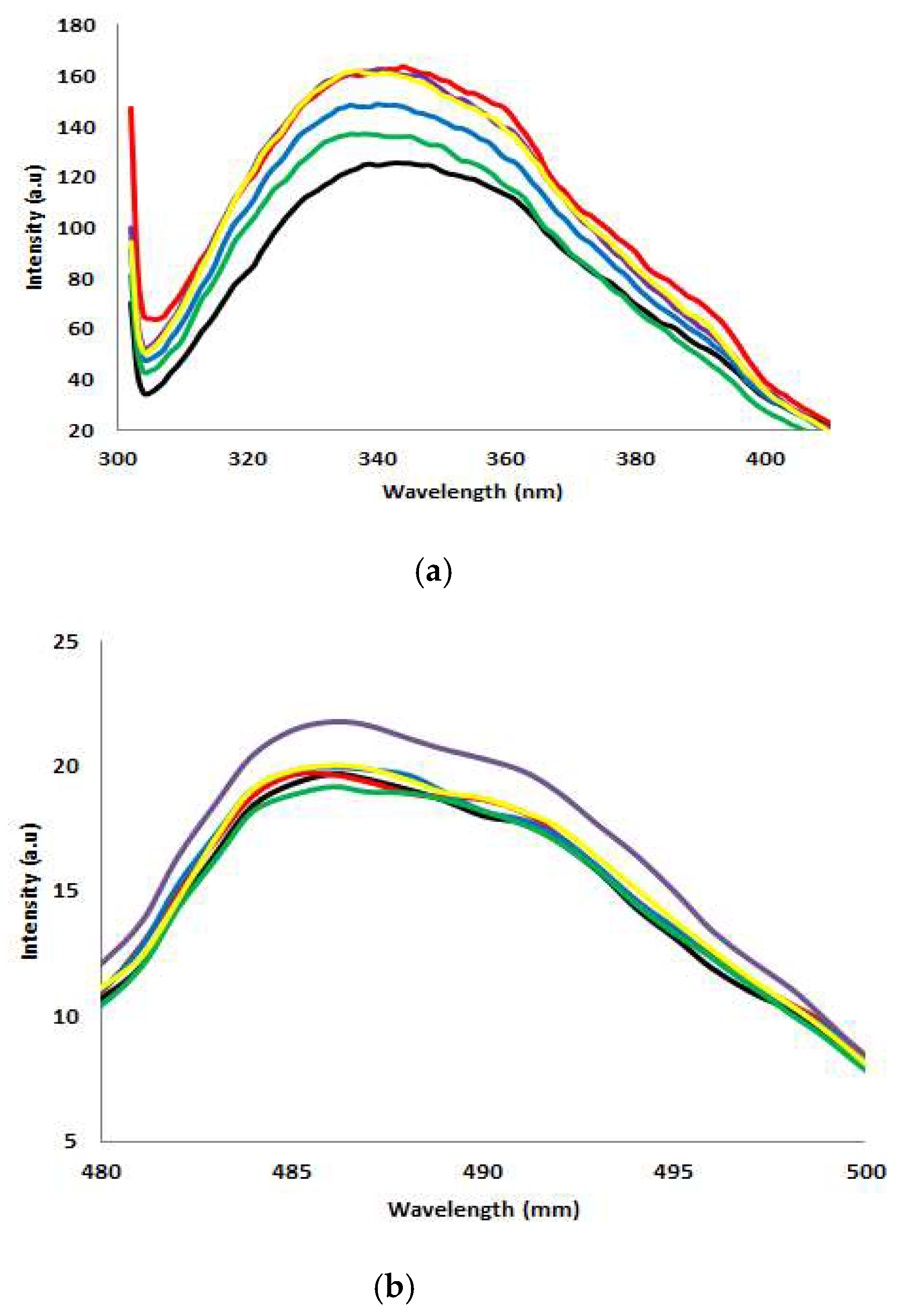
2.5. Fluorescence Spectroscopy Analysis
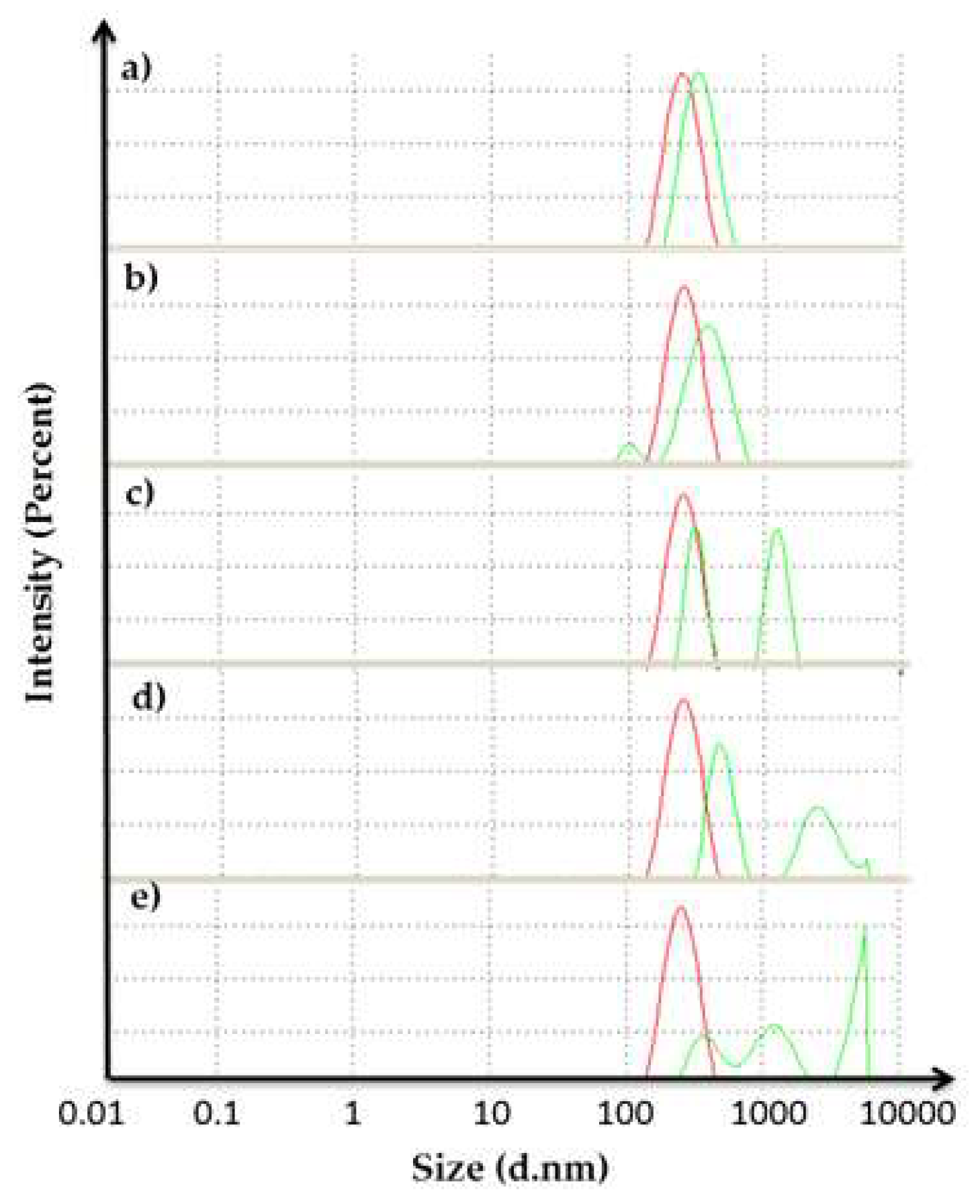
2.6. Dynamic Light Scattering (DLS) Analysis
2.7. Isothermal Titration Calorimetry (ITC) Analysis
3. Conclusions
4. Materials and Methods
4.1. Calcium-Binding Site of AMS8 Lipase Predicted Model
4.2. Expression, Solubilization and Refolding of AMS8 IBs with Different Ca2+ Ion Concentrations
4.3. Determination of Lipase Activity and Protein Content
4.4. Biophysical Characterizations
4.4.1. SDS-PAGE (Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis) Analysis
4.4.2. Circular Dichroism (CD) Spectra Analysis
4.4.3. Fourier-Transform Infrared (FTIR) Spectroscopy Analysis
4.4.4. Fluorescence Spectra Analysis
4.4.5. Dynamic Light Scattering (DLS) Analysis
4.4.6. Isothermal Titration Calorimetry (ITC) Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Linhartova, I.; Bumba, L.; Masin, J.; Basler, M.; Osicka, R.; Kamanova, J.; Prochazkova, K.; Adkins, I.; Hejnova-Holubova, J.; Sadilkova, L.; et al. RTX Proteins: A Highly Diverse Family Secreted by a Common Mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef]
- Sotomayor-Perez, A.C.; Ladant, D.; Chenal, A. Calcium-Induced Folding of Intrinsically Disordered Repeat-in-Toxin (RTX) Motifs via Changes of Protein Charges and Oligomerization States. J. Biol. Chem. 2011, 286, 16997–17004. [Google Scholar] [CrossRef] [PubMed]
- Chenal, A.; Guijarro, J.I.; Raynal, B.; Delepierre, M.; Ladant, D. RTX Calcium Binding Motifs Are Intrinsically Disordered in the Absence of Calcium: Implication for Protein Secretion. J. Biol. Chem. 2009, 284, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Lilie, H.; Haehnel, W.; Rudolph, R.; Baumann, U. Folding of a Synthetic Parallel β-Roll Protein. FEBS Lett. 2000, 470, 173–177. [Google Scholar] [CrossRef]
- Zhang, L.; Conway, J.F.; Thibodeau, P.H. Calcium-Induced Folding and Stabilization of the Pseudomonas Aeruginosa Alkaline Protease. J. Biol. Chem. 2012, 287, 4311–4322. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.S.M.; Ganasen, M.; Rahman, R.N.Z.R.A.; Chor, A.L.T.; Salleh, A.B.; Basri, M. Cold-Adapted RTX Lipase from Antarctic Pseudomonas sp. Strain AMS8: Isolation, Molecular Modeling and Heterologous Expression. Protein J. 2013, 32, 317–325. [Google Scholar] [CrossRef]
- Ali, M.S.M.; Fuzi, S.F.M.; Ganasen, M.; Rahman, R.N.Z.R.A.; Basri, M.; Salleh, A.B. Structural Adaptation of Cold-Active RTX Lipase from Pseudomonas sp. Strain AMS8 Revealed via Homology and Molecular Dynamics Simulation Approaches. BioMed Res. Int. 2013, 2013, 925373. [Google Scholar] [CrossRef]
- Yaacob, N.; Mohamad Ali, M.S.; Salleh, A.B.; Rahman, R.N.Z.R.A.; Leow, A.T.C. Toluene Promotes Lid 2 Interfacial Activation of Cold Active Solvent Tolerant Lipase from Pseudomonas Fluorescens Strain AMS8. J. Mol. Graph. Model. 2016, 68, 224–235. [Google Scholar] [CrossRef]
- Short, E.C.; Kurtz, H.J. Properties of the Hemolytic Activities of Escherichia coli. Infect. Immun. 1971, 3, 678–687. [Google Scholar]
- Stigler, J.; Ziegler, F.; Gieseke, A.; Gebhardt, J.C.M.; Rief, M. The Complex Folding Network of Single Calmodulin Molecules. Science 2011, 334, 512–516. [Google Scholar] [CrossRef]
- Ganasen, M.; Yaacob, N.; Rahman, R.N.Z.R.A.; Leow, A.T.C.; Basri, M.; Salleh, A.B.; Ali, M.S.M. Cold-Adapted Organic Solvent Tolerant Alkalophilic Family I.3 Lipase from an Antarctic Pseudomonas. Int. J. Biol. Macromol. 2016, 92, 1266–1276. [Google Scholar] [CrossRef] [PubMed]
- Abd. Jalil, F.; Raja Abd. Rahman, R.; Salleh, A.; Mohamad Ali, M. Optimization and in Silico Analysis of a Cold-Adapted Lipase from an Antarctic Pseudomonas sp. Strain AMS8 Reaction in Triton X-100 Reverse Micelles. Catalysts 2018, 8, 289. [Google Scholar] [CrossRef]
- Farfel, Z.; Friedman, E.; Hanski, E. The Invasive Adenylate Cyclase of Bordetella Pertussis. Intracellular Localization and Kinetics of Penetration into Various Cells. Biochem. J. 1987, 243, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Gangola, P.; RosenS, B.P. Maintenance of Intracellular Calcium in Escherichia coli. J. Biol. Chem. 1987, 262, 12570–12574. [Google Scholar]
- Kenny, B.; Haigh, R. Analysis of the haemolysin transport process through the secretion from Escherichia coli of PCM, CAT or β-galactosidase fused to the Hly C-terminal signal domain. Mol. Microbiol. 1991, 5, 2557–2568. [Google Scholar] [CrossRef]
- Koronakis, E.; Hughes, C.; Milisav, I.; Koronakis, V. Protein exporter function and in vitro ATPase activity are correlated in ABC-domain mutants of HlyB. Mol. Microbiol. 1995, 16, 87–96. [Google Scholar] [CrossRef]
- Felmlee, T.; Welch, R.A. Alterations of Amino Acid Repeats in the Escherichia Coli Hemolysin Affect Cytolytic Activity and Secretion. Proc. Natl. Acad. Sci. USA 1988, 85, 5269–5273. [Google Scholar] [CrossRef]
- Rhodes, C.R.; Gray, M.C.; Watson, J.M.; Muratore, T.L.; Kim, S.B.; Hewlett, E.L.; Grisham, C.M. Structural Consequences of Divalent Metal Binding by the Adenylyl Cyclase Toxin of Bordetella pertussis. Arch. Biochem. Biophys. 2001, 395, 169–176. [Google Scholar] [CrossRef]
- Rose, T.; Sebo, P.; Bellalou, J.; Ladant, D. Characterization of Multiple Calcium-Binding Sites and Calcium-Induced Conformational Changes. J. Biol. Chem. 1995, 44, 26370–26376. [Google Scholar] [CrossRef]
- Bindreither, D.; Lackner, P. Structural Diversity of Calcium Binding Sites. Gen. Physiol. Biophys. 2009, 28, 82–88. [Google Scholar]
- O’Brien, D.P.; Perez, A.C.S.; Karst, J.; Cannella, S.E.; Enguéné, V.Y.N.; Hessel, A.; Raoux-Barbot, D.; Voegele, A.; Subrini, O.; Davi, M.; et al. Calcium-Dependent Disorder-to-Order Transitions Are Central to the Secretion and Folding of the CyaA Toxin of Bordetella Pertussis, the Causative Agent of Whooping Cough. Toxicon 2018, 149, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Isik, M.; Altas, L.; Ozcan, S.; Simsek, I.; Agdag, O.N.; Alas, A. Effect of Urea Concentration on Microbial Ca Precipitation. J. Ind. Eng. Chem. 2012, 18, 1908–1911. [Google Scholar] [CrossRef]
- Alquati, C.; De Gioia, L.; Santarossa, G.; Alberghina, L.; Fantucci, P.; Lotti, M. The Cold-Active Lipase of Pseudomonas Fragi: Heterologous Expression, Biochemical Characterization and Molecular Modeling. Eur. J. Biochem. 2002, 269, 3321–3328. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Nakatani, Y.; Hisatomi, O. Accuracy of Protein Size Estimates Based on Light Scattering Measurements. Open J. Biophys. 2014, 4, 83–91. [Google Scholar] [CrossRef]
- Bauche, C.; Chenal, A.; Knapp, O.; Bodenreider, C.; Benz, R.; Chaffotte, A.; Ladant, D. Structural and Functional Characterization of an Essential RTX Subdomain of Bordetella Pertussis Adenylate Cyclase Toxin. J. Biol. Chem. 2006, 281, 16914–16926. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, W. FTIR Analysis of Protein Structure. Course Man. Chem. 2009, 455, 1–8. [Google Scholar]
- Fabian, H.; Vogel, H.J. Fourier Transform Infrared Spectroscopy of Calcium-Binding Proteins. In Calcium-Binding Protein Protocols; Humana Press: Totowa, NJ, USA, 2002; Volume 173, pp. 57–74. [Google Scholar] [CrossRef]
- Alhazmi, H.A. FT-IR Spectroscopy for the Identification of Binding Sites and Measurements of the Binding Interactions of Important Metal Ions with Bovine Serum Albumin. Sci. Pharm. 2019, 87, 1–5. [Google Scholar] [CrossRef]
- Kwaambwa, H.M.; Maikokera, R. A Fluorescence Spectroscopic Study of a Coagulating Protein Extracted from Moringa Oleifera Seeds. Colloids Surf. B Biointerfaces 2007, 60, 213–220. [Google Scholar] [CrossRef]
- Das, S.; Pal, U.; Chandra Maiti, N. Metal Ions Provide Structural Stability and Compactness to Tetrameric Purothionin. RSC Adv. 2016, 6, 90690–90700. [Google Scholar] [CrossRef]
- Singh, N.; Kumar, R.; Jagannadham, M.V.; Kayastha, A.M. Evidence for a Molten Globule State in Cicer α-Galactosidase Induced by PH, Temperature, and Guanidine Hydrochloride. Appl. Biochem. Biotechnol. 2013, 169, 2315–2325. [Google Scholar] [CrossRef]
- Falconer, R.J. Applications of Isothermal Titration Calorimetry—The Research and Technical Developments from 2011 to 2015: Review of Isothermal Titration Calorimetry from 2011 to 2015. J. Mol. Recognit. 2016, 29, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, A.; Stein, A.; Jahn, R.; Fasshauer, D. The Ca2+ affinity of synaptotagmin 1 is markedly increased by a specific interaction of its C2B domain with phosphatidylinositol 4,5-bisphosphate. J. Biol. Chem. 2009, 284, 25749–25760. [Google Scholar] [CrossRef] [PubMed]
- Krieger, E.; Koraimann, G.; Vriend, G. Increasing the Precision of Comparative Models with YASARA NOVA-a Self-Parameterizing Force Field. Proteins 2002, 47, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.Y.; Rhee, J.S. A Simple and Rapid Colorimetric Method for Determination of Free Fatty Acids for Lipase Assay. J. Am. Oil Chem. 1986, 63, 89–92. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Yang, J.T.; Wu, C.-S.C.; Martinez, H.M. [11] Calculation of Protein Conformation from Circular Dichroism. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1986; Volume 130, pp. 208–269. [Google Scholar] [CrossRef]
- Hall, H. Malvern Zetasizer Nano Basic Operating Instructions; Iowa State University: Ames, IA, USA, 2015. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CaCl2 Concentration (mM) | Lipase Activity (U/mL) |
|---|---|
| 0 | 26.40 ± 0.74 |
| 5 | 137.99 ± 7.70 |
| 10 | 157.08 ± 1.81 |
| 20 | 160.00 ± 3.69 |
| 30 | 169.23 ± 2.92 |
| 40 | 173.88 ± 2.98 |
| 50 | 183.67 ± 0.93 |
| 60 | 198.48 ± 1.58 |
| 70 | 218.23 ± 0.25 |
| 80 | 262.49 ± 3.91 |
| 90 | 255.18 ± 1.74 |
| 100 | 226.69 ± 1.32 |
| CaCl2 Concentrations (mM) | Percentage (%) | |||
|---|---|---|---|---|
| Beta | Helix | Turn | Random | |
| 0 | 7.7 | 25.3 | 30.7 | 36.5 |
| 20 | 12.9 | 29.7 | 22.1 | 35.2 |
| 40 | 22.1 | 19.4 | 21.7 | 36.8 |
| 60 | 24.3 | 22.3 | 21.2 | 36.3 |
| 80 | 26.7 | 21.2 | 19.9 | 32.2 |
| 100 | 19.6 | 24.7 | 20.3 | 35.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, N.S.M.; Salleh, A.B.; Rahman, R.N.Z.R.A.; Leow, T.C.; Ali, M.S.M. Calcium-Induced Activity and Folding of a Repeat in Toxin Lipase from Antarctic Pseudomonas fluorescens Strain AMS8. Toxins 2020, 12, 27. https://doi.org/10.3390/toxins12010027
Ali NSM, Salleh AB, Rahman RNZRA, Leow TC, Ali MSM. Calcium-Induced Activity and Folding of a Repeat in Toxin Lipase from Antarctic Pseudomonas fluorescens Strain AMS8. Toxins. 2020; 12(1):27. https://doi.org/10.3390/toxins12010027
Chicago/Turabian StyleAli, Nur Shidaa Mohd, Abu Bakar Salleh, Raja Noor Zaliha Raja Abd Rahman, Thean Chor Leow, and Mohd Shukuri Mohamad Ali. 2020. "Calcium-Induced Activity and Folding of a Repeat in Toxin Lipase from Antarctic Pseudomonas fluorescens Strain AMS8" Toxins 12, no. 1: 27. https://doi.org/10.3390/toxins12010027
APA StyleAli, N. S. M., Salleh, A. B., Rahman, R. N. Z. R. A., Leow, T. C., & Ali, M. S. M. (2020). Calcium-Induced Activity and Folding of a Repeat in Toxin Lipase from Antarctic Pseudomonas fluorescens Strain AMS8. Toxins, 12(1), 27. https://doi.org/10.3390/toxins12010027