Granulocyte Colony-Stimulating Factor Does Not Influence Clostridium Perfringens α-Toxin-Induced Myonecrosis in Mice

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
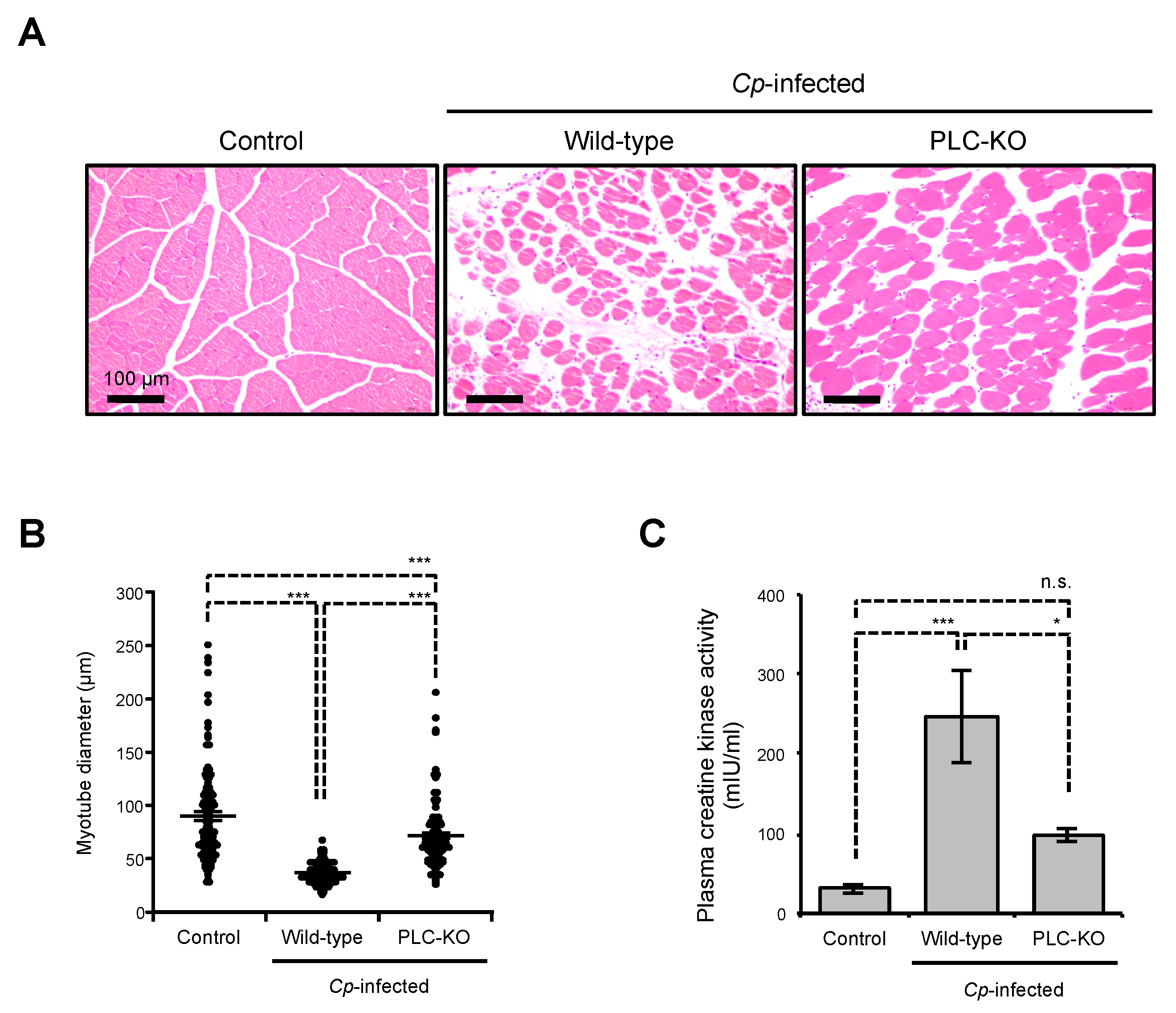
2.1. C. perfringens Induces Myonecrosis in a Toxin-Dependent Manner
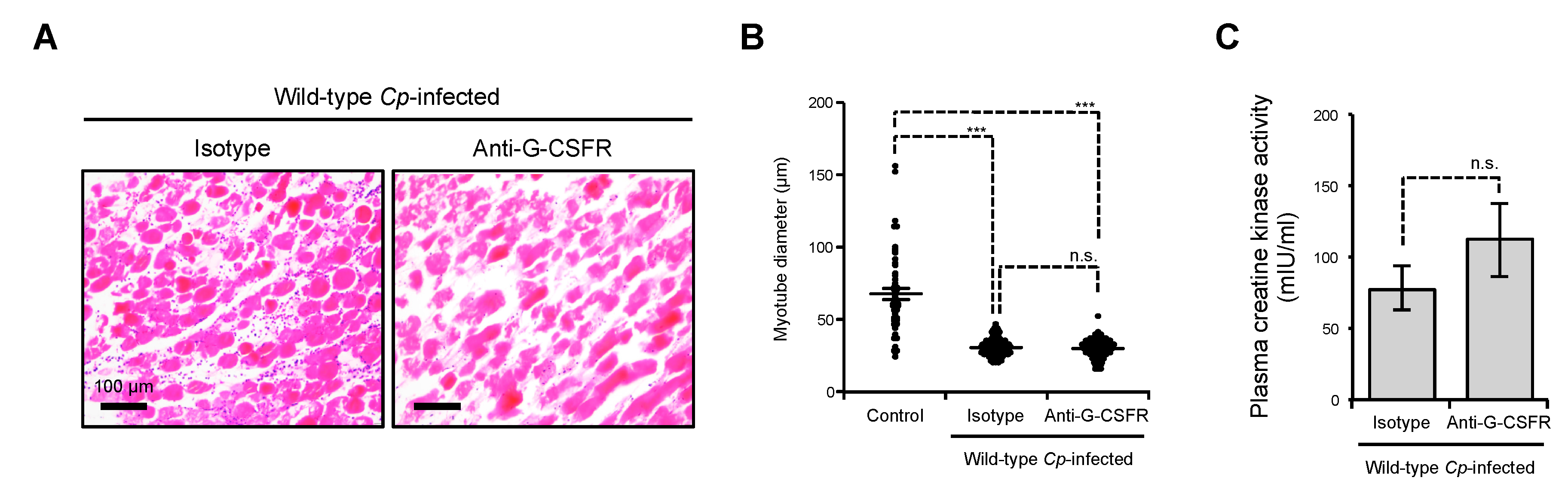
2.2. Inhibition of G-CSFR Does Not Affect C. perfringens α-Toxin-Induced Myonecrosis In Vivo
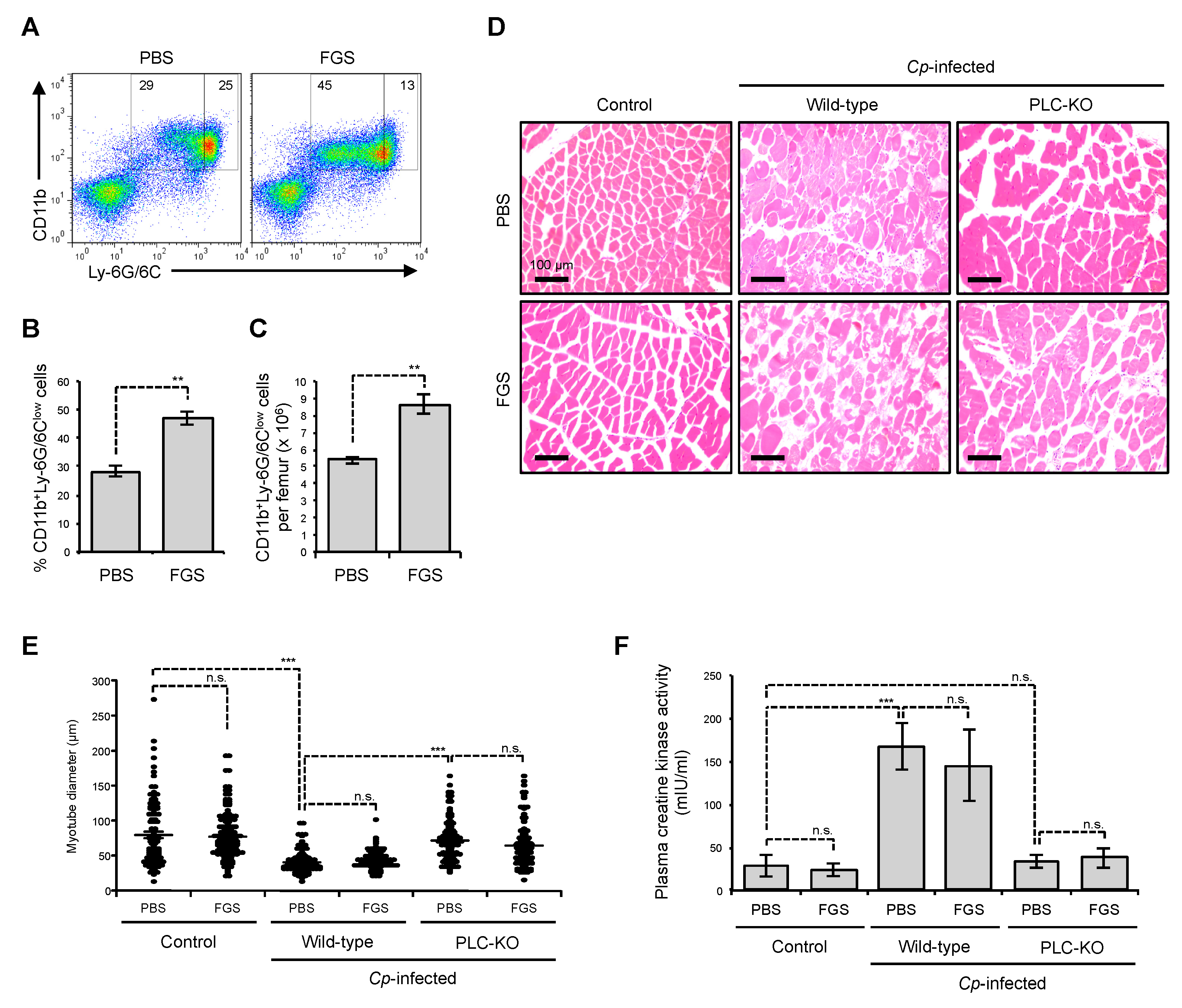
2.3. Filgrastim Has no Protective Effect on Skeletal Muscle During C. perfringens α-Toxin-Mediated Myonecrosis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Mice
5.2. Reagents and Strains
5.3. Flow Cytometry Analysis
5.4. Bacterial Culture and Infection
5.5. Myotube Morphology Analysis
5.6. Administration of G-CSF and Inhibition of G-CSFR In Vivo
5.7. Measurement of Plasma Creatine Kinase
5.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Songer, J.G. Clostridial enteric diseases of domestic animals. Clin. Microbiol. Rev. 1996, 9, 216–234. [Google Scholar] [CrossRef] [PubMed]
- Petit, L.; Gibert, M.; Popoff, M.R. Clostridium perfringens: Toxinotype and genotype. Trends Microbiol. 1999, 7, 104–110. [Google Scholar] [CrossRef]
- Bryant, A.E. Biology and pathogenesis of thrombosis and procoagulant activity in invasive infections caused by group A streptococci and Clostridium perfringens. Clin. Microbiol. Rev. 2003, 16, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, J.; Nagahama, M.; Oda, M. Clostridium perfringens alpha-toxin: Characterization and mode of action. J. Biochem. 2004, 136, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Rood, J.I. Virulence genes of Clostridium perfringens. Annu. Rev. Microbiol. 1998, 52, 333–360. [Google Scholar] [CrossRef]
- Verherstraeten, S.; Goossens, E.; Valgaeren, B.; Pardon, B.; Timbermont, L.; Haesebrouck, F.; Ducatelle, R.; Deprez, P.; Wade, K.R.; Tweten, R.; et al. Perfringolysin O: The Underrated Clostridium perfringens Toxin? Toxins 2015, 7, 1702–1721. [Google Scholar] [CrossRef]
- Ellemor, D.M.; Baird, R.N.; Awad, M.M.; Boyd, R.L.; Rood, J.I.; Emmins, J.J. Use of genetically manipulated strains of Clostridium perfringens reveals that both alpha-toxin and theta-toxin are required for vascular leukostasis to occur in experimental gas gangrene. Infect. Immun. 1999, 67, 4902–4907. [Google Scholar]
- Stevens, D.L.; Mitten, J.; Henry, C. Effects of alpha and theta toxins from Clostridium perfringens on human polymorphonuclear leukocytes. J. Infect. Dis. 1987, 156, 324–333. [Google Scholar] [CrossRef]
- Stevens, D.L.; Bryant, A.E. Role of theta toxin, a sulfhydryl-activated cytolysin, in the pathogenesis of clostridial gas gangrene. Clin. Infect. Dis. 1993, 16, 195–199. [Google Scholar] [CrossRef]
- O’Brien, D.K.; Melville, S.B. Effects of Clostridium perfringens alpha-toxin (PLC) and perfringolysin O (PFO) on cytotoxicity to macrophages, on escape from the phagosomes of macrophages, and on persistence of C. perfringens in host tissues. Infect. Immun. 2004, 72, 5204–5215. [Google Scholar] [CrossRef]
- Bryant, A.E.; Bayer, C.R.; Aldape, M.J.; Wallace, R.J.; Titball, R.W.; Stevens, D.L. Clostridium perfringens phospholipase C-induced platelet/leukocyte interactions impede neutrophil diapedesis. J. Med. Microbiol. 2006, 55, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Ochi, S.; Miyawaki, T.; Matsuda, H.; Oda, M.; Nagahama, M.; Sakurai, J. Clostridium perfringens alpha-toxin induces rabbit neutrophil adhesion. Microbiology 2002, 148, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Bryant, A.E.; Chen, R.Y.; Nagata, Y.; Wang, Y.; Lee, C.H.; Finegold, S.; Guth, P.H.; Stevens, D.L. Clostridial gas gangrene. II. Phospholipase C-induced activation of platelet gpIIbIIIa mediates vascular occlusion and myonecrosis in Clostridium perfringens gas gangrene. J. Infect. Dis. 2000, 182, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Stevens, D.L.; Tweten, R.K.; Awad, M.M.; Rood, J.I.; Bryant, A.E. Clostridial gas gangrene: Evidence that alpha and theta toxins differentially modulate the immune response and induce acute tissue necrosis. J. Infect. Dis. 1997, 176, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Bahl, H.; Dürre, P. Clostridia: Biotechnology and Medical Applications; Wiley-VCH: Weinheim, Germany; Chichester, UK, 2001; p. xii. 279p. [Google Scholar]
- Takehara, M.; Takagishi, T.; Seike, S.; Ohtani, K.; Kobayashi, K.; Miyamoto, K.; Shimizu, T.; Nagahama, M. Clostridium perfringens α-toxin impairs innate immunity via inhibition of neutrophil differentiation. Sci. Rep. 2016, 6, 28192. [Google Scholar] [CrossRef]
- Takehara, M.; Takagishi, T.; Seike, S.; Oishi, K.; Fujihara, Y.; Miyamoto, K.; Kobayashi, K.; Nagahama, M. Clostridium perfringens α-toxin impairs lipid raft integrity in neutrophils. Biol. Pharm. Bull. 2016, 39, 1694–1700. [Google Scholar] [CrossRef]
- Shimizu, T.; Ohtani, K.; Hirakawa, H.; Ohshima, K.; Yamashita, A.; Shiba, T.; Ogasawara, N.; Hattori, M.; Kuhara, S.; Hayashi, H. Complete genome sequence of Clostridium perfringens, an anaerobic flesh-eater. Proc. Natl. Acad. Sci. USA 2002, 99, 996–1001. [Google Scholar] [CrossRef]
- Low, L.Y.; Harrison, P.F.; Gould, J.; Powell, D.R.; Choo, J.M.; Forster, S.C.; Chapman, R.; Gearing, L.J.; Cheung, J.K.; Hertzog, P.; et al. Concurrent Host-Pathogen Transcriptional Responses in a Clostridium perfringens Murine Myonecrosis Infection. MBio 2018, 9, e00473-18. [Google Scholar] [CrossRef]
- Roberts, A.W. G-CSF: A key regulator of neutrophil production, but that’s not all! Growth Factors 2005, 23, 33–41. [Google Scholar] [CrossRef]
- Piscaglia, A.C.; Shupe, T.D.; Oh, S.H.; Gasbarrini, A.; Petersen, B.E. Granulocyte-colony stimulating factor promotes liver repair and induces oval cell migration and proliferation in rats. Gastroenterology 2007, 133, 619–631. [Google Scholar] [CrossRef]
- Wright, C.R.; Brown, E.L.; Della-Gatta, P.A.; Ward, A.C.; Lynch, G.S.; Russell, A.P. G-CSF does not influence C2C12 myogenesis despite receptor expression in healthy and dystrophic skeletal muscle. Front. Physiol. 2014, 5, 170. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shimoji, K.; Yuasa, S.; Onizuka, T.; Hattori, F.; Tanaka, T.; Hara, M.; Ohno, Y.; Chen, H.; Egasgira, T.; Seki, T.; et al. G-CSF promotes the proliferation of developing cardiomyocytes in vivo and in derivation from ESCs and iPSCs. Cell Stem Cell 2010, 6, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Yata, K.; Matchett, G.A.; Tsubokawa, T.; Tang, J.; Kanamaru, K.; Zhang, J.H. Granulocyte-colony stimulating factor inhibits apoptotic neuron loss after neonatal hypoxia-ischemia in rats. Brain Res. 2007, 1145, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Yuasa, S.; Shimoji, K.; Onizuka, T.; Hayashiji, N.; Ohno, Y.; Arai, T.; Hattori, F.; Kaneda, R.; Kimura, K.; et al. G-CSF influences mouse skeletal muscle development and regeneration by stimulating myoblast proliferation. J. Exp. Med. 2011, 208, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Komine-Kobayashi, M.; Zhang, N.; Liu, M.; Tanaka, R.; Hara, H.; Osaka, A.; Mochizuki, H.; Mizuno, Y.; Urabe, T. Neuroprotective effect of recombinant human granulocyte colony-stimulating factor in transient focal ischemia of mice. J. Cereb. Blood Flow Metab. 2006, 26, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Pitzer, C.; Kruger, C.; Plaas, C.; Kirsch, F.; Dittgen, T.; Muller, R.; Laage, R.; Kastner, S.; Suess, S.; Spoelgen, R.; et al. Granulocyte-colony stimulating factor improves outcome in a mouse model of amyotrophic lateral sclerosis. Brain 2008, 131, 3335–3347. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Takemura, G.; Li, Y.; Miyata, S.; Esaki, M.; Okada, H.; Kanamori, H.; Ogino, A.; Maruyama, R.; Nakagawa, M.; et al. Granulocyte colony-stimulating factor improves left ventricular function of doxorubicin-induced cardiomyopathy. Lab. Investig. 2007, 87, 440–455. [Google Scholar] [CrossRef] [PubMed]
- Solaroglu, I.; Cahill, J.; Jadhav, V.; Zhang, J.H. A novel neuroprotectant granulocyte-colony stimulating factor. Stroke 2006, 37, 1123–1128. [Google Scholar] [CrossRef]
- Stratos, I.; Rotter, R.; Eipel, C.; Mittlmeier, T.; Vollmar, B. Granulocyte-colony stimulating factor enhances muscle proliferation and strength following skeletal muscle injury in rats. J. Appl. Physiol. 2007, 103, 1857–1863. [Google Scholar] [CrossRef]
- Monturiol-Gross, L.; Flores-Diaz, M.; Araya-Castillo, C.; Pineda-Padilla, M.J.; Clark, G.C.; Titball, R.W.; Alape-Giron, A. Reactive oxygen species and the MEK/ERK pathway are involved in the toxicity of clostridium perfringens alpha-toxin, a prototype bacterial phospholipase C. J. Infect. Dis. 2012, 206, 1218–1226. [Google Scholar] [CrossRef]
- Takehara, M.; Seike, S.; Sonobe, Y.; Bandou, H.; Yokoyama, S.; Takagishi, T.; Miyamoto, K.; Kobayashi, K.; Nagahama, M. Clostridium perfringens alpha-toxin impairs granulocyte colony-stimulating factor receptor-mediated granulocyte production while triggering septic shock. Commun. Biol. 2019, 2, 45. [Google Scholar] [CrossRef] [PubMed]
- Files, D.C.; Liu, C.; Pereyra, A.; Wang, Z.M.; Aggarwal, N.R.; D’Alessio, F.R.; Garibaldi, B.T.; Mock, J.R.; Singer, B.D.; Feng, X.; et al. Therapeutic exercise attenuates neutrophilic lung injury and skeletal muscle wasting. Sci. Transl. Med. 2015, 7, 278ra232. [Google Scholar] [CrossRef] [PubMed]
- Frampton, J.E.; Lee, C.R.; Faulds, D.F. A review of its pharmacological properties and therapeutic efficacy in neutropenia. Drugs 1994, 48, 731–760. [Google Scholar] [CrossRef] [PubMed]
- Takehara, M.; Seike, S.; Takagishi, T.; Kobayashi, K.; Nagahama, M. Peptidoglycan accelerates granulopoiesis through a TLR2- and MyD88-dependent pathway. Biochem. Biophys. Res. Commun. 2017, 487, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takehara, M.; Sonobe, Y.; Bandou, H.; Kobayashi, K.; Nagahama, M. Granulocyte Colony-Stimulating Factor Does Not Influence Clostridium Perfringens α-Toxin-Induced Myonecrosis in Mice. Toxins 2019, 11, 509. https://doi.org/10.3390/toxins11090509
Takehara M, Sonobe Y, Bandou H, Kobayashi K, Nagahama M. Granulocyte Colony-Stimulating Factor Does Not Influence Clostridium Perfringens α-Toxin-Induced Myonecrosis in Mice. Toxins. 2019; 11(9):509. https://doi.org/10.3390/toxins11090509
Chicago/Turabian StyleTakehara, Masaya, Yuuta Sonobe, Hiroto Bandou, Keiko Kobayashi, and Masahiro Nagahama. 2019. "Granulocyte Colony-Stimulating Factor Does Not Influence Clostridium Perfringens α-Toxin-Induced Myonecrosis in Mice" Toxins 11, no. 9: 509. https://doi.org/10.3390/toxins11090509
APA StyleTakehara, M., Sonobe, Y., Bandou, H., Kobayashi, K., & Nagahama, M. (2019). Granulocyte Colony-Stimulating Factor Does Not Influence Clostridium Perfringens α-Toxin-Induced Myonecrosis in Mice. Toxins, 11(9), 509. https://doi.org/10.3390/toxins11090509