Virulence Characteristics and Antimicrobial Resistance Profiles of Shiga Toxin-Producing Escherichia coli Isolates from Humans in South Africa: 2006–2013

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. STEC Serotypes and Their Frequency in Human Outbreaks
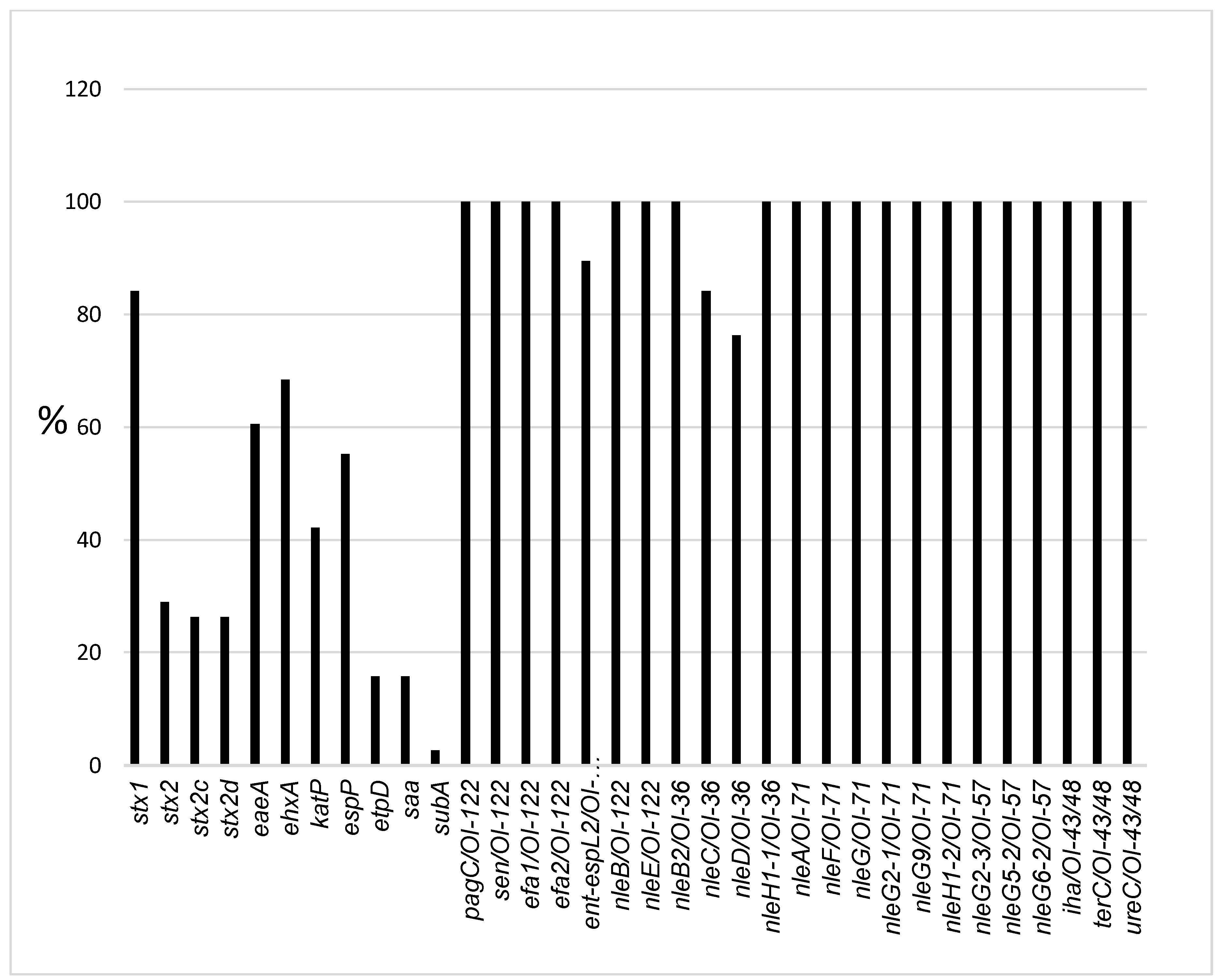
2.2. Virulence-Associated Gene Distribution
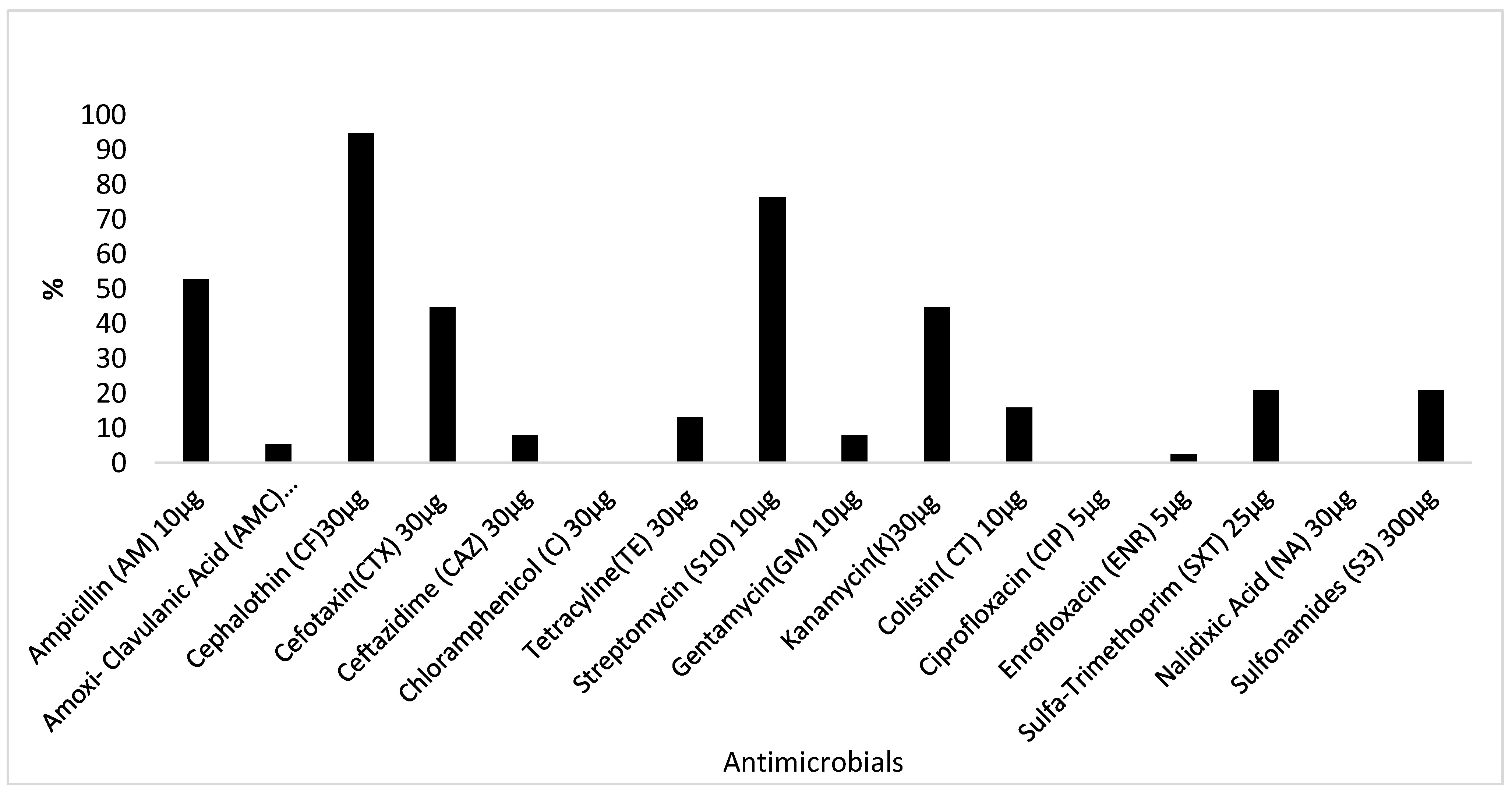
2.3. Antimicrobial Resistance Profiling
2.4. Pulsed-Field Gel Electrophoresis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. STEC Strains and Culture Conditions
5.2. STEC Cultures
5.3. STEC PCR Serotyping (O:H)
5.4. Virulotyping
5.5. Antimicrobial Susceptibility Testing
5.6. Pulsed-Field Gel Electrophoresis
5.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Kaper, J.B.; Karmali, M.A. The continuing evolution of a bacterial pathogen. Proc. Natl. Acad. Sci. USA 2008, 105, 4535–4536. [Google Scholar] [CrossRef]
- Majowicz, S.E.; Scallan, E.; Jones-Bitton, A.; Sargeant, J.M.; Stapleton, J.; Angulo, F.J.; Yeung, D.H.; Kirk, M.D. Global incidence of human Shiga toxin–producing Escherichia coli infections and deaths: A systematic review and knowledge synthesis. Foodborne Pathog. Dis. 2014, 11, 447–455. [Google Scholar] [CrossRef]
- Hussein, H.S. Prevalence and pathogenicity of Shiga toxin-producing Escherichia coli in beef cattle and their products. J. Anim. Sci. 2007, 85, E63–E72. [Google Scholar] [CrossRef]
- Hussein, H.S.; Sakuma, T. Invited review: prevalence of Shiga toxin-producing Escherichia coli in dairy cattle and their products. J. Dairy Sci. 2005, 88, 450–465. [Google Scholar] [CrossRef]
- Berger, C.N.; Sodha, S.V.; Shaw, R.K.; Griffin, P.M.; Pink, D.; Hand, P.; Frankel, G. Fresh fruit and vegetables as vehicles for the transmission of human pathogens. Environ. Microbiol. 2010, 12, 2385–2397. [Google Scholar] [CrossRef]
- Beutin, L.; Fach, P. Detection of Shiga toxin-producing Escherichia coli from nonhuman sources and strain typing. In Enterohemorrhagic Escherichia coli and Other Shiga Toxin-Producing, E. coli; American Society of Microbiology: Washington, DC, USA, 2015; pp. 299–319. [Google Scholar]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on VTEC-Seropathotype and Scientific Criteria Regarding Pathogenicity Assessment. EFSA J. 2013, 11, 3138. [Google Scholar]
- Bettelheim, K.A. The non-O157 Shiga-toxigenic (verocytotoxigenic) Escherichia coli; under-rated pathogens. Crit. Rev. Microbiol. 2007, 33, 67–87. [Google Scholar] [CrossRef]
- Johnson, K.E.; Thorpe, C.M.; Sears, C.L. The emerging clinical importance of non-O157 Shiga toxin—Producing Escherichia coli. Clin. Infect. Dis. 2006, 43, 1587–1595. [Google Scholar]
- Scallan, E.; Griffin, P.M.; Angulo, F.J.; Tauxe, R.V.; Hoekstra, R.M. Foodborne illness acquired in the United States—Unspecified agents. Emerg. Infect. Dis. 2011, 17, 16–22. [Google Scholar] [CrossRef]
- Karmali, M.A.; Mascarenhas, M.; Shen, S.; Ziebell, K.; Johnson, S.; Reid-Smith, R.; Isaac-Renton, J.; Clark, C.; Rahn, K.; Kaper, J.B. Association of genomic O island 122 of Escherichia coli EDL 933 with verocytotoxin-producing Escherichia coli seropathotypes that are linked to epidemic and/or serious disease. J. Clin. Microbiol. 2003, 41, 4930–4940. [Google Scholar] [CrossRef]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Piérard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef]
- Lacher, D.W.; Gangiredla, J.; Jackson, S.A.; Elkins, C.A.; Feng, P.C.H. A novel microarray design for molecular serotyping of Shiga toxin-producing Escherichia coli isolated from fresh produce. Appl. Environ. Microbiol. 2014, 80, 4677–4682. [Google Scholar] [CrossRef]
- Bai, X.; Fu, S.; Zhang, J.; Fan, R.; Xu, Y.; Sun, H.; He, X.; Xu, J.; Xiong, Y. Identification and pathogenomic analysis of an Escherichia coli strain producing a novel Shiga toxin 2 subtype. Sci. Rep. 2018. [Google Scholar] [CrossRef]
- Tzipori, S.; Gunzer, F.; Donnenberg, M.S.; de Montigny, L.; Kaper, J.B.; Donohue-Rolfe, A. The role of the eaeA gene in diarrhea and neurological complications in a gnotobiotic piglet model of enterohemorrhagic Escherichia coli infection. Infect. Immun. 1995, 63, 3621–3627. [Google Scholar]
- McDaniel, T.K.; Jarvis, K.G.; Donnenberg, M.S.; Kaper, J.B. A genetic locus of enterocyte effacement conserved among diverse enterobacterial pathogens. Proc. Natl. Acad. Sci. USA 1995, 92, 1664–1668. [Google Scholar] [CrossRef]
- Donnenberg, M.S.; Tacket, C.O.; James, S.P.; Losonsky, G.; Nataro, J.P.; Wasserman, S.S.; Kaper, J.B.; Levine, M.M. Role of the eaeA gene in experimental enteropathogenic Escherichia coli infection. J. Clin. Investig. 1993, 92, 1412–1417. [Google Scholar] [CrossRef]
- Schmidt, H.; Beutin, L.; Karch, H. Molecular analysis of the plasmid-encoded hemolysin of Escherichia coli O157:H7 strain EDL 933. Infect. Immun. 1995, 63, 1055–1061. [Google Scholar]
- Brunder, W.; Schmidt, H.; Karch, H. KatP, a novel catalase-peroxidase encoded by the large plasmid of enterohaemorrhagic Escherichia coli O157:H7. Microbiology 1996, 142, 3305–3315. [Google Scholar] [CrossRef]
- Brunder, W.; Schmidt, H.; Karch, H. EspP, a novel extracellular serine protease of enterohaemorrhagic Escherichia coli O157:H7 cleaves human coagulation factor V. Mol. Microbiol. 1997, 24, 767–778. [Google Scholar] [CrossRef]
- Schmidt, H.; Henkel, B.; Karch, H. A gene cluster closely related to type II secretion pathway operons of gram-negative bacteria is located on the large plasmid of enterohemorrhagic Escherichia coli O157 strains. FEMS Microbiol. Lett. 1997, 148, 265–272. [Google Scholar] [CrossRef]
- Paton, A.W.; Srimanote, P.; Talbot, U.M.; Wang, H.; Paton, J.C. A new family of potent AB5 cytotoxins produced by Shiga toxigenic Escherichia coli. J. Exp. Med. 2004, 200, 35–46. [Google Scholar] [CrossRef]
- Paton, A.W.; Srimanote, P.; Woodrow, M.C.; Paton, J.C. Characterization of Saa, a novel autoagglutinating adhesin produced by locus of enterocyte effacement-negative Shiga-Toxigenic Escherichia coli Strains that are virulent for humans. Infect. Immun. 2001, 69, 6999–7009. [Google Scholar] [CrossRef]
- Brunder, W.; Schmidt, H.; Frosch, M.; Karch, H. The large plasmids of Shiga-toxin-producing Escherichia coli (STEC) are highly variable genetic elements. Microbiology 1999, 145, 1005–1014. [Google Scholar] [CrossRef]
- Paton, A.W.; Paton, J.C. Escherichia coli subtilase cytotoxin. Toxins 2010, 2, 215–228. [Google Scholar] [CrossRef]
- Schmidt, H.; Hensel, M. Pathogenicity islands in bacterial pathogenesis. Clin. Microbiol. Rev. 2004, 17, 14–56. [Google Scholar] [CrossRef]
- Konczy, P.; Ziebell, K.; Mascarenhas, M.; Choi, A.; Michaud, C.; Kropinski, A.M.; Whittam, T.S.; Wickham, M.; Finlay, B.; Karmali, M.A. Genomic O island 122, locus for enterocyte effacement, and the evolution of virulent verocytotoxin-producing Escherichia coli. J. Bacteriol. 2008, 190, 5832–5840. [Google Scholar] [CrossRef]
- Abu-Median, A.B.; Van Diemen, P.M.; Dziva, F.; Vlisidou, I.; Wallis, T.S.; Stevens, M.P. Functional analysis of lymphostatin homologues in enterohaemorrhagic Escherichia coli. FEMS Microbiol. Lett. 2006, 258, 43–49. [Google Scholar] [CrossRef]
- Schmidt, H.; Zhang, W.-L.; Hemmrich, U.; Jelacic, S.; Brunder, W.; Tarr, P.I.; Dobrindt, U.; Hacker, J.; Karch, H. Identification and characterization of a novel genomic island integrated at selC in locus of enterocyte effacement-negative, Shiga toxin-producing Escherichia coli. Infect. Immun. 2001, 69, 6863–6873. [Google Scholar] [CrossRef]
- Nakano, M.; Iida, T.; Ohnishi, M.; Kurokawa, K.; Takahashi, A.; Tsukamoto, T.; Yasunaga, T.; Hayashi, T.; Honda, T. Association of the urease gene with enterohemorrhagic Escherichia coli strains irrespective of their serogroups. J. Clin. Microbiol. 2001, 39, 4541–4543. [Google Scholar] [CrossRef]
- Taylor, D.E.; Rooker, M.; Keelan, M.; Ng, L.-K.; Martin, I.; Perna, N.T.; Burland, N.T.V.; Blattner, F.R. Genomic variability of O islands encoding tellurite resistance in enterohemorrhagic Escherichia coli O157:H7 isolates. J. Bacteriol. 2002, 184, 4690–4698. [Google Scholar] [CrossRef]
- Coombes, B.K.; Wickham, M.E.; Mascarenhas, M.; Gruenheid, S.; Finlay, B.B.; Karmali, M.A. Molecular analysis as an aid to assess the public health risk of non-O157 Shiga toxin-producing Escherichia coli strains. Appl. Environ. Microbiol. 2008, 74, 2153–2160. [Google Scholar] [CrossRef]
- Coombes, B.K.; Gilmour, M.W.; Goodman, C.D. The evolution of virulence in non-O157 Shiga toxin-producing Escherichia coli. Front. Microbiol. 2011, 2, 90. [Google Scholar] [CrossRef]
- Karmali, M.A.; Gannon, V.; Sargeant, J.M. Verocytotoxin-producing Escherichia coli (VTEC). Vet. Microbiol. 2010, 140, 360–370. [Google Scholar] [CrossRef]
- Melton-Celsa, A.; Mohawk, K.; Teel, L.; O’Brien, A. Pathogenesis of Shiga-toxin producing Escherichia coli. In Ricin and Shiga Toxins; Springer: Berlin/Heidelberg, Germany, 2011; pp. 67–103. [Google Scholar]
- Schwarz, S.; Kehrenberg, C.; Walsh, T.R. Use of antimicrobial agents in veterinary medicine and food animal production. Int. J. Antimicrob. Agents 2001, 17, 431–437. [Google Scholar] [CrossRef]
- Walsh, C.; Fanning, S. Antimicrobial resistance in foodborne pathogens-a cause for concern? Curr. Drug Targets 2008, 9, 808–815. [Google Scholar] [CrossRef]
- Dibner, J.J.; and Richards, J.D. Antibiotic growth promoters in agriculture: history and mode of action. Poult. Sci. 2005, 84, 634–643. [Google Scholar] [CrossRef]
- Smith, K.E.; Wilker, P.R.; Reiter, P.L.; Hedican, E.B.; Bender, J.B.; Hedberg, C.W. Antibiotic treatment of Escherichia coli O157 infection and the risk of hemolytic uremic syndrome, Minnesota. Pediatr. Infect. Dis. J. 2012, 31, 37–41. [Google Scholar] [CrossRef]
- Melton-Celsa, A.R.; O’Brien, A.D. New therapeutic developments against Shiga toxin-producing Escherichia coli. In Enterohemorrhagic Escherichia coli and Other Shiga Toxin-Producing, E. coli; American Society of Microbiology: Washington, DC, USA, 2015; pp. 361–380. [Google Scholar]
- Browning, N.G.; Botha, J.R.; Sacho, H.; Moore, P.J. Escherichia coli O157:H7 haemorrhagic colitis. Report of the first South African case. S. Afr. J. Surg. 1990, 28, 28–29. [Google Scholar]
- Isaäcson, M.; Canter, P.H.; Effler, P.; Arntzen, L.; Bomans, P.; Heenan, R. Haemorrhagic colitis epidemic in Africa. Lancet 1993, 341, 961. [Google Scholar]
- Riley, L.W.; Remis, R.S.; Helgerson, S.D.; McGee, H.B.; Wells, J.G.; Davis, B.R.; Hebert, R.J.; Olcott, E.S.; Johnson, L.M.; Hargrett, N.T.; et al. Hemorrhagic colitis associated with a rare Escherichia coli serotype. N. Engl. J. Med. 1983, 308, 681–685. [Google Scholar] [CrossRef]
- Mainga, A.O.; Cenci-Goga, B.T.; Malahlela, M.N.; Tshuma, T.; Kalake, A.; Karama, M. Occurrence and characterization of seven major Shiga toxin-producing Escherichia coli serotypes from healthy cattle on cow–calf operations in South Africa. Zoonoses Public Health. 2018, 65, 777–789. [Google Scholar] [CrossRef]
- Rivero, M.A.; Passucci, J.A.; Rodriguez, E.M.; Parma, A.E. Role and clinical course of verotoxigenic Escherichia coli infections in childhood acute diarrhoea in Argentina. J. Med. Microbiol. 2010, 59, 345–352. [Google Scholar] [CrossRef]
- Haugum, K.; Brandal, L.T.; Lindstedt, B.-A.; Wester, A.L.; Bergh, K.; Afset, J.E. PCR-based detection and molecular characterization of shiga toxin-producing Escherichia coli strains in a routine microbiology laboratory over 16 years. J. Clin. Microbiol. 2014, 52, 3156–3163. [Google Scholar] [CrossRef]
- Friedrich, A.W.; Bielaszewska, M.; Zhang, W.-L.; Pulz, M.; Kuczius, T.; Ammon, A.; Karch, H. Escherichia coli harboring Shiga toxin 2 gene variants: frequency and association with clinical symptoms. J. Infect. Dis. 2002, 185, 74–84. [Google Scholar] [CrossRef]
- Boerlin, P.; McEwen, S.A.; Boerlin-Petzold, F.; Wilson, J.B.; Johnson, R.P.; Gyles, C.L. Association between virulence factors of Shiga toxin-producing Escherichia coli and disease in humans. J. Clin. Microbiol. 1999, 37, 497–503. [Google Scholar]
- Feng, P.C.; Jinneman, K.; Scheutz, F.; Monday, S.R. Specificity of PCR and serological assays in the detection of Escherichia coli Shiga toxin subtypes. Appl. Environ. Microbiol. 2011, 77, 6699–6702. [Google Scholar] [CrossRef]
- Iyoda, S.; Manning, S.D.; Seto, K.; Kimata, K.; Isobe, J.; Etoh, Y.; Ichihara, S.; Migita, Y.; Ogata, K.; Honda, M.; et al. Phylogenetic clades 6 and 8 of enterohemorrhagic Escherichia coli O157:H7 with particular stx subtypes are more frequently found in isolates from hemolytic uremic syndrome patients than from asymptomatic carriers. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Kawano, K.; Ono, H.; Iwashita, O.; Kurogi, M.; Haga, T.; Maeda, K.; Goto, Y. Relationship between stx genotype and Stx2 expression level in Shiga toxin-producing Escherichia coli O157 strains. Jpn. J. Infect. Dis. 2012, 65, 322–325. [Google Scholar] [CrossRef][Green Version]
- Orth, D.; Grif, K.; Khan, A.B.; Naim, A.; Dierich, M.P.; Würzner, R. The Shiga toxin genotype rather than the amount of Shiga toxin or the cytotoxicity of Shiga toxin in vitro correlates with the appearance of the hemolytic uremic syndrome. Diagn. Microbiol. Infect. Dis. 2007, 59, 235–242. [Google Scholar] [CrossRef]
- Jenkins, C.; Perry, N.T.; Cheasty, T.; Shaw, D.J.; Frankel, G.; Dougan, G.; Gunn, G.J.; Smith, H.R.; Paton, A.W.; Paton, J.C. Distribution of the saa gene in strains of Shiga toxin-producing Escherichia coli of human and bovine origins. J. Clin. Microbiol. 2003, 41, 1775–1778. [Google Scholar] [CrossRef]
- Aidar-Ugrinovich, L.; Blanco, J.; Blanco, M.; Blanco, J.E.; Leomil, L.; Dahbi, G.; Mora, A.; Onuma, D.L.; Silveira, W.D.; de Castro, A.F.P. Serotypes, virulence genes, and intimin types of Shiga toxin-producing Escherichia coli (STEC) and enteropathogenic E. coli (EPEC) isolated from calves in São Paulo, Brazil. Int. J. Food Microbiol. 2007, 115, 297–306. [Google Scholar] [CrossRef]
- Ju, W.; Shen, J.; Toro, M.; Zhao, S.; Meng, J. Distribution of pathogenicity islands OI-122, OI-43/48, OI-57 and high pathogenicity island (HPI) in Shiga Toxin-producing Escherichia coli. Appl. Environ. Microbiol. 2013, 79, 3406–3412. [Google Scholar] [CrossRef] [PubMed]
- Buvens, G.; De Gheldre, Y.; Dediste, A.; de Moreau, A.-I.; Mascart, G.; Simon, A.; Allemeersch, D.; Scheutz, F.; Lauwers, S.; Piérard, D. Incidence and virulence determinants of verocytotoxin-producing Escherichia coli infections in the Brussels-Capital Region, Belgium, in 2008–2010. J. Clin. Microbiol. 2012, 50, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Buvens, G.; Piérard, D. Virulence profiling and disease association of verocytotoxin-producing Escherichia coli O157 and non-O157 isolates in Belgium. Foodborne Pathog. Dis. 2012, 9, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Bugarel, M.; Beutin, L.; Martin, A.; Gill, A.; Fach, P. Micro-array for the identification of Shiga toxin-producing Escherichia coli (STEC) seropathotypes associated with hemorrhagic colitis and hemolytic uremic syndrome in humans. Int. J. Food Microbiol. 2010, 142, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Cergole-Novella, M.C.; Pignatari, A.C.C.; Castanheira, M.; Guth, B.E.C. Molecular typing of antimicrobial-resistant Shiga-toxin-producing Escherichia coli strains (STEC) in Brazil. Res. Microbiol. 2011, 162, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Mosci, R.E.; Anderson, C.M.; Snyder, B.A.; Collins, J.; Rudrik, J.T.; Manning, S.D. Antimicrobial drug–resistant Shiga toxin–producing Escherichia coli infections, Michigan, USA. Emerg. Infect. Dis. 2017, 23, 1609–1611. [Google Scholar] [CrossRef] [PubMed]
- Day, M.; Doumith, M.; Jenkins, C.; Dallman, T.J.; Hopkins, K.L.; Elson, R.; Godbole, G.; Woodford, N. Antimicrobial resistance in Shiga toxin-producing Escherichia coli serogroups O157 and O26 isolated from human cases of diarrhoeal disease in England, 2015. J. Antimicrobial Chemother. 2017, 72, 145–152. [Google Scholar] [CrossRef]
- Feng, P.C.H.; Monday, S.R. Multiplex PCR for specific identification of enterohemorrhagic Escherichia coli strains in the O157:H7 complex. In Foodborne Pathogens; Humana Press: Totowa, NJ, USA, 2006; pp. 47–55. [Google Scholar]
- Singh, P.; Sha, Q.; Lacher, D.W.; Del Valle, J.; Mosci, R.E.; Moore, J.A.; Scribner, K.T.; Manning, S.D. Characterization of enteropathogenic and Shiga toxin-producing Escherichia coli in cattle and deer in a shared agroecosystem. Front. Cell. Infect. Microbiol. 2015, 5, 29. [Google Scholar] [CrossRef]
- Paton, A.W.; Paton, J.C. Direct detection and characterization of Shiga toxigenic Escherichia coli by multiplex PCR for stx1, stx2, eae, ehxA, and saa. J. Clin. Microbiol. 2002, 40, 271–274. [Google Scholar] [CrossRef]
- Paton, A.W.; Paton, J.C. Multiplex PCR for direct detection of Shiga toxigenic Escherichia coli strains producing the novel subtilase cytotoxin. J. Clin. Microbiol. 2005, 43, 2944–2947. [Google Scholar] [CrossRef]
- Clinical Laboratory Standards. CLSI Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, Approved Standard, 3rd ed.; CLSI document VET01S; National Committee for Clinical Laboratory Standards: Wayne, PA, USA, 2015. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| No | Year | Serotype | stx1 | stx2 | stx2c | stx2d | eaeA | ehxA | katP | espP | etpD | saa | subA |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2007 | O26:H11 | + | − | − | − | + | + | + | + | − | − | − |
| 2 | 2008 | O26:H11 | + | − | − | − | + | + | + | + | − | − | − |
| 3 | 2010 | O111:H8 | + | − | − | − | + | + | + | − | − | − | − |
| 4 | 2010 | O26:H11 | + | − | − | − | + | − | − | − | − | − | − |
| 5 | 2011 | O26:H11 | + | − | − | − | + | + | + | + | − | − | − |
| 6 | 2011 | O157:H7 | − | + | + | + | + | + | + | + | + | − | − |
| 7 | 2010 | O26:H11 | + | − | − | − | + | + | + | + | − | − | − |
| 8 | 2010 | O117:H7 | + | − | − | − | − | − | − | − | − | − | − |
| 9 | 2010 | O157:H7 | − | + | + | + | + | + | + | + | + | − | − |
| 10 | 2003 | ONT:H8 | + | − | − | − | + | − | − | − | − | − | − |
| 11 | 2006 | O111:H8 | + | − | − | − | + | + | − | − | − | − | − |
| 12 | 2006 | O117:H7 | + | − | − | − | − | − | − | − | − | − | − |
| 13 | 2006 | O117:H7 | + | − | − | − | − | − | − | − | − | − | − |
| 14 | 2007 | O22:H16 | − | + | − | − | − | − | − | − | − | − | − |
| 15 | 2007 | O8:H19 | + | + | + | + | − | + | − | + | − | + | − |
| 16 | 2007 | O107:H7 | + | + | − | + | − | + | − | + | − | + | − |
| 17 | 2007 | O107:H7 | + | + | − | + | − | + | − | + | − | + | − |
| 18 | 2007 | O107:H7 | + | + | − | + | − | + | − | + | − | + | − |
| 19 | 2007 | O113:H21 | − | + | + | + | − | + | − | + | − | + | + |
| 20 | 2009 | ONT:H21 | + | − | − | − | − | − | + | − | − | + | − |
| 21 | 2009 | O111:H8 | + | − | + | − | − | − | − | − | − | − | − |
| 22 | 2009 | O111:H8 | + | − | − | − | + | + | − | − | − | − | − |
| 23 | 2009 | O156:H7 | + | − | − | − | − | − | − | − | − | − | − |
| 24 | 2009 | O26:H11 | + | − | − | − | + | + | + | + | − | − | − |
| 25 | 2009 | O157:H7 | + | + | + | + | + | + | + | + | + | − | − |
| 26 | 2009 | O111:H8 | + | − | − | − | + | + | − | − | − | − | − |
| 27 | 2012 | O117:H7 | + | − | − | − | − | − | − | − | − | − | − |
| 28 | 2012 | O117:H7 | + | − | − | − | − | − | − | − | − | − | − |
| 29 | 2012 | O157:H7 | − | + | + | + | + | + | + | + | + | − | − |
| 30 | 2012 | O157:H7 | − | + | + | + | + | + | + | + | + | − | − |
| 31 | 2013 | O118:H16 | + | − | − | − | + | + | + | + | − | − | − |
| 32 | 2013 | O156:H7 | + | − | − | − | − | − | − | − | − | − | − |
| 33 | 2009 | O26:H11 | + | − | − | − | + | + | + | + | − | − | − |
| 34 | 2013 | O26:H11 | + | − | − | − | + | + | + | + | − | − | − |
| 35 | 2008 | O111:H8 | + | − | − | − | + | + | − | − | − | − | − |
| 36 | 2008 | O26:H11 | + | − | − | − | + | + | + | + | − | − | − |
| 37 | 2010 | O5:HNT | + | − | + | − | + | + | − | + | − | − | − |
| 38 | 2008 | O5:HNT | + | − | + | − | + | + | − | + | + | − | − |
| TOTAL | 32 | 11 | 10 | 10 | 23 | 26 | 16 | 21 | 6 | 6 | 1 | ||
| % Positive | 84 | 29 | 26 | 26 | 61 | 68 | 42 | 55 | 16 | 16 | 3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karama, M.; Cenci-Goga, B.T.; Malahlela, M.; Smith, A.M.; Keddy, K.H.; El-Ashram, S.; Kabiru, L.M.; Kalake, A. Virulence Characteristics and Antimicrobial Resistance Profiles of Shiga Toxin-Producing Escherichia coli Isolates from Humans in South Africa: 2006–2013. Toxins 2019, 11, 424. https://doi.org/10.3390/toxins11070424
Karama M, Cenci-Goga BT, Malahlela M, Smith AM, Keddy KH, El-Ashram S, Kabiru LM, Kalake A. Virulence Characteristics and Antimicrobial Resistance Profiles of Shiga Toxin-Producing Escherichia coli Isolates from Humans in South Africa: 2006–2013. Toxins. 2019; 11(7):424. https://doi.org/10.3390/toxins11070424
Chicago/Turabian StyleKarama, Musafiri, Beniamino T. Cenci-Goga, Mogaugedi Malahlela, Anthony M. Smith, Karen H. Keddy, Saeed El-Ashram, Lawan M. Kabiru, and Alan Kalake. 2019. "Virulence Characteristics and Antimicrobial Resistance Profiles of Shiga Toxin-Producing Escherichia coli Isolates from Humans in South Africa: 2006–2013" Toxins 11, no. 7: 424. https://doi.org/10.3390/toxins11070424
APA StyleKarama, M., Cenci-Goga, B. T., Malahlela, M., Smith, A. M., Keddy, K. H., El-Ashram, S., Kabiru, L. M., & Kalake, A. (2019). Virulence Characteristics and Antimicrobial Resistance Profiles of Shiga Toxin-Producing Escherichia coli Isolates from Humans in South Africa: 2006–2013. Toxins, 11(7), 424. https://doi.org/10.3390/toxins11070424