Deoxynivalenol-3-Glucoside Content Is Highly Associated with Deoxynivalenol Levels in Two-Row Barley Genotypes of Importance to Canadian Barley Breeding Programs


Abstract
1. Introduction
2. Results
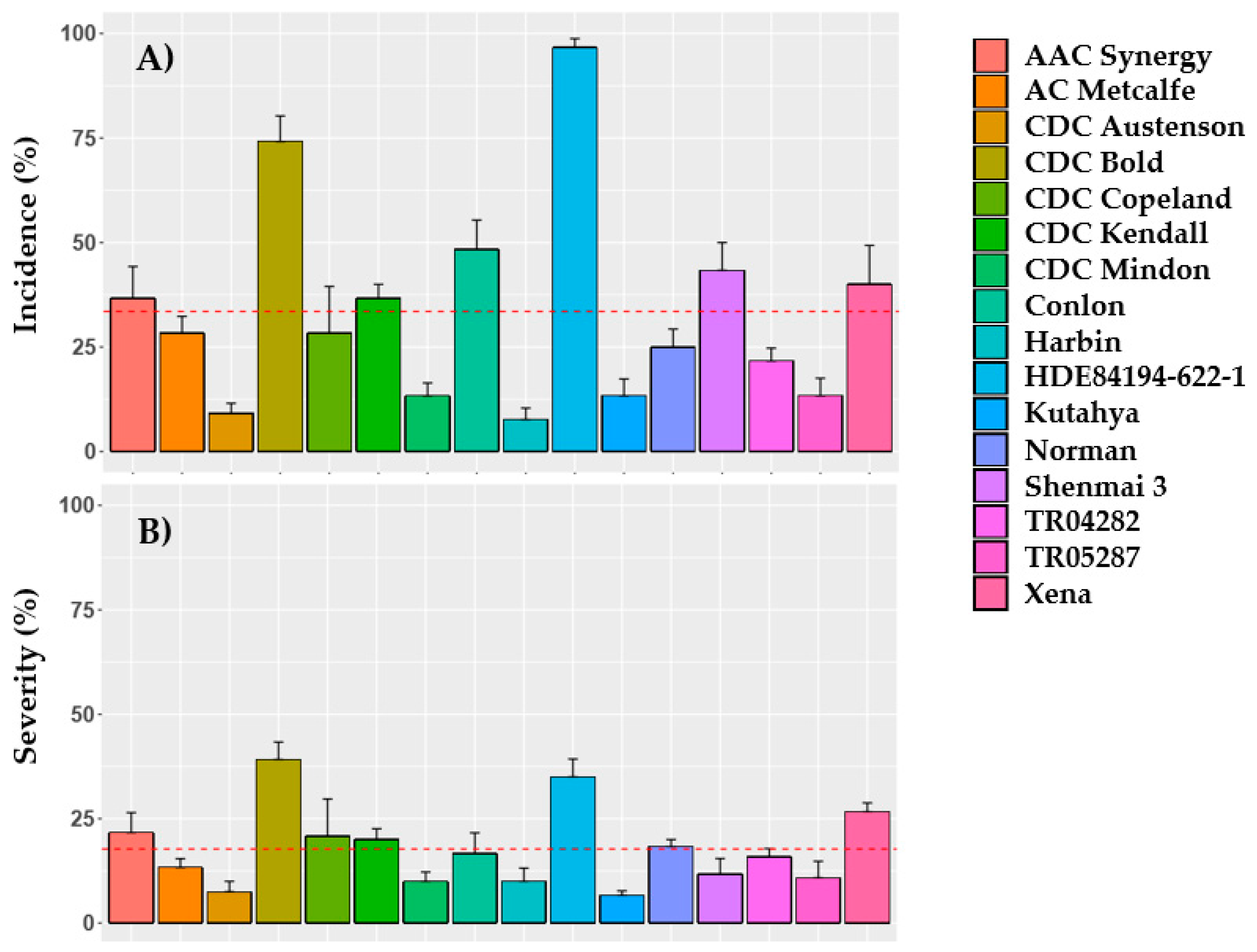
2.1. Heading, Incidence and Severity
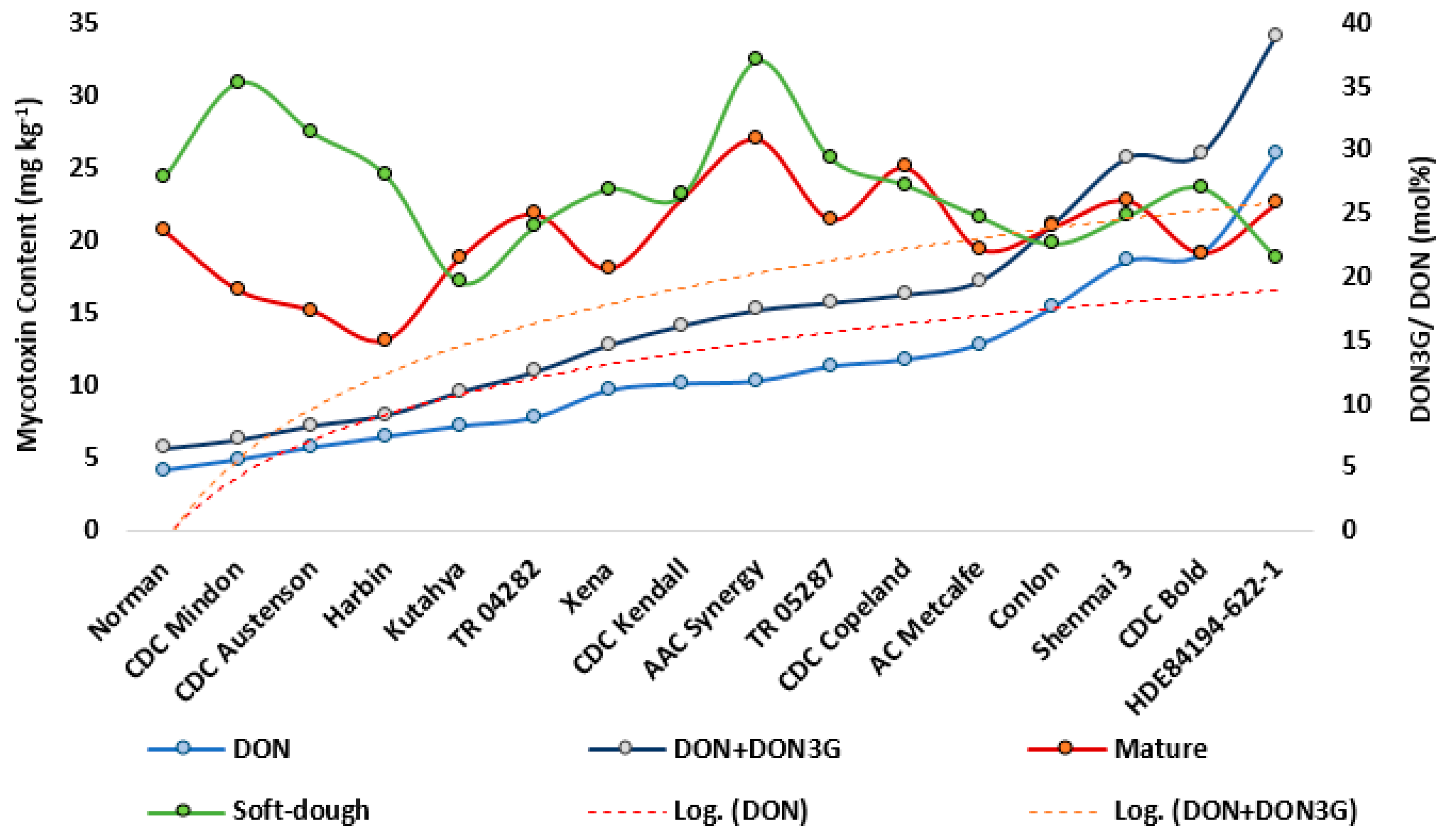
2.2. Mycotoxins
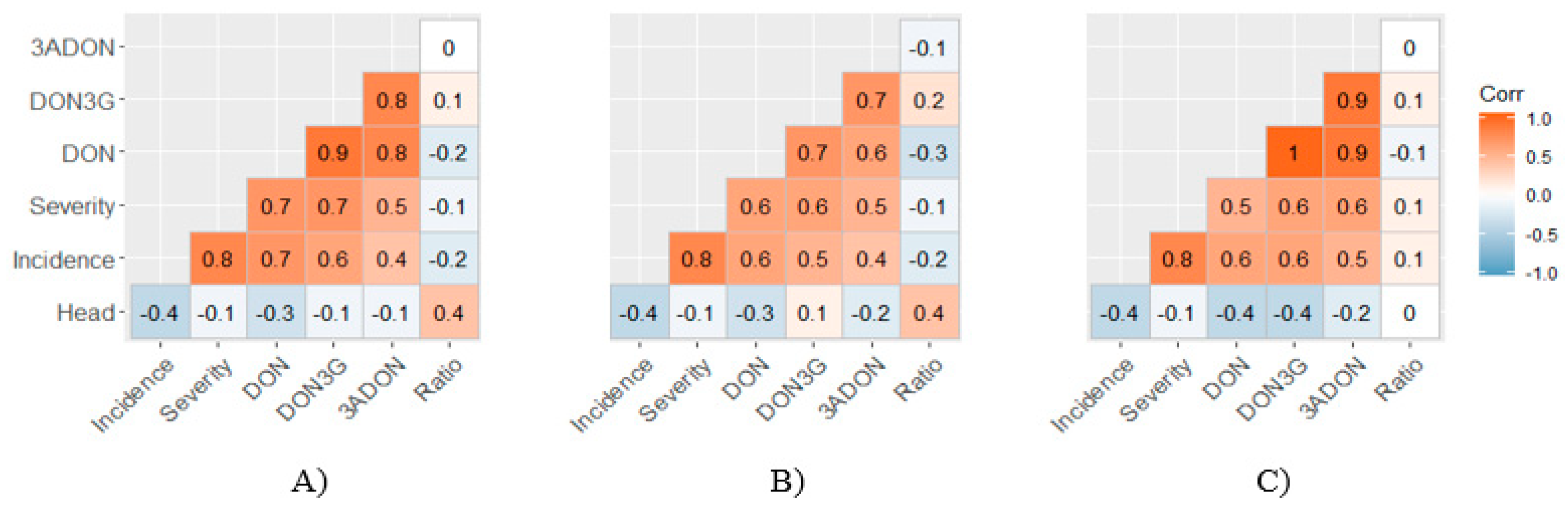
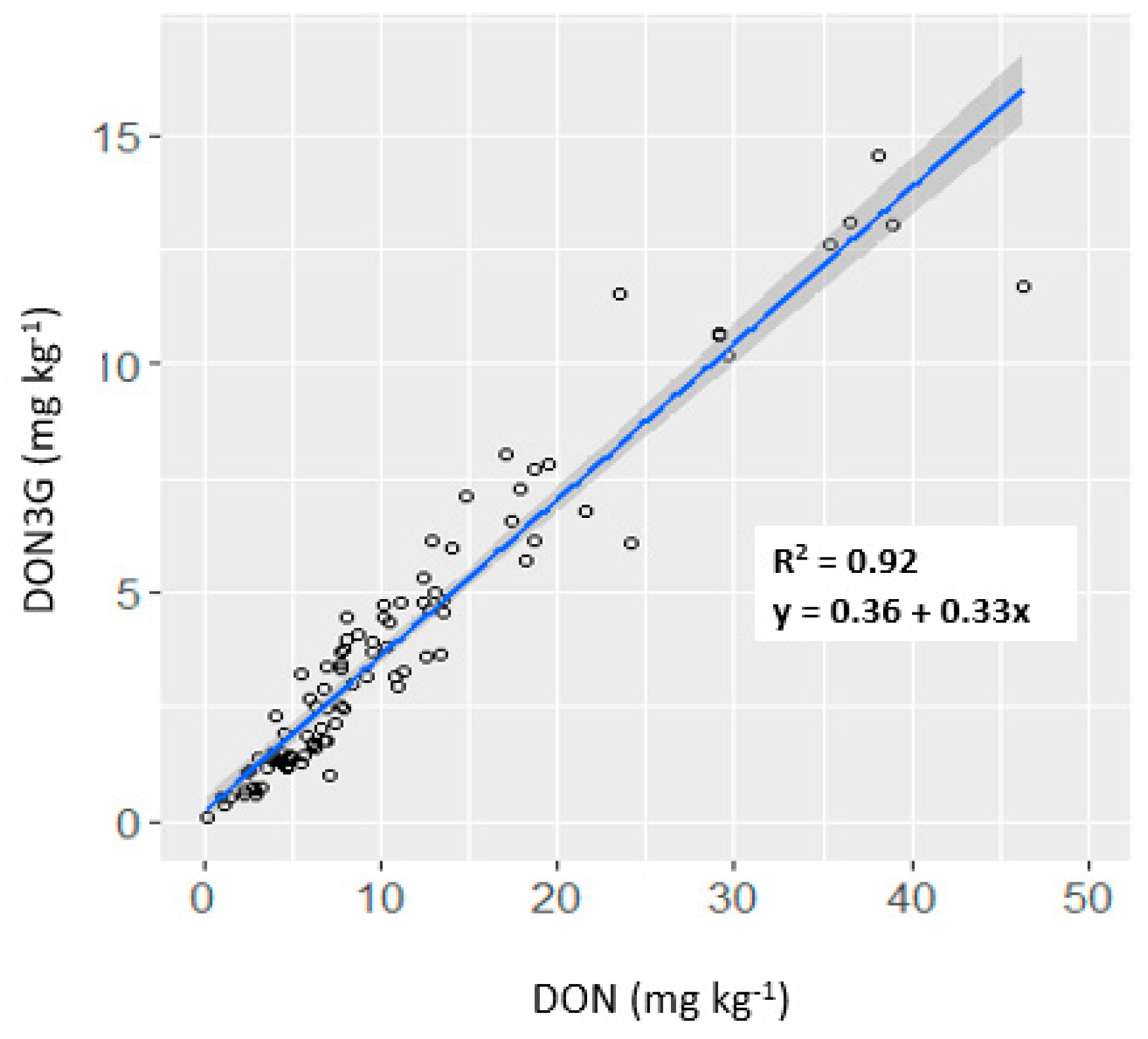
2.3. Relationships between Characters
3. Discussion
4. Materials and Methods
4.1. Inoculum Preparation
4.2. Field Experiment
4.3. Mycotoxin Analysis
4.3.1. Sample Preparation
4.3.2. Extraction and Assay
4.3.3. Quality Control during Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilson, W.; Dahl, B.; Nganje, W. Economic costs of Fusarium Head Blight, scab and deoxynivalenol. World Mycotoxin J. 2018, 11, 291–302. [Google Scholar] [CrossRef]
- Gale, L.R. Population biology of Fusarium species causing head blight in grain crops. In Fusarium Head Blight of Wheat and Barley; Leonard, K.J., Bushnell, W.R., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2003; pp. 120–143. [Google Scholar]
- McCormick, S.P.; Stanley, A.P.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef]
- Rocha, O.; Ansari, K.; Doohan, F.M. Effects of trichothecene mycotoxins on eukaryotic cells: A review. Food Addit. Contam. 2005, 22, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Martel, A.; Pasmans, F.; Ducatelle, R.; Verbrugghe, E.; Vandenbroucke, V.; Li, S.; Haesebrouck, F.; Van Immerseel, F.; Croubels, S. The Impact of Fusarium mycotoxins on human and animal host susceptibility to infectious diseases. Toxins 2014, 6, 430–452. [Google Scholar] [CrossRef] [PubMed]
- Hueza, I.M.; Raspantini, P.C.F.; Raspantini, L.E.R.; Latorre, A.O.; Górniak, S.L. Zearalenone, an estrogenic mycotoxin, is an immunotoxic compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium Toxins in Cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef]
- Kostelanska, M.; Hajslova, J.; Zachariasova, M.; Malachova, A.; Kalachova, K.; Poustka, J.; Fiala, J.; Scott, P.M.; Berthiller, F.; Krskal, R. Occurrence of deoxynivalenol and its major conjugate, deoxynivalenol-3-glucoside, in beer and some brewing intermediates. J. Agric. Food Chem. 2009, 57, 3187–3194. [Google Scholar] [CrossRef]
- Peters, J.; van Dam, R.; van Doorn, R.; Katerere, D.; Berthiller, F.; Haasnoot, W.; Nielen, M.W.F. Mycotoxin profiling of 1000 beer samples with a special focus on craft beer. PLoS ONE 2017, 12, e0185887. [Google Scholar] [CrossRef]
- Trail, F. For blighted waves of grain: Fusarium graminearum in the postgenomics era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef]
- Ma, L.-J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef]
- Boenisch, M.J.; Schäfer, W. Fusarium graminearum forms mycotoxin producing infection structures on wheat. BMC Plant Biol. 2011, 11, 110. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.K.; Xie, W.; Dill-Macky, R.; and Mirocha, C.J. Biosynthesis of deoxynivalenol in spikelets of barley inoculated with macroconidia of Fusarium graminearum. Plant Dis. 2000, 84, 654–660. [Google Scholar] [CrossRef]
- Boddu, J.; Cho, S.; Kruger, W.M.; Muehlbauer, G.J. Transcriptome analysis of the barley–Fusarium graminearum interaction. Mol. Plant Microbe Interact. 2006, 19, 407–417. [Google Scholar] [CrossRef]
- Mesterházy, A. Types and components of resistance to Fusarium Head Blight of wheat. Plant Breed. 1995, 114, 377–386. [Google Scholar] [CrossRef]
- Janssen, E.M.; Liu, C.; Van der Fels-Klerx, H.J. Fusarium infection and trichothecenes in barley and its comparison with wheat. World Mycotoxin J. 2018, 11, 33–46. [Google Scholar] [CrossRef]
- Bai, G.; Shaner, G. Management and resistance in wheat and barley to Fusarium Head Blight. Annu. Rev. Phytopathol. 2004, 42, 135–161. [Google Scholar] [CrossRef]
- Jansen, C.; von Wettstein, D.; Schäfer, W.; Kogel, K.-H.; Felk, A.; Maier, F.J. Infection patterns in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene disrupted Fusarium graminearum. PNAS 2005, 102, 16892–16897. [Google Scholar] [CrossRef] [PubMed]
- Maier, F.J.; Miedaner, T.; Hadeler, B.; Felk, A.; Salomon, S.; Lemmens, M.; Kassner, H.; Schäfer, W. Involvement of trichothecenes in fusarioses of wheat, barley and maize evaluated by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of different chemotype and virulence. Mol. Plant Pathol. 2006, 7, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Langevin, F.; Eudes, F.; Comeau, A. Effect of Trichothecenes Produced by Fusarium graminearum during Fusarium Head Blight development in six cereal species. Eur. J. Plant Pathol. 2004, 110, 735–746. [Google Scholar] [CrossRef]
- Boddu, J.; Cho, S.; Muehlbauer, G.J. Transcriptome Analysis of Trichothecene-Induced Gene Expression in Barley. Mol. Plant Microbe Interact. 2007, 20, 1364–1375. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Kawada, N.; Nakajima, T. Effect of Infection Timing on Fusarium Head Blight and Mycotoxin Accumulation in Open- and Closed-Flowering Barley. Phytopathology 2007, 97, 1054–1062. [Google Scholar] [CrossRef][Green Version]
- Osborne, L.E.; Stein, J.M. Epidemiology of Fusarium head blight on small-grain cereals. Int. J. Food Microbiol. 2007, 119, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Boutigny, A.-L.; Richard-Forget, F.; Barreau, C. Natural mechanisms for cereal resistance to the accumulation of Fusarium trichothecenes. Eur. J. Plant Pathol. 2008, 121, 411–423. [Google Scholar] [CrossRef]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; De Saeger, S.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority) Panel on Contaminants in the Food Chain (CONTAM). Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, e04718. [Google Scholar] [CrossRef]
- Schweiger, W.; Boddu, J.; Shin, S.; Poppenberger, B.; Berthiller, F.; Lemmens, M.; Muehlbauer, G.J.; Adam, G. Validation of a candidate deoxynivalenol-inactivating UDP-glucosyltransferase from barley by heterologous expression in yeast. Mol. Plant Microbe Interact. 2010, 23, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Torres-Acosta, J.A.; Heinen, S.J.; McCormick, S.; Lemmens, M.; Paris, M.P.K.; Berthiller, F.; Adam, G.; Muehlbauer, G.J. Transgenic Arabidopsis thaliana expressing a barley UDP-glucosyltransferase exhibit resistance to the mycotoxin deoxynivalenol. J. Exp. Bot. 2012, 63, 4731–4740. [Google Scholar] [CrossRef]
- Lemmens, M.; Scholz, U.; Berthiller, F.; Dall’Asta, C.; Koutnik, A.; Schuhmacher, R.; Adam, G.; Buerstmayr, H.; Mesterházy, Á.; Krska, R.; et al. The ability to detoxify the mycotoxin deoxynivalenol colocalizes with a major quantitative trait locus for Fusarium head blight resistance in wheat. Mol. Plant Microbe Interact. 2005, 18, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Kluger, B.; Bueschl, C.; Lemmens, M.; Michlmayr, H.; Malachova, A.; Koutnik, A.; Maloku, I.; Berthiller, F.; Adam, G.; Krska, R.; et al. Biotransformation of the mycotoxin deoxynivalenol in Fusarium resistant and susceptible near isogenic wheat lines. PLoS ONE 2015, 10, e0119656. [Google Scholar] [CrossRef]
- Lemmens, M.; Steiner, B.; Sulyok, M.; Nicholson, P.; Mesterhazy, A.; Buerstmayer, H. Masked mycotoxins: Does breeding for enhanced Fusarium head blight resistance result in more deoxynivalenol-3-glucoside in new wheat varieties? World Mycotoxin J. 2016, 9, 741–754. [Google Scholar] [CrossRef]
- Buerstmayr, H.; Lemmens, M. Breeding healthy cereals: Genetic improvement of Fusarium resistance and consequences for mycotoxins. World Mycotoxin J. 2015, 8, 591–602. [Google Scholar] [CrossRef]
- Amarasinghe, C.C.; Senay Simsek, S.; Brûlé-Babel, A.; Fernando, W.G.D. Analysis of deoxynivalenol and deoxynivalenol-3-glucosides content in Canadian spring wheat cultivars inoculated with Fusarium graminearum. Food Addit. Contam. 2016, 33, 1254–1264. [Google Scholar] [CrossRef] [PubMed]
- Soleimany, F.; Jinap, S.; Faridah, A.; Khatib, A. A UPLC–MS/MS for simultaneous determination of aflatoxins, ochratoxin A, zearalenone, DON, fumonisins, T-2 toxin and HT-2 toxin, in cereals. Food Control 2012, 25, 647–653. [Google Scholar] [CrossRef]
- US Wheat and Barley Scab Initiative. Available online: https://scabusa.org/pdfs_dbupload/nabsen-report_05.pdf (accessed on 6 April 2019).
- McMullen, M.; Jones, R.; Gallenberg, D. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Tekauz, A.; McCallum, B.; Gilbert, J. Review: Fusarium head blight of barley in western Canada. Can. J. Plant Pathol. 2000, 22, 9–16. [Google Scholar] [CrossRef]
- Legge, W.G.; Therrien, M.C.; Tucker, J.R.; Banik, M.; Tekauz, A.; Somers, D.; Savard, M.E.; Rossnagel, B.G.; Lefol, E.; Voth, D.; et al. Progress in breeding for resistance to fusarium head blight in barley. Can. J. Plant Pathol. 2004, 26, 436–442. [Google Scholar] [CrossRef]
- Ward, T.J.; Clear, R.M.; Rooney, A.P.; O’Donnell, K.; Gaba, D.; Patrick, S.; Starkey, D.E.; Gilbert, J.; Geiser, D.M.; Nowicki, T.W. An adaptive evolutionary shift in Fusarium head blight pathogen populations is driving the rapid spread of more toxigenic Fusarium graminearum in North America. Fungal Genet. Biol. 2008, 45, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Puri, K.D.; Zhong, S. The 3ADON population of Fusarium graminearum found in North Dakota is more aggressive and produces a higher level of DON than the prevalent 15ADON population in spring wheat. Phytopathology 2010, 100, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Foroud, N.A.; McCormick, S.P.; MacMillan, T.; Badea, A.; Kendra, D.F.; Ellis, B.E.; Eudes, F. Greenhouse studies reveal increased aggressiveness of emergent Canadian Fusarium graminearum chemotypes in wheat. Plant Dis. 2012, 96, 1271–1279. [Google Scholar] [CrossRef]
- Von der Ohe, C.; Gauthier, V.; Tamburic-Ilincic, L.; Brule-Babel, A.; Fernando, W.G.D.; Clear, R.; Ward, T.J.; and Miedaner, T. A comparison of aggressiveness and deoxynivalenol production between Canadian Fusarium graminearum isolates with 3-acetyl and 15-acetyldeoxynivalenol chemotypes in field-grown spring wheat. Eur. J. Plant Pathol. 2010, 127, 407–417. [Google Scholar] [CrossRef]
- Clear, R.M.; Tucker, J.R.; Gaba, D.; Patrick, S.K.; Lee, S.-J.; Demeke, T.; Tittlemier, S.A.; Legge, W.G.; Gräfenhan, T. Deoxynivalenol levels and chemotype frequency in barley cultivars inoculated with two chemotypes of Fusarium graminearum. Can. J. Phytopathol. 2013, 35, 37–45. [Google Scholar] [CrossRef]
- Lewandowski, S.M.; Bushnell, W.R.; Evans, C.K. Distribution of mycelial colonies and lesions in field-grown barley inoculated with Fusarium graminearum. PhytoPathology 2006, 96, 567–581. [Google Scholar] [CrossRef] [PubMed]
- Skadsen, R.W.; Hohn, T.M. Use of Fusarium graminearum transformed with gfp to follow infection patterns in barley and Arabidopsis. Physiol. Mol. Plant Pathol. 2004, 64, 45–53. [Google Scholar] [CrossRef]
- Prom, L.K.; Horsley, R.D.; Steffenson, B.J.; Schwarz, P.B. Development of fusarium head blight and accumulation of deoxynivalenol in barley sampled at different growth stages. J. Am. Soc. Brew. Chem. 1999, 57, 60–63. [Google Scholar] [CrossRef]
- Cowger, C.; Arellano, C. Fusarium graminearum infection and deoxynivalenol concentrations during development of wheat spikes. Phytopathology 2013, 103, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Imboden, L.; Afton, D.; Trail, F. Surface interactions of Fusarium graminearum on barley. Mol. Plant Pathol. 2018, 19, 1332–1342. [Google Scholar] [CrossRef]
- Gardiner, S.A.; Boddu, J.; Berthiller, F.; Hametner, C.; Stupar, R.M.; Adam, G.; Muehlbauer, G.J. Transcriptome analysis of the barley–deoxynivalenol interaction: Evidence for a role of glutathione in deoxynivalenol detoxification. Mol. Plant Microbe Interact. 2010, 23, 962–976. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sanghyun, S.; Heinen, S.; Dill-Macky, R.; Berthiller, F.; Nersesian, N.; Clemente, T.; McCormick, S.; Muehlbauer, G. Transgenic wheat expressing a barley UDP-glucosyltransferase detoxifies deoxynivalenol and provides high levels of resistance to Fusarium graminearum. Mol. Plant Microbe Interact. 2015, 28, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, L.; Smith, K.P.; Muehlbauer, G.J. Differential transcriptomic responses to Fusarium graminearum infection in two barley quantitative trait loci associated with Fusarium head blight resistance. BMC Genom. 2016, 17, 387. [Google Scholar] [CrossRef]
- Ponts, N.; Pinson-Gadais, L.; Barreau, C.; Richard-Forget, F.; Ouellet, T. Exogenous H2O2 and catalase treatments interfere with Tri genes expression in liquid cultures of Fusarium graminearum. FEBS Lett. 2007, 581, 443–447. [Google Scholar] [CrossRef]
- Schwartz, P.; Qian, S.Y.; Zhou, B.; Xu, Y.; Barr, J.M.; Horsley, R.; Gillespie, J. Occurrence of deoxynivalenol-3-glucoside on barley from the upper midwestern United States. J. Am. Soc. Brew. Chem. 2014, 72, 208–213. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Tittlemier, S.A.; Blagden, R.; Chan, J.; Gaba, D.; McKendry, T.; Pleskach, K.; Roscoe, M. Fusarium and Alternaria mycotoxins in Canadian wheat and durum harvest samples. Can. J. Plant Pathol. 2019, in press. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (Alias 1) | Pedigree | Origin 2 | Type 3 | Release 4 |
|---|---|---|---|---|
| AAC Synergy (TR09208) | TR 02267/Newdale | CDN | MALT | 2012 |
| AC Metcalfe (TR232) | AC Oxbow/Manley | CDN | MALT | 1997 |
| CDC Austenson (TR06389) | TR358/94Ab12271 | CDN | GP/F | 2010 |
| CDC Bold (SD422) | SB88403/Tyra | CDN | GP | 1999 |
| CDC Copeland (TR150) | WM861-5/TR118 | CDN | MALT | 1999 |
| CDC Kendall (TR133) | Manley/SM85221 | CDN | MALT | 1999 |
| CDC Mindon (TR04378) | TR339/TR251 | CDN | GP | 2008 |
| Conlon (TR982) | Bowman*2/Birgitta dwarf//ND10232 | USA | MALT 5 | 2004 |
| Harbin | CHN | - | - | |
| HDE84194-622-1 | CHN | - | - | |
| Kutahya | NLD | - | - | |
| Norman (TR05915) | In vitro selection from CDC Kendall | CDN | MALT | 2009 |
| Shenmai 3 | Gobernadora /Humai 10 | CHN | - | - |
| TR04282 | Harbin/TR253//TR253 | CDN | MALT | - |
| TR05287 | Svanhals/AC Metcalfe//TR253 | CDN | MALT | - |
| Xena (TR975) | Stark/Baroness | USA | GP | 2002 |
| Source | DON | DON3G | 3ADON | DON3G/DON | ||||
|---|---|---|---|---|---|---|---|---|
| F | Pr > F | F | Pr > F | F | Pr > F | F | Pr > F | |
| (A) Grain–Soft-dough | ||||||||
| Genotype | 4.09 | <0.0001 | 4.72 | <0.0001 | 5.27 | <0.0001 | 2.21 | 0.02 |
| Year | 2.96 | 0.16 | 5.58 | 0.08 | 16.92 | 0.01 | 6.74 | 0.01 |
| Genotype x Year | 2.17 | 0.02 | 2.51 | 0.01 | 9.15 | <0.0001 | 1.11 | 0.37 |
| (B) Rachis–Soft-dough | ||||||||
| Genotype | 3.63 | 0.0002 | 2.43 | 0.01 | 2.12 | 0.03 | 4.72 | <0.0001 |
| Year | 5.81 | 0.07 | 1.38 | 0.3 | 3.43 | 0.07 | 12.01 | <0.001 |
| Genotype x Year | 1.5 | 0.13 | 2.1 | 0.02 | 5.24 | < 0.0001 | 3.51 | <0.001 |
| (C) Grain–Mature | ||||||||
| Genotype | 4.31 | <0.0001 | 5.56 | <0.0001 | 3.14 | 0.001 | 9.51 | <0.0001 |
| Year | 5.36 | 0.08 | 10.07 | 0.03 | 16.39 | 0.02 | 23.04 | <0.0001 |
| Genotype x Year | 3.91 | <0.0001 | 2.53 | 0.01 | 4.32 | <0.0001 | 3.09 | <0.001 |
| Genotype | DON | DON3G | 3ADON | DON3G/DON | ||||
|---|---|---|---|---|---|---|---|---|
| (A) Grain–Soft-dough | ||||||||
| AAC Synergy | 19.5 | abc | 11.2 | a | 1.5 | a | 37% | a |
| AC Metcalfe | 12.9 | abc | 4.9 | ab | 1.4 | a | 25% | ab |
| CDC Austenson | 4.2 | bc | 2.1 | ab | 0.4 | a | 31% | ab |
| CDC Bold | 33.2 | ab | 14.0 | a | 2.8 | a | 27% | ab |
| CDC Copeland | 9.0 | abc | 3.3 | ab | 0.6 | a | 27% | ab |
| CDC Kendall | 11.0 | abc | 4.4 | ab | 1.0 | a | 26% | ab |
| CDC Mindon | 4.0 | c | 2.7 | b | 0.5 | a | 35% | ab |
| Conlon | 17.3 | abc | 6.0 | ab | 0.9 | a | 23% | ab |
| Harbin | 9.2 | c | 2.5 | b | 0.8 | a | 28% | ab |
| HDE84194-622-1 | 32.8 | a | 10.9 | a | 1.8 | a | 22% | ab |
| Kutahya | 5.8 | abc | 1.8 | b | 0.5 | a | 20% | b |
| Norman | 3.8 | c | 1.8 | b | 0.3 | a | 28% | ab |
| Shenmai 3 | 17.8 | abc | 5.7 | ab | 1.4 | a | 25% | ab |
| TR 04282 | 14.4 | abc | 6.0 | ab | 1.5 | a | 24% | ab |
| TR 05287 | 14.7 | abc | 6.4 | ab | 1.3 | a | 29% | ab |
| Xena | 11.4 | abc | 4.4 | ab | 1.0 | a | 27% | ab |
| Overall Mean | 13.8 | 5.5 | 1.1 | 27% | ||||
| (B) Rachis–Soft-dough | ||||||||
| AAC Synergy | 34.0 | ab | 40.1 | a | 2.3 | a | 89% | abc |
| AC Metcalfe | 17.7 | abc | 15.2 | ab | 1.8 | a | 73% | abc |
| CDC Austenson | 6.1 | bc | 7.4 | ab | 0.8 | a | 95% | ab |
| CDC Bold | 28.7 | abc | 37.7 | a | 2.5 | a | 88% | abc |
| CDC Copeland | 13.0 | abc | 13.7 | ab | 0.8 | a | 69% | abc |
| CDC Kendall | 15.8 | abc | 16.3 | ab | 1.3 | a | 99% | abc |
| CDC Mindon | 8.6 | abc | 8.0 | ab | 1.0 | a | 75% | abc |
| Conlon | 26.5 | abc | 10.1 | ab | 1.1 | a | 48% | cd |
| Harbin | 12.8 | c | 11.8 | ab | 1.1 | a | 111% | a |
| HDE84194-622-1 | 48.8 | a | 22.9 | ab | 3.7 | a | 29% | d |
| Kutahya | 5.6 | bc | 5.9 | b | 0.3 | a | 57% | bcd |
| Norman | 5.1 | c | 5.6 | ab | 0.3 | a | 85% | abc |
| Shenmai 3 | 17.2 | abc | 9.2 | ab | 1.0 | a | 35% | cd |
| TR 04282 | 18.1 | abc | 16.5 | ab | 1.9 | a | 60% | abcd |
| TR 05287 | 20.3 | abc | 17.8 | ab | 1.7 | a | 65% | abc |
| Xena | 16.6 | abc | 15.8 | ab | 1.5 | a | 61% | abc |
| Overall Mean | 18.4 | 15.9 | 1.5 | 71% | ||||
| (C) Grain–Mature | ||||||||
| AAC Synergy | 10.3 | abc | 4.9 | abcd | 0.6 | ab | 31% | a |
| AC Metcalfe | 12.9 | abc | 4.4 | abcd | 0.7 | ab | 22% | bcd |
| CDC Austenson | 5.7 | bc | 1.6 | cde | 0.3 | b | 17% | f |
| CDC Bold | 19.2 | ab | 6.9 | abc | 1.2 | a | 22% | bcd |
| CDC Copeland | 11.8 | abc | 4.5 | abcde | 0.8 | ab | 29% | ab |
| CDC Kendall | 10.1 | abc | 4.1 | abcde | 0.6 | ab | 26% | abc |
| CDC Mindon | 4.9 | c | 1.4 | e | 0.3 | ab | 19% | df |
| Conlon | 15.5 | ab | 5.8 | ab | 0.6 | ab | 23% | abcd |
| Harbin | 6.5 | bc | 1.5 | d | 0.3 | ab | 15% | f |
| HDE84194-622-1 | 26.0 | a | 8.2 | a | 1.1 | a | 26% | abcd |
| Kutahya | 7.2 | abc | 2.4 | abcde | 0.3 | ab | 22% | bcd |
| Norman | 4.1 | c | 1.6 | bcde | 0.2 | b | 24% | abcd |
| Shenmai 3 | 18.6 | ab | 7.1 | a | 0.7 | ab | 26% | abc |
| TR 04282 | 7.8 | abc | 3.2 | abcde | 0.6 | ab | 25% | abcd |
| TR 05287 | 9.9 | abc | 3.9 | abcde | 0.5 | ab | 25% | abcd |
| Xena | 9.7 | abc | 3.2 | abcde | 0.5 | ab | 21% | cd |
| Overall Mean | 11.3 | 4.0 | 0.6 | 23% | ||||
© 2019 by Her Majesty the Queen in Right of Canada as represented by the Minister of Agriculture and Agri-Food Canada; Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tucker, J.R.; Badea, A.; Blagden, R.; Pleskach, K.; Tittlemier, S.A.; Fernando, W.G.D. Deoxynivalenol-3-Glucoside Content Is Highly Associated with Deoxynivalenol Levels in Two-Row Barley Genotypes of Importance to Canadian Barley Breeding Programs. Toxins 2019, 11, 319. https://doi.org/10.3390/toxins11060319
Tucker JR, Badea A, Blagden R, Pleskach K, Tittlemier SA, Fernando WGD. Deoxynivalenol-3-Glucoside Content Is Highly Associated with Deoxynivalenol Levels in Two-Row Barley Genotypes of Importance to Canadian Barley Breeding Programs. Toxins. 2019; 11(6):319. https://doi.org/10.3390/toxins11060319
Chicago/Turabian StyleTucker, James R., Ana Badea, Richard Blagden, Kerri Pleskach, Sheryl A. Tittlemier, and W. G. Dilantha Fernando. 2019. "Deoxynivalenol-3-Glucoside Content Is Highly Associated with Deoxynivalenol Levels in Two-Row Barley Genotypes of Importance to Canadian Barley Breeding Programs" Toxins 11, no. 6: 319. https://doi.org/10.3390/toxins11060319
APA StyleTucker, J. R., Badea, A., Blagden, R., Pleskach, K., Tittlemier, S. A., & Fernando, W. G. D. (2019). Deoxynivalenol-3-Glucoside Content Is Highly Associated with Deoxynivalenol Levels in Two-Row Barley Genotypes of Importance to Canadian Barley Breeding Programs. Toxins, 11(6), 319. https://doi.org/10.3390/toxins11060319