Fluorescence Spectroscopic Investigation of Competitive Interactions between Quercetin and Aflatoxin B1 for Binding to Human Serum Albumin



 ,
,  and
and
Abstract
:1. Introduction
2. Results
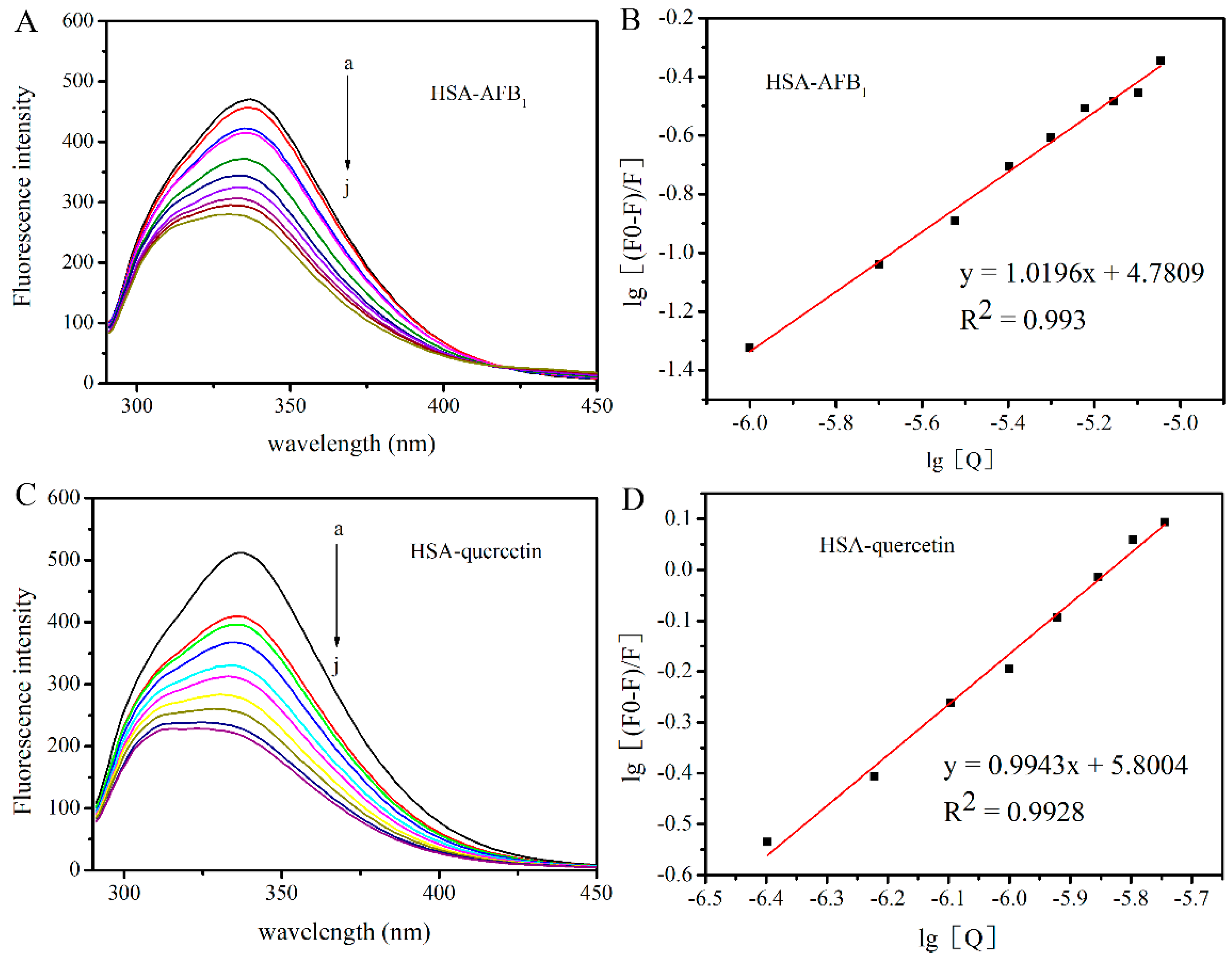
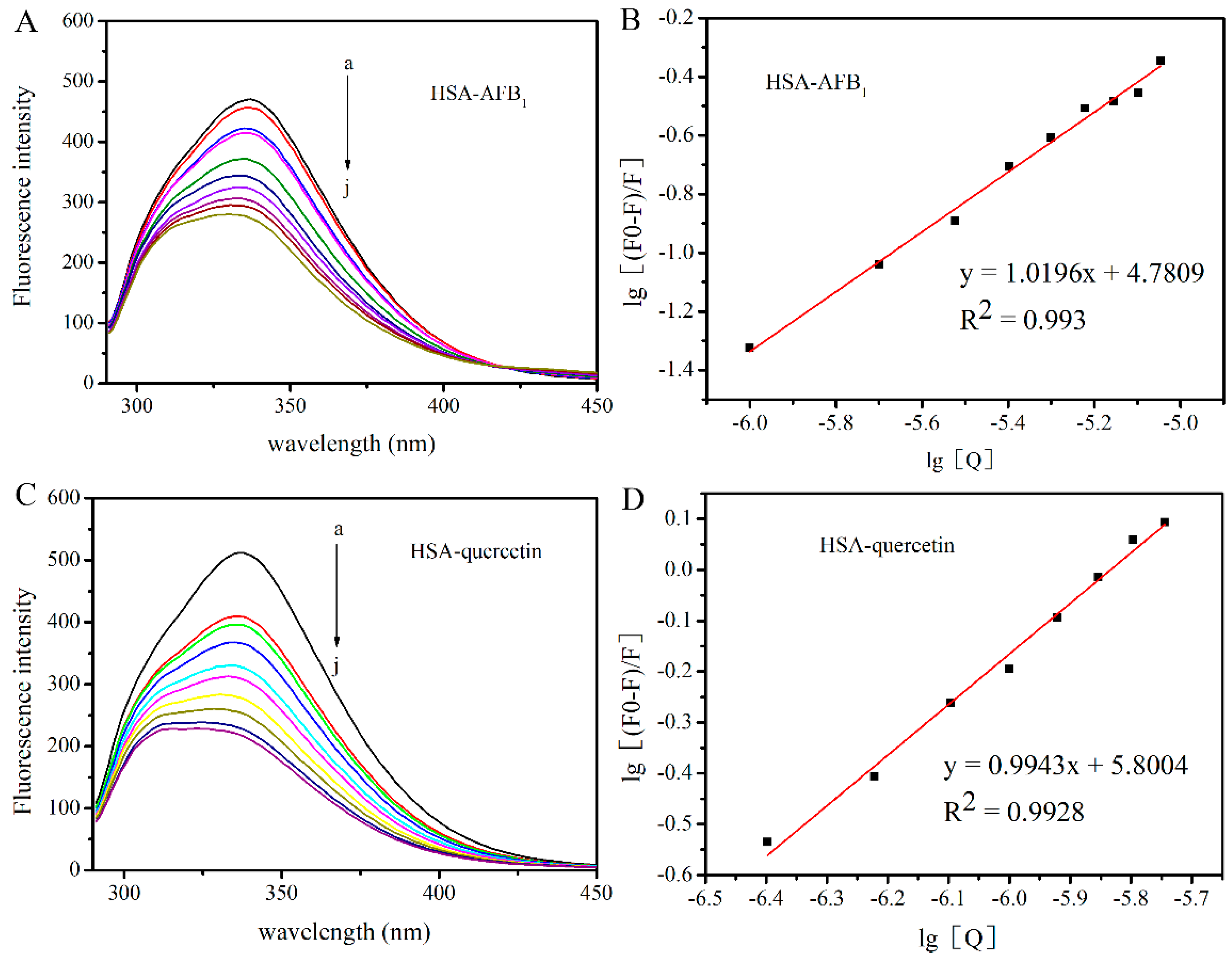
2.1. Investigation of the Interaction between HSA with AFB1 and Quercetin
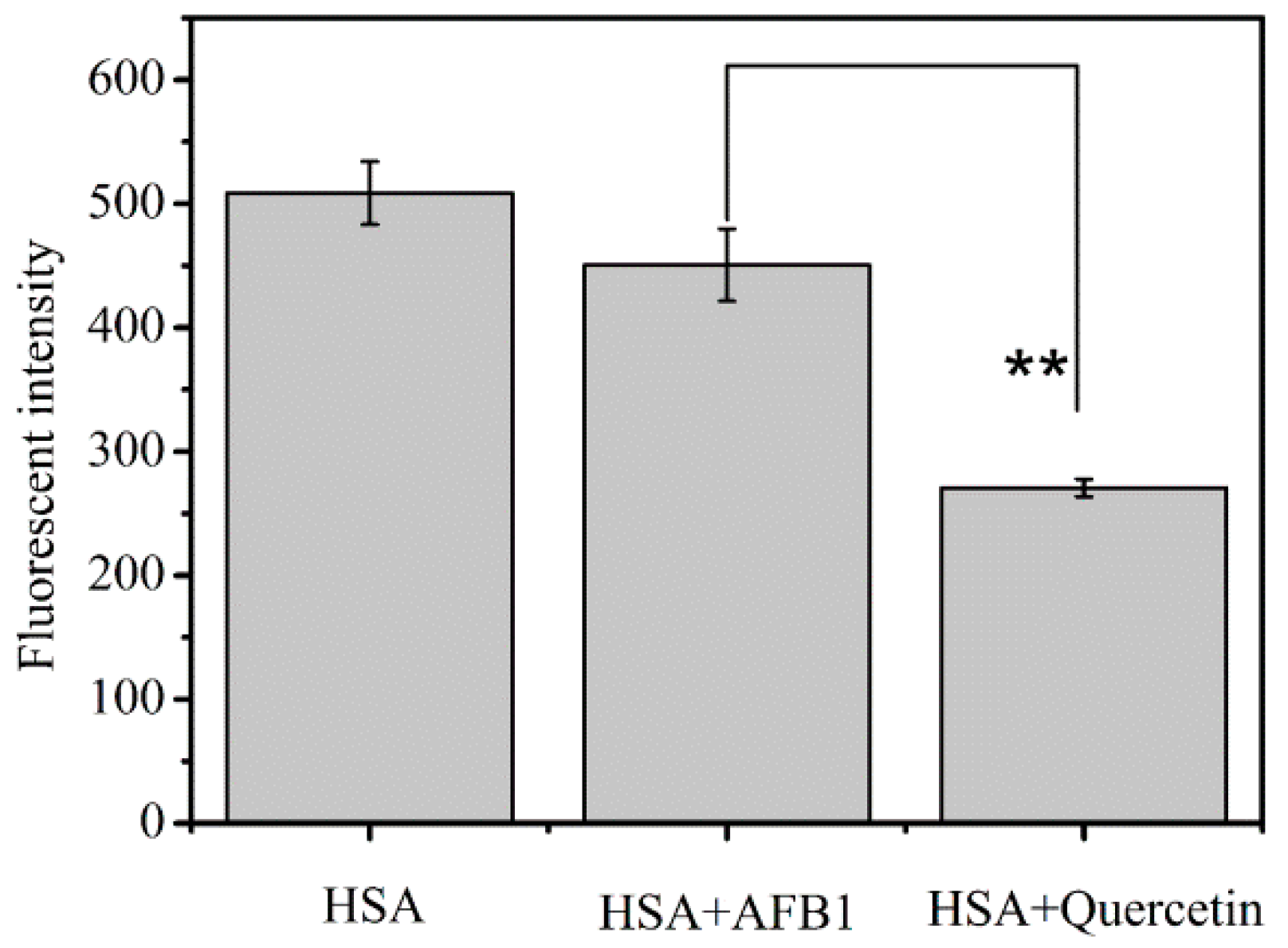
2.2. Investigation of the Competitive Interaction between Quercetin and AFB1 for HSA
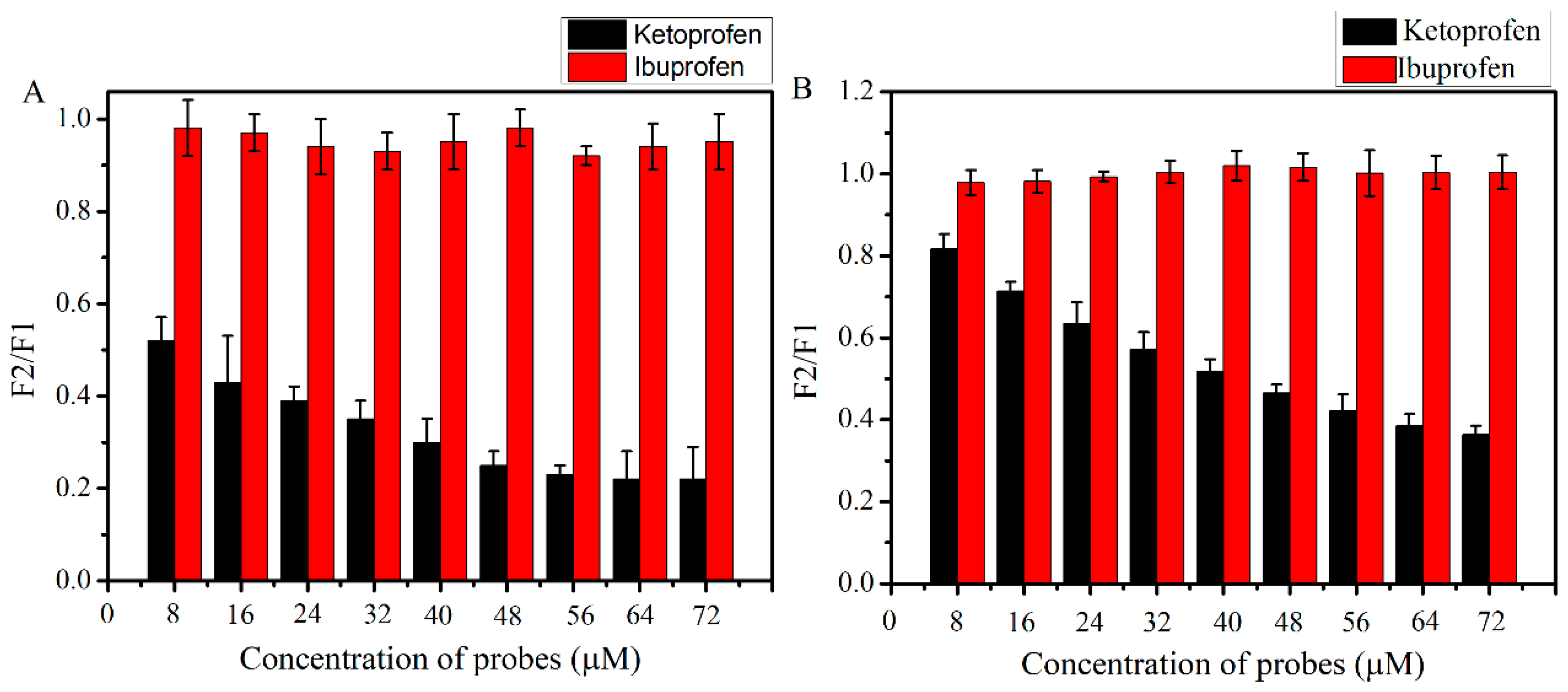
2.3. Ultrafiltration Studies
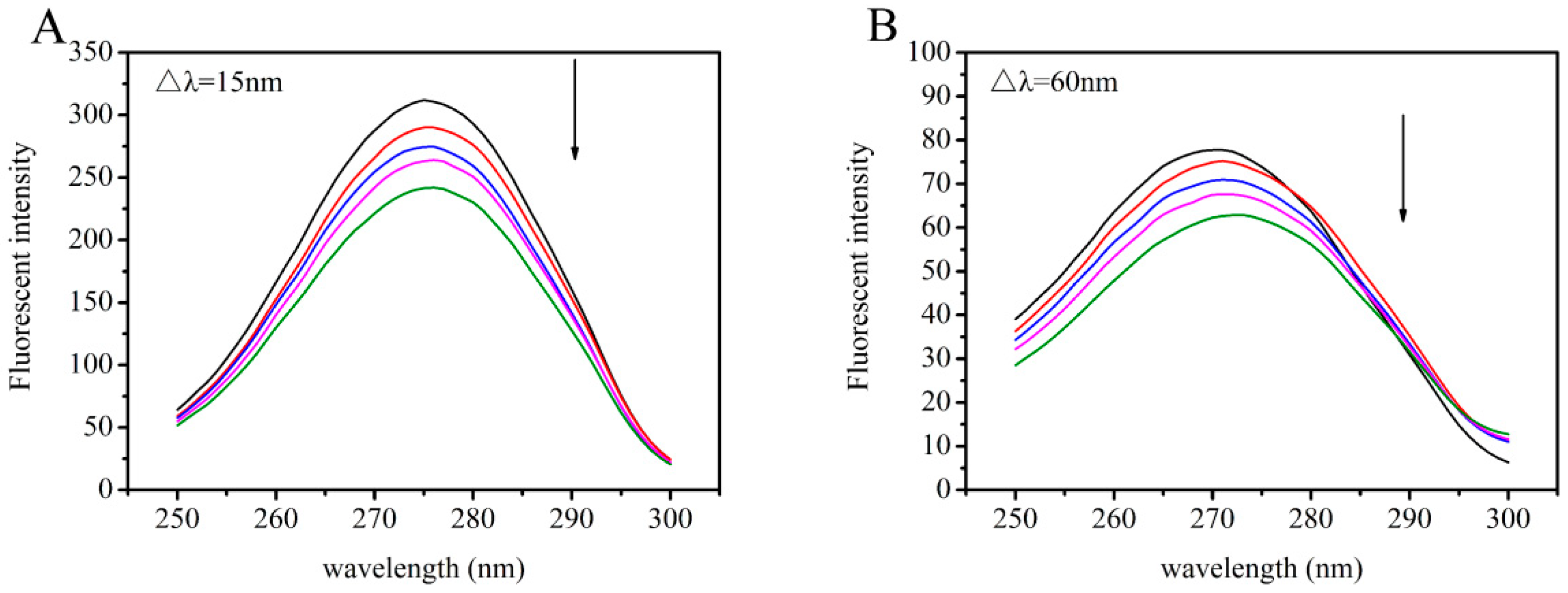
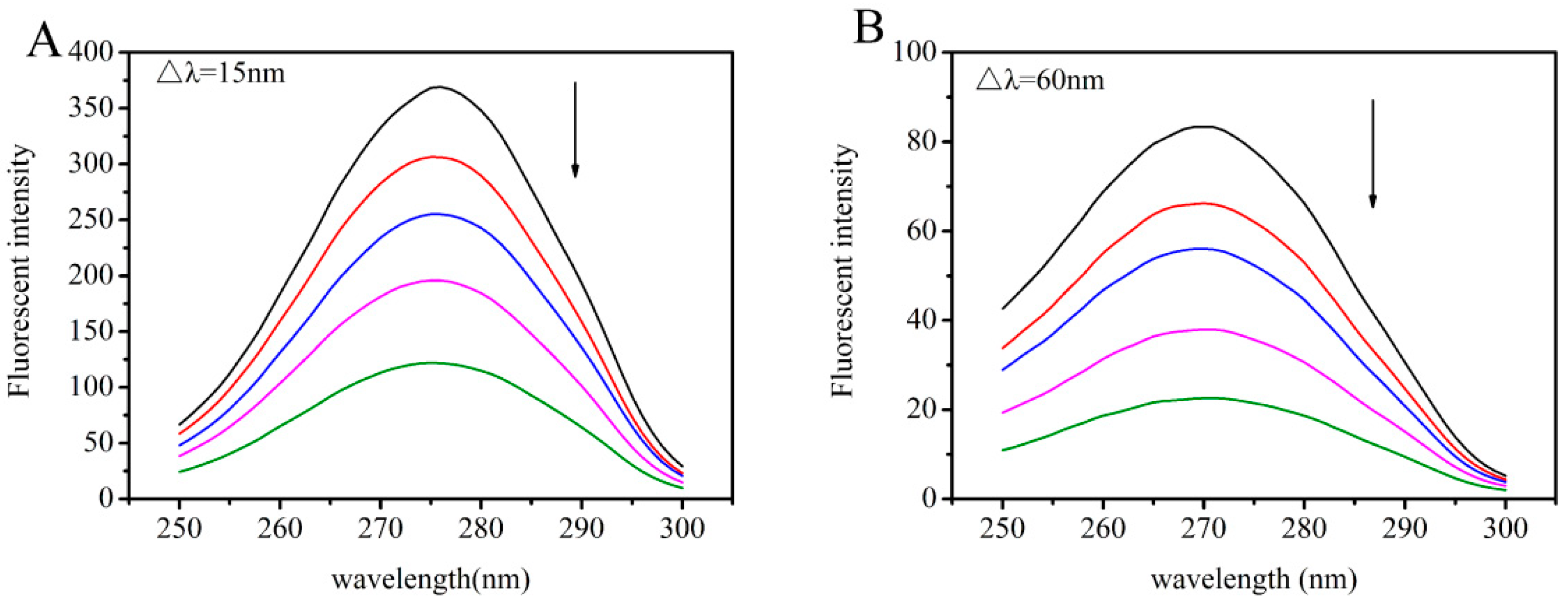
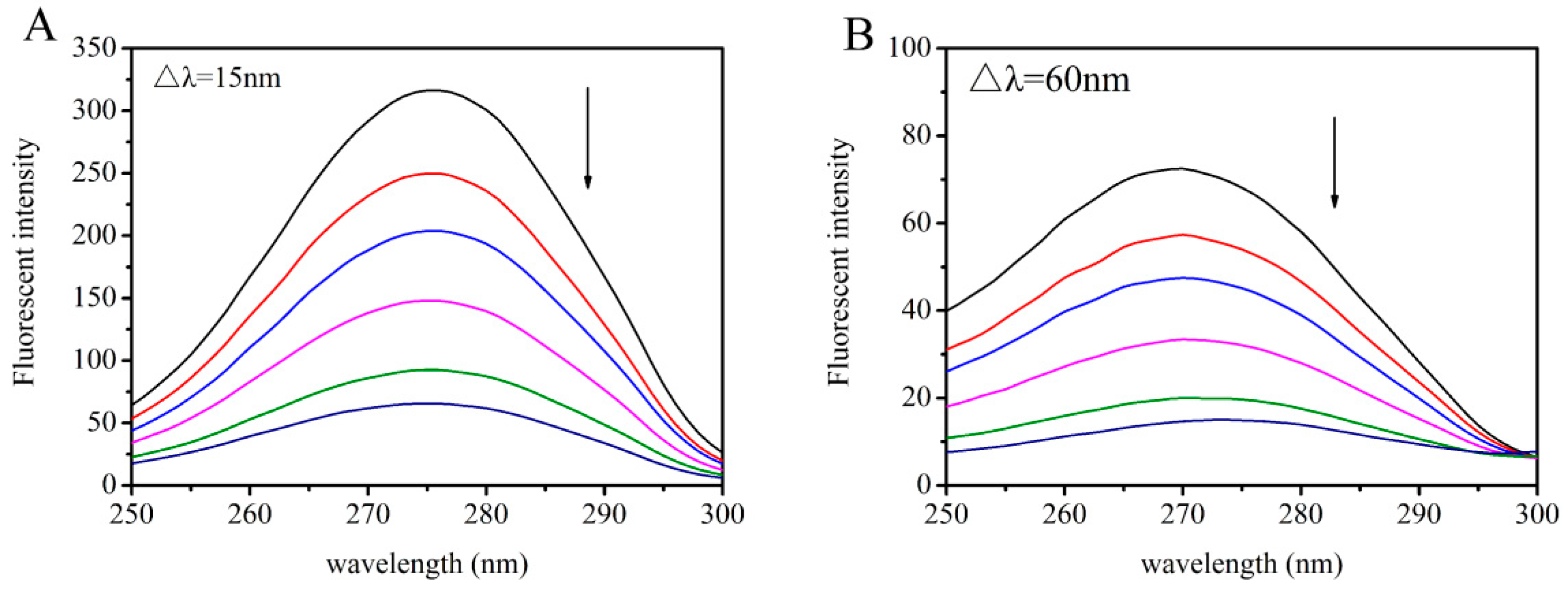
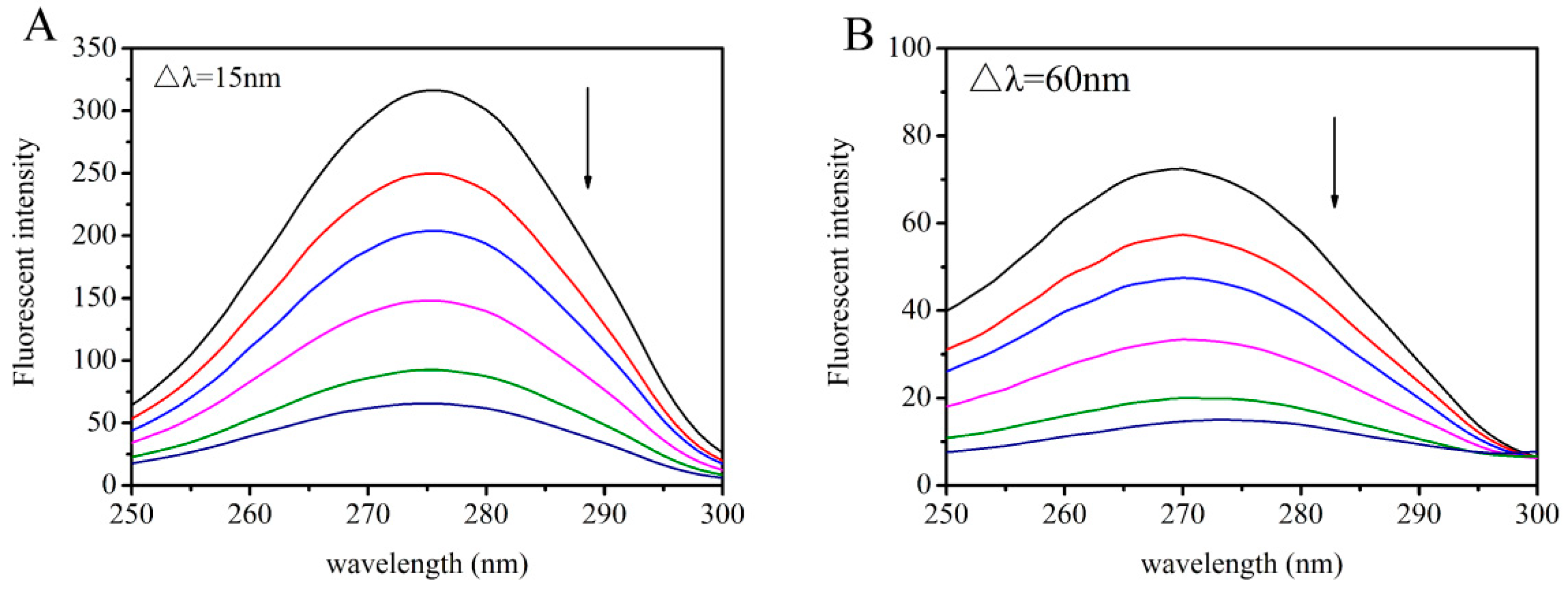
2.4. Synchronous Fluorescence Studies
2.5. The Effect of the Ratio of Quercetin to HSA on the Competitive System
3. Discussion
4. Conclusion
5. Materials and Methods
5.1. Reagents
5.2. Fluorescence Spectroscopic Measurements
5.3. Synchronous Fluorescence
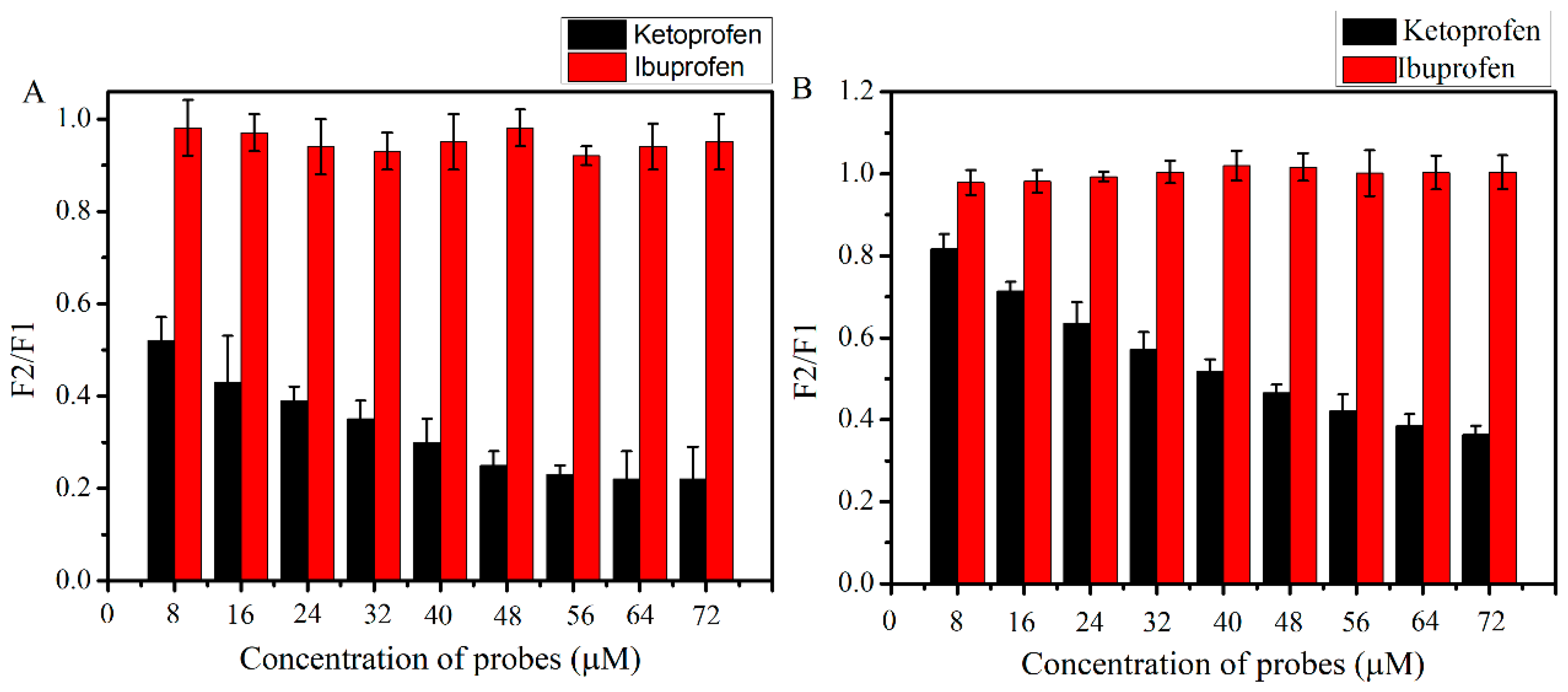
5.4. Site Marker Competitive Experiments
5.5. Ultrafiltration
5.6. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef]
- Dai, Y.Q.; Huang, K.L.; Zhang, B.Y.; Zhu, L.Y.; Xu, W.T. Aflatoxin B1-induced epigenetic alterations: An overview. Food Chem. Toxicol. 2017, 109, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.S.; Tesfamariam, I.G.; Zhang, Y.C.; Zhang, Z.G. Aflatoxin B1-induced hepatocellular carcinoma in developing countries: Geographical distribution, mechanism of action and prevention (Review). Oncol. Lett. 2013, 5, 1087–1092. [Google Scholar]
- Su, C.; Hu, Y.; Gao, D.; Luo, Y.; Chen, A.J.; Jiao, X.; Gao, W. Occurrence of Toxigenic Fungi and Mycotoxins on Root Herbs from Chinese Markets. J. Prot. 2018, 81, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Zhang, L.; Liu, M.; Su, Y.-T.; Xie, W.-M.; Zhang, N.-Y.; Dai, J.-F.; Wang, Y.; Rajput, S.A.; Qi, D.-S.; et al. Individual and Combined Occurrence of Mycotoxins in Feed Ingredients and Complete Feeds in China. Toxins 2018, 10, 113. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, S.; Kaya, E.; Comakli, S. Vitamin E (alpha tocopherol) attenuates toxicity and oxidative stress induced by aflatoxin in rats. Adv. Clin. Exp. Med. 2017, 26, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-Bujaidar, E.; Morales-Gonzalez, J.A.; Sanchez-Gutierrez, M.; Izquierdo-Vega, J.A.; Reyes-Arellano, A.; Alvarez-Gonzalez, I.; Perez-Pasten, R.; Madrigal-Santillan, E. Prevention of Aflatoxin B1-induced dna breaks by beta-D-glucan. Toxins 2015, 7, 2145–2158. [Google Scholar] [CrossRef]
- Mannaa, F.A.; Abdel-Wahhab, K.G.; Abdel-Wahhab, M.A. Prevention of cardiotoxicity of aflatoxin B-1 via dietary supplementation of papaya fruit extracts in rats. Cytotechnology 2014, 66, 327–334. [Google Scholar] [PubMed]
- Selim, K.M.; El-hofy, H.; Khalil, R.H. The efficacy of three mycotoxin adsorbents to alleviate aflatoxin B-1-induced toxicity in Oreochromis niloticus. Aquac. Int. 2014, 22, 523–540. [Google Scholar] [CrossRef]
- Jaynes, W.F.; Zartman, R.E. Aflatoxin Toxicity Reduction in Feed by Enhanced Binding to Surface-Modified Clay Additives. Toxins 2011, 3, 551–565. [Google Scholar] [PubMed]
- Jenkins, T.P.; Fryer, T.; Dehli, R.I.; Jürgensen, J.A.; Fuglsang-Madsen, A.; Føns, S.; Laustsen, A.H. Toxin Neutralization Using Alternative Binding Proteins. Toxins 2019, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Rawal, S.; Kim, J.E.; Coulombe, R., Jr. Aflatoxin B1 in poultry: Toxicology, metabolism and prevention. Res. Sci. 2010, 89, 325–331. [Google Scholar] [CrossRef]
- Ma, L.; Maragos, C.M.; Zhang, Y. Interaction of zearalenone with bovine serum albumin as determined by fluorescence quenching. Mycotoxin Res. 2017, 34, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Bonomo, S.; Jørgensen, F.S.; Olsen, L. Dissecting the Cytochrome P450 1A2- and 3A4-Mediated Metabolism of Aflatoxin B1 in Ligand and Protein Contributions. Chem. A Eur. J. 2017, 23, 2884–2893. [Google Scholar] [CrossRef] [PubMed]
- Galtier, P.; Camguilhem, R.; Bodin, G. Evidence for in vitro and in vivo interaction between ochratoxin A and three acidic drugs. Cosmet. Toxicol. 1980, 18, 493–496. [Google Scholar] [CrossRef]
- Kumagai, S. Ochratoxin A: Plasma concentration and excretion into bile and urine in albumin-deficient rats. Chem. Toxicol. 1985, 23, 941–943. [Google Scholar] [CrossRef]
- Baudrimont, I.; Murn, M.; Betbeder, A.; Guilcher, J.; Creppy, E. Effect of piroxicam on the nephrotoxicity induced by ochratoxin A in rats. Toxicology 1995, 95, 147–154. [Google Scholar] [CrossRef]
- Poór, M.; Kunsági-Máté, S.; Bencsik, T.; Petrik, J.; Vladimir-Knežević, S.; Kőszegi, T. Flavonoid aglycones can compete with Ochratoxin A for human serum albumin: A new possible mode of action. Int. J. Biol. Macromol. 2012, 51, 279–283. [Google Scholar] [CrossRef]
- Poór, M.; Boda, G.; Needs, P.W.; Kroon, P.A.; Lemli, B.; Bencsik, T. Interaction of quercetin and its metabolites with warfarin: Displacement of warfarin from serum albumin and inhibition of CYP2C9 enzyme. Biomed. Pharmacother. 2017, 88, 574–581. [Google Scholar] [CrossRef]
- Di Bari, L.; Ripoli, S.; Pradhan, S.; Salvadori, P. Interactions between quercetin and warfarin for albumin binding: A New Eye on Food/Drug Interference. Chirality 2010, 22, 593–596. [Google Scholar] [CrossRef]
- Jing, J.-J.; Liu, B.; Wang, X.; He, L.-L.; Guo, X.-Y.; Xu, M.-L.; Li, Q.-Y.; Gao, B.; Dong, B.-Y. Binding of fluphenazine with human serum albumin in the presence of rutin and quercetin: An evaluation of food-drug interaction by spectroscopic techniques. Luminescence 2017, 32, 1056–1065. [Google Scholar] [CrossRef]
- Poór, M.; Boda, G.; Kunsági-Máté, S.; Needs, P.W.; Kroon, P.A.; Lemli, B. Fluorescence spectroscopic evaluation of the interactions of quercetin, isorhamnetin, and quercetin-3′-sulfate with different albumins. J. Lumin. 2018, 194, 156–163. [Google Scholar] [CrossRef]
- Das, P.; Chaudhari, S.K.; Das, A.; Kundu, S.; Saha, C. Interaction of flavonols with human serum albumin: A biophysical study showing structure–activity relationship and enhancement when coated on silver nanoparticles. J. Biomol. Struct. Dyn. 2018, 37, 1414–1426. [Google Scholar] [CrossRef]
- Poor, M.; Kunsagi-Mate, S.; Czibulya, Z.; Li, Y.; Peles-Lemli, B.; Petrik, J.; Vladimir-Knezevic, S.; Koszegi, T. Fluorescence spectroscopic investigation of competitive interactions between ochratoxin A and 13 drug molecules for binding to human serum albumin. Luminescence 2013, 28, 726–733. [Google Scholar] [CrossRef]
- Sengupta, B.; Sengupta, P.K. Binding of quercetin with human serum albumin: A critical spectroscopic study. Biopolymers 2003, 72, 427–434. [Google Scholar] [CrossRef]
- Mohseni-Shahri, F.S.; Housaindokht, M.R.; Bozorgmehr, M.R.; Moosavi-Movahedi, A.A. The influence of the flavonoid quercetin on the interaction of propranolol with human serum albumin: Experimental and theoretical approaches. J. Lumin. 2014, 154, 229–240. [Google Scholar] [CrossRef]
- Poór, M.; Bálint, M.; Hetényi, C.; Gődér, B.; Kunsági-Máté, S.; Kőszegi, T.; Lemli, B.; Moretti, A. Investigation of Non-Covalent Interactions of Aflatoxins (B1, B2, G1, G2, and M1) with Serum Albumin. Toxins 2017, 9, 339. [Google Scholar] [CrossRef]
- Pacheco, M.E.; Bruzzone, L. Interactions between imazethapyr and bovine serum albumin: Spectrofluorimetric study. J. Lumin. 2012, 132, 2730–2735. [Google Scholar] [CrossRef]
- Neamtu, S.; Tosa, N.; Bogdan, M. Spectroscopic investigation of tolmetin interaction with human serum albumin. J. Pharm. Biomed. Anal. 2013, 85, 277–282. [Google Scholar] [CrossRef]
- Nunes, N.M.; Pacheco, A.F.C.; Agudelo, Á.J.P.; Da Silva, L.H.M.; Pinto, M.S.; Hespanhol, M.D.C.; Pires, A.C.D.S. Interaction of cinnamic acid and methyl cinnamate with bovine serum albumin: A thermodynamic approach. Food Chem. 2017, 237, 525–531. [Google Scholar] [CrossRef]
- Kőszegi, T.; Poór, M.; Manderville, R.A.; Pfohl-Leszkowicz, A. Ochratoxin A: Molecular Interactions, Mechanisms of Toxicity and Prevention at the Molecular Level. Toxins 2016, 8, 111. [Google Scholar] [CrossRef]
- Faisal, Z.; Lemli, B.; Szerencses, D.; Kunsagi-Mate, S.; Balint, M.; Hetenyi, C.; Kuzma, M.; Mayer, M.; Poor, M. Interactions of zearalenone and its reduced metabolites alpha-zearalenol and beta-zearalenol with serum albumins: Species differences, binding sites, and thermodynamics. Mycotoxin Res. 2018, 34, 269–278. [Google Scholar] [CrossRef]
- Fanali, G.; Di Masi, A.; Trezza, V.; Marino, M.; Fasano, M.; Ascenzi, P. Human serum albumin: From bench to bedside. Mol. Asp. Med. 2012, 33, 209–290. [Google Scholar] [CrossRef]
- Poór, M.; Li, Y.; Matisz, G.; Kiss, L.; Kunsági-Máté, S.; Kőszegi, T. Quantitation of species differences in albumin–ligand interactions for bovine, human and rat serum albumins using fluorescence spectroscopy: A test case with some Sudlow’s site I ligands. J. Lumin. 2014, 145, 767–773. [Google Scholar] [CrossRef]
- Faisal, Z.; Derdak, D.; Lemli, B.; Kunsagi-Mate, S.; Balint, M.; Hetenyi, C.; Csepregi, R.; Koszegi, T.; Sueck, F.; Cramer, B.; et al. Interaction of 2R-ochratoxin A with Serum Albumins: Binding Site, Effects of Site Markers, Thermodynamics, Species Differences of Albumin-binding, and Influence of Albumin on Its Toxicity in MDCK Cells. Toxins 2018, 10, 353. [Google Scholar] [CrossRef]
- Hagelberg, S.; Hult, K.; Fuchs, R. Toxicokinetics of ochratoxin A in several species and its plasma-binding properties. J. Appl. Toxicol. 1989, 9, 91–96. [Google Scholar] [CrossRef]
- Studer-Rohr, I.; Schlatter, J.; Dietrich, D.R. Kinetic parameters and intraindividual fluctuations of ochratoxin A plasma levels in humans. Arch. Toxicol. 2000, 74, 499–510. [Google Scholar] [CrossRef]
- Rimac, H.; Debeljak, Ž.; Bojić, M.; Miller, L.; Rimac, Ž.D.H. Displacement of Drugs from Human Serum Albumin: From Molecular Interactions to Clinical Significance. Curr. Med. Chem. 2017, 24, 1. [Google Scholar] [CrossRef]
- Yamasaki, K.; Chuang, V.T.G.; Maruyama, T.; Otagiri, M. Albumin–drug interaction and its clinical implication. Biochim. Biophys. (BBA) Gen. Subj. 2013, 1830, 5435–5443. [Google Scholar] [CrossRef]
- Perry, J.L.; Il’ichev, Y.V.; Kempf, V.R.; McClendon, J.; Park, G.; Manderville, R.A.; Ruker, F.; Dockal, M.; Simon, J.D. Binding of ochratoxin A derivatives to human serum albumin. J. Phys. Chem. B 2003, 107, 6644–6647. [Google Scholar] [CrossRef]
- Xiao, J.B.; Kai, G.Y. A Review of dietary polyphenol-plasma protein interactions: Characterization, influence on the bioactivity, and structure-affinity relationship. Crit. Rev. Food Sci. Nutr. 2012, 52, 85–101. [Google Scholar] [CrossRef]
- Tang, F.; Xie, Y.; Cao, H.; Yang, H.; Chen, X.; Xiao, J. Fetal bovine serum influences the stability and bioactivity of resveratrol analogues: A polyphenol-protein interaction approach. Food Chem. 2017, 219, 321–328. [Google Scholar] [CrossRef]
- Xiao, J.B.; Zhao, Y.R.; Wang, H.; Yuan, Y.M.; Yang, F.; Zhang, C.; Yamamoto, K. Noncovalent interaction of dietary polyphenols with common human plasma proteins. J. Agric. Food Chem. 2017, 59, 10747–10754. [Google Scholar] [CrossRef]
- Choi, K.-C.; Chung, W.-T.; Kwon, J.-K.; Yu, J.-Y.; Jang, Y.-S.; Park, S.-M.; Lee, S.-Y.; Lee, J.-C. Inhibitory effects of quercetin on aflatoxin B1-induced hepatic damage in mice. Chem. Toxicol. 2010, 48, 2747–2753. [Google Scholar] [CrossRef]
- El-Nekeety, A.A.; Abdel-Azeim, S.H.; Hassan, A.M.; Hassan, N.S.; Aly, S.E.; Abdel-Wahhab, M.A.; Abdel-Wahhab, P.D.M.A. Quercetin inhibits the cytotoxicity and oxidative stress in liver of rats fed aflatoxin-contaminated diet. Toxicol. Rep. 2014, 1, 319–329. [Google Scholar] [CrossRef]
- Panda, D.; Datta, A. Evidence for covalent binding of epicocconone with proteins from synchronous fluorescence spectra and fluorescence lifetimes. J. Chem. Sci. 2007, 119, 99–104. [Google Scholar] [CrossRef]
- Maurya, N.; Parray, M.U.D.; Maurya, J.K.; Kumar, A.; Patel, R. Interaction of promethazine and adiphenine to human hemoglobin: A comparative spectroscopic and computational analysis. Spectrochim. A Mol. Biomol. Spectrosc. 2018, 199, 32–42. [Google Scholar] [CrossRef]
- Poór, M.; Lemli, B.; Bálint, M.; Hetenyi, C.; Sali, N.; Kőszegi, T.; Kunsagi-Mate, S.; Van Der Fels-Klerx, H. (Ine) Interaction of Citrinin with Human Serum Albumin. Toxins 2015, 7, 5155–5166. [Google Scholar] [CrossRef]
- Kameníková, M.; Furtmüller, P.G.; Klacsová, M.; Lopez-Guzman, A.; Toca-Herrera, J.L.; Vitkovská, A.; Devínsky, F.; Mučaji, P.; Nagy, M. Influence of quercetin on the interaction of gliclazide with human serum albumin-spectroscopic and docking approaches. Luminescence 2017, 32, 1203–1211. [Google Scholar] [CrossRef]
- Bagheri, M.; Fatemi, M.H. Fluorescence spectroscopy, molecular docking and molecular dynamic simulation studies of HSA-Aflatoxin B1 and G1 interactions. J. Lumin. 2018, 202, 345–353. [Google Scholar] [CrossRef]
- Poór, M.; Kunsági-Máté, S.; Bálint, M.; Hetényi, C.; Gerner, Z.; Lemli, B. Interaction of mycotoxin zearalenone with human serum albumin. J. Photochem. Photobiol. B: Biol. 2017, 170, 16–24. [Google Scholar] [CrossRef]
- Wang, J.; Ma, L.; Zhang, Y.; Jiang, T. Investigation of the interaction of deltamethrin (DM) with human serum albumin by multi-spectroscopic method. J. Mol. Struct. 2017, 1129, 160–168. [Google Scholar] [CrossRef]
- Shiri, F.; Rahimi-Nasrabadi, M.; Ahmadi, F.; Ehrlich, H. Multispectroscopic and molecular modeling studies on the interaction of copper-ibuprofenate complex with bovine serum albumin (BSA). Spectrochim. Acta A Mol. Biomol. Spectrosc. 2018, 203, 510–521. [Google Scholar] [CrossRef]
- Nafisi, S.; Vishkaee, T.S. Study on the interaction of tamiflu and oseltamivir carboxylate with human serum albumin. J. Photochem. Photobiol. B: Biol. 2011, 105, 34–39. [Google Scholar] [CrossRef]
- Hao, C.; Xu, G.; Feng, Y.; Lu, L.; Sun, W.; Sun, R. Fluorescence quenching study on the interaction of ferroferric oxide nanoparticles with bovine serum albumin. Spectrochim. A Mol. Biomol. Spectrosc. 2017, 184, 191–197. [Google Scholar] [CrossRef]
- Östlund, J.; Zlabek, V.; Zamaratskaia, G. In vitro inhibition of human CYP2E1 and CYP3A by quercetin and myricetin in hepatic microsomes is not gender dependent. Toxicology 2017, 381, 10–18. [Google Scholar] [CrossRef]
- Sun, M.; Tang, Y.; Ding, T.; Liu, M.; Wang, X. Inhibitory effects of celastrol on rat liver cytochrome P450 1A2, 2C11, 2D6, 2E1 and 3A2 activity. Fitoterapia 2014, 92, 1–8. [Google Scholar] [CrossRef]
- Li, Y.; Czibulya, Z.; Poór, M.; LeComte, S.; Kiss, L.; Harté, E.; Kőszegi, T.; Kunsági-Máté, S. Thermodynamic study of the effects of ethanol on the interaction of ochratoxin A with human serum albumin. J. Lumin. 2014, 148, 18–25. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Ksv (LmoL−1) | Kq (LmoL−1S−1) | Ka (M−1) | n | R2 |
|---|---|---|---|---|---|
| AFB1-HSA | 1.62 × 104 | 1.62 × 1012 | 6.02×104 | 1 | 0.993 |
| Quercetin-HSA | 1.83 × 105 | 1.83 × 1013 | 6.31×105 | 1 | 0.992 |
| Systems | HSA:AFB1:Quercetin | |
|---|---|---|
| 2:1:2 | 1:1:2 | |
| HSA | 494.3 ± 41.6 a | 498.7 ± 61.8 a |
| HSA + AFB1 | 424.7 ± 58.5 b | 444.9 ± 19.4 b |
| HSA + AFB1 + Quercetin | 251.4 ± 17.8 d | 282.7 ± 37.9 c |
| HSA + Quercetin | 292.6 ± 31.0 b, c | 319.8 ± 31.3 b |
| HSA + Quercetin + AFB1 | 254.4 ± 22.9 c | 265.4 ± 38.5 b |
| HSA + (AFB1 + Quercetin) | 315.6 ± 20.7 c | 281.8 ± 49.9 c |
| Systems | HSA:AFB1:Quercetin | |
|---|---|---|
| 2:1:1 | 1:1:1 | |
| HSA | 512.8 ± 5.7 a | 530.9 ± 57.8 a |
| HSA + AFB1 | 433.6 ± 15.2 b | 497.1 ± 23.9 b |
| HSA + AFB1 + Quercetin | 376.3 ± 14.3 c | 375.7 ± 64.1 c |
| HSA + Quercetin | 374.1 ± 16.7 b | 407.3 ± 21.6 b |
| HSA + Quercetin + AFB1 | 360.5 ± 27.5 b | 386.7 ± 64.2 b |
| HSA + (AFB1 + Quercetin) | 398.6 ± 19.9 c | 414.1 ± 23.4 c |
| Systems | HSA:AFB1:Quercetin | |
|---|---|---|
| 2:2:1 | 1:2:1 | |
| HSA | 467.3 ± 40.9 a | 501.8 ± 31.2 a |
| HSA + AFB1 | 405.7 ± 42.9 b | 473.8 ± 2.7 b |
| HSA + AFB1 + Quercetin | 315.9 ± 21.6 c | 37.3 ± 19.4 c |
| HSA + Quercetin | 328.0 ± 21.6 b | 365.5 ± 7.6 b |
| HSA + Quercetin + AFB1 | 362.9 ± 30.4 b | 72.0 ± 43.4 b |
| HSA + (AFB1 + Quercetin) | 362.9 ± 30.4 b | 72.0 ± 43.4 b, c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, H.; Chen, L.; Ma, L.; Liu, S.; Zhou, H.; Zhang, Y.; Guo, T.; Liu, W.; Dai, H.; Yu, Y. Fluorescence Spectroscopic Investigation of Competitive Interactions between Quercetin and Aflatoxin B1 for Binding to Human Serum Albumin. Toxins 2019, 11, 214. https://doi.org/10.3390/toxins11040214
Tan H, Chen L, Ma L, Liu S, Zhou H, Zhang Y, Guo T, Liu W, Dai H, Yu Y. Fluorescence Spectroscopic Investigation of Competitive Interactions between Quercetin and Aflatoxin B1 for Binding to Human Serum Albumin. Toxins. 2019; 11(4):214. https://doi.org/10.3390/toxins11040214
Chicago/Turabian StyleTan, Hongxia, Lu Chen, Liang Ma, Shuang Liu, Hongyuan Zhou, Yuhao Zhang, Ting Guo, Wei Liu, Hongjie Dai, and Yong Yu. 2019. "Fluorescence Spectroscopic Investigation of Competitive Interactions between Quercetin and Aflatoxin B1 for Binding to Human Serum Albumin" Toxins 11, no. 4: 214. https://doi.org/10.3390/toxins11040214
APA StyleTan, H., Chen, L., Ma, L., Liu, S., Zhou, H., Zhang, Y., Guo, T., Liu, W., Dai, H., & Yu, Y. (2019). Fluorescence Spectroscopic Investigation of Competitive Interactions between Quercetin and Aflatoxin B1 for Binding to Human Serum Albumin. Toxins, 11(4), 214. https://doi.org/10.3390/toxins11040214