Quercetin Inhibits the Proliferation and Aflatoxins Biosynthesis of Aspergillus flavus



Abstract
1. Introduction
2. Results
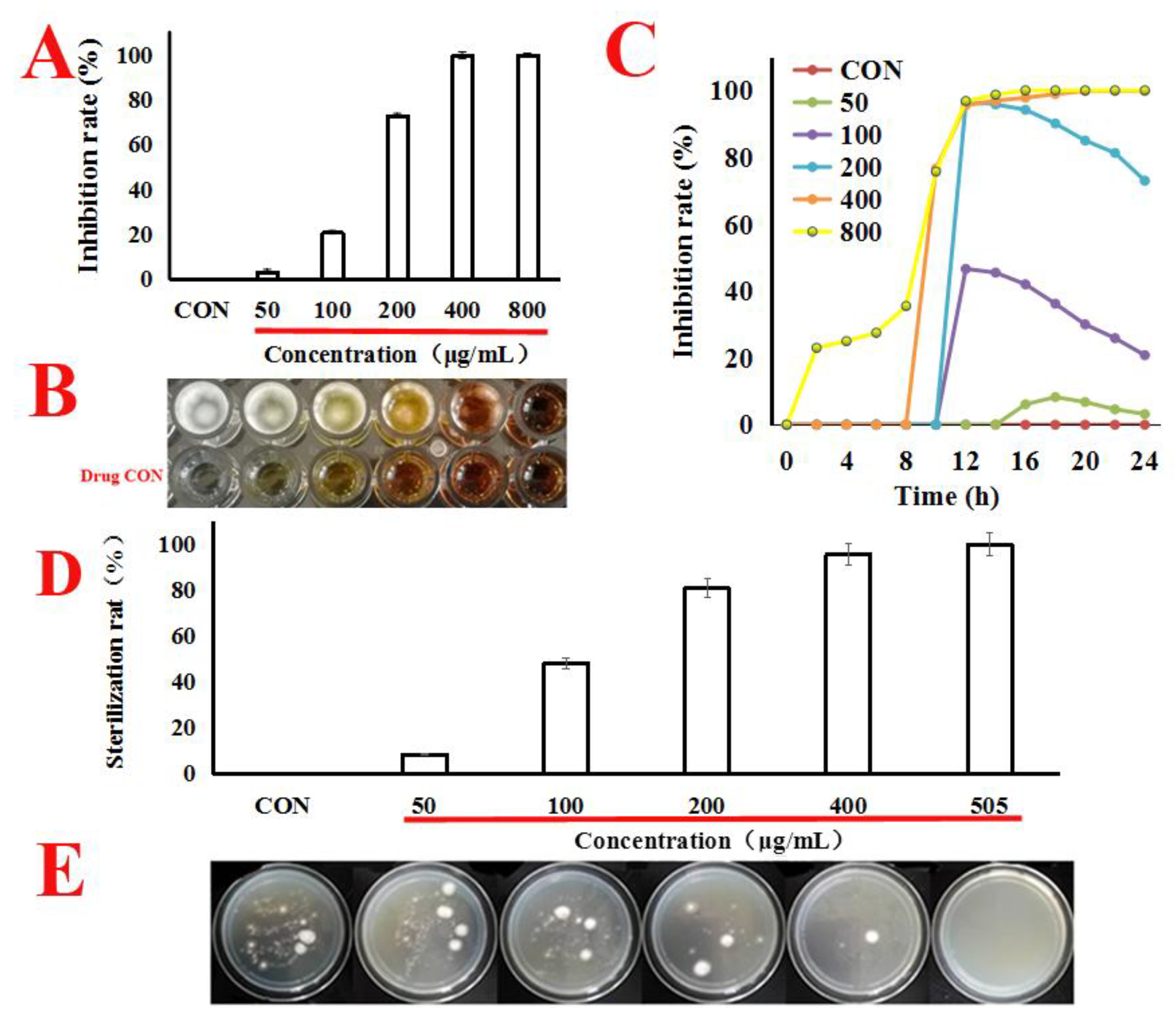
2.1. Quercetin Inhibited the Proliferation of A. flavus
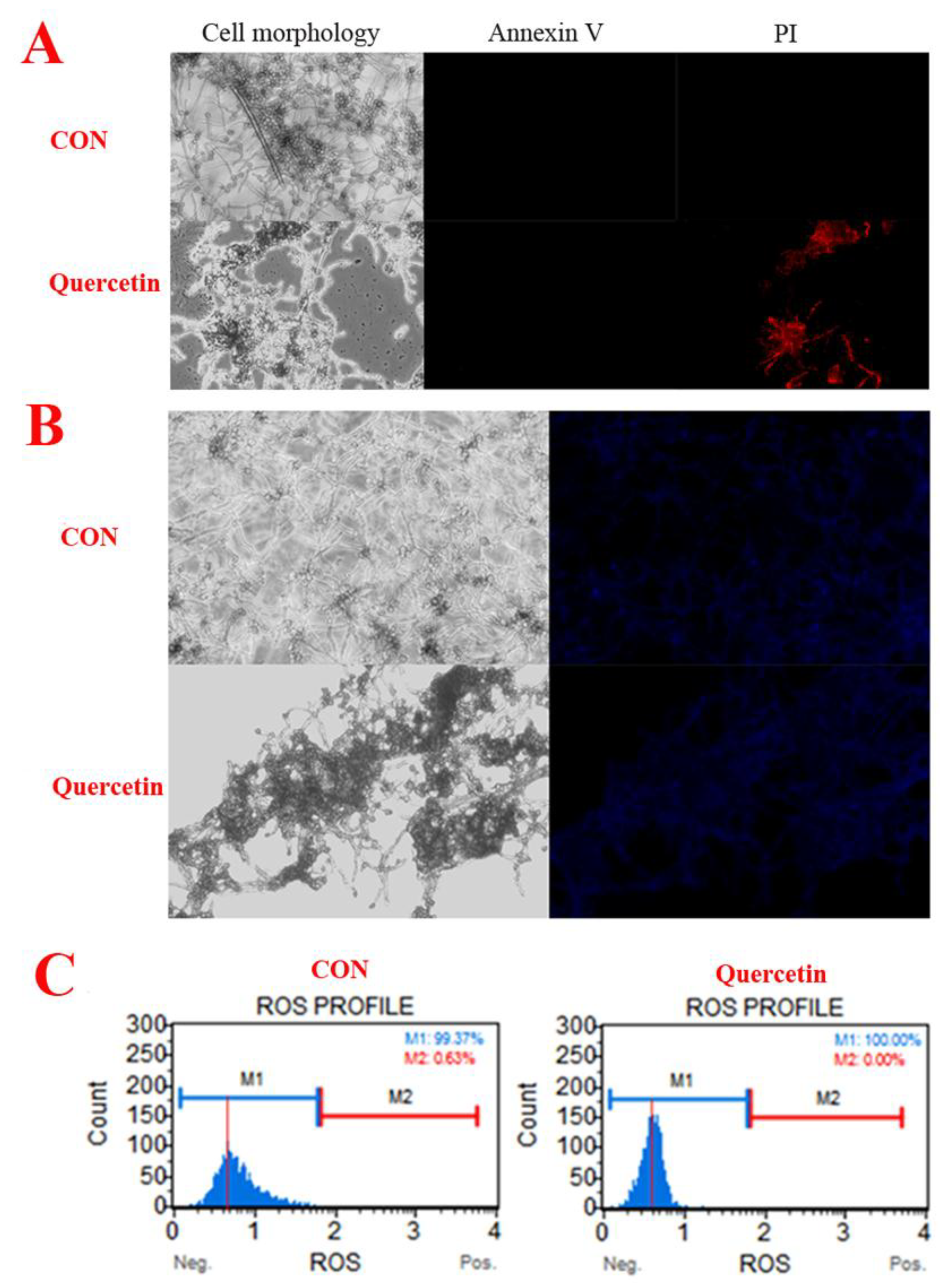
2.2. Morphological Changes of A. flavus
2.3. Cell Apoptosis
2.4. RNA-Seq Data
2.5. Identification and Functional Annotation
2.6. Expression Analysis of Conidial Development- and A. flavus Growth-Related Genes in Response to Quercetin
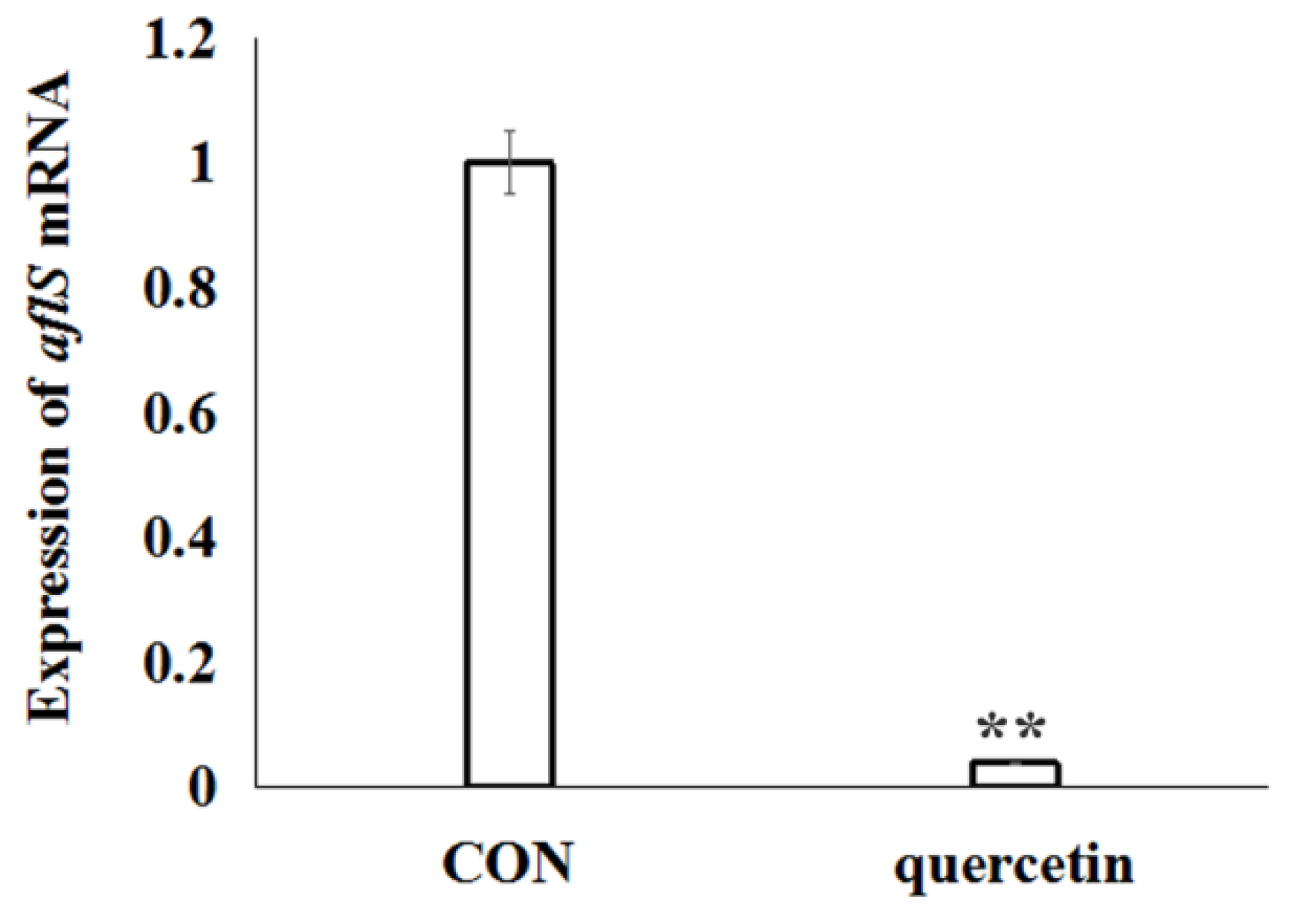
2.7. Expression Analysis of A. flavus AF Biosynthesis-Related Genes in Response to Quercetin
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Fungus Strain and Cultivation
4.3. Anti-Proliferative Activity
4.4. Morphological Changes of A. flavus
4.5. Phosphatidylserine (PS) Externalization
4.6. Hochest 33342
4.7. Measurement of Reactive Oxygen Species
4.8. cDNA Preparation and Illumina Sequencing
4.9. RNA-Seq and Enrichment Analysis of Differentially Expressed Genes
4.10. Validation of RNA-Seq Analysis by Quantitative Real-Time PCR (qRT-PCR)
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cary, J.W.; Han, Z.; Yin, Y.; Lohmarc, J.M.; Shantappa, S.; YHarris-Coward, P. Transcriptome analysis of Aspergillus flavus reveals veA-dependent regulation of secondary metabolite gene clusters, including the novel aflavarin cluster. Eukaryot. Cell. 2015, 14, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Amaike, S.; Kelle, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Alshannaq, A.F.; Gibbons, J.G.; Lee, M.K.; Han, K.H.; Hong, S.B.; Yu, J.H. Controlling aflatoxin contamination and propagation of Aspergillus flavus by a soy-fermenting Aspergillus oryzae strain. Sci. Rep. 2018, 8, 16871. [Google Scholar] [CrossRef] [PubMed]
- Caceres, I.; Snini, S.P.; Puel, O.; Mathieu, F. Streptomyces roseolus, A promising biocontrol agent against Aspergillus flavus, the main Aflatoxin B₁producer. Toxins 2018, 30, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, X.M.; Liu, J.; Pan, F.F.; Shi, D.D.; Wen, Z.G.; Yang, P.L. Quercetin and aconitine synergistically induces the human cervical carcinoma HeLa cell apoptosis via endoplasmic reticulum (ER) stress pathway. PLoS ONE 2018, 13, e0191062. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.Y.; Zhang, H.; Yang, J.; Hu, L.B.; MO, H.Z. Inhibition Effect of Quercetin on Aflatoxin B1 Production. Mod. Food Sci. Technol. 2014, 30, 56–62. [Google Scholar]
- Wang, X.Y.; Wang, Y.Z.; Zhou, Y.G.; Wei, X.L. Farnesol induces apoptosis-like cell death in the pathogenic fungus Aspergillus flavus. Mycologia 2014, 106, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Lei, Y.; Yan, L.Y.; Cheng, K.; Dai, X.F.; Wan, L.Y. Deep sequencing analysis of transcriptomes in Aspergillus flavus in response to resveratrol. BMC Microbiol. 2015, 15, 182. [Google Scholar] [CrossRef]
- Lv, C.; Wang, P.; Ma, L.X.; Zheng, M.M.; Liu, Y.; Xing, F.G. Large-scale comparative analysis of Eugenol-Induced/Repressed genes expression in Aspergillus flavus using RNA-seq. Front. Microbiol. 2018, 9, 1116. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Q.; Zhao, X.X.; Zhi, Q.Q.; Zhao, M.; He, Z.M. Transcriptomic profiling of Aspergillus flavus in response to 5-azacytidine. Fungal Genet. Biol. 2013, 56, 78–86. [Google Scholar] [CrossRef]
- Li, X.M.; Luo, X.G.; Si, C.L.; Wang, N.; Zhou, H.; He, J.F. Antibacterial active compounds from Hypericum ascyron L. induce bacterial cell death through apoptosis pathway. Eur. J. Med. Chem. 2015, 96, 436–444. [Google Scholar] [CrossRef] [PubMed]
- De Souza, R.F.; de Giovani, W.F. Antioxidant properties of complexes of flavonoids with metal ions. Redox Rep. 2004, 9, 97–104. [Google Scholar] [CrossRef]
- Yoon, J.S.; Chae, M.K.; Lee, S.Y.; Lee, E.J. Anti-inflammatory effect of quercetin in a whole orbital tissue culture of Graves’ orbitopathy. Br. J. Ophthalmol. 2012, 96, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Rieger, A.M.; Nelson, K.L.; Konowalchuk, J.D.; Barreda, D.R. Modified annexin V/propidium iodide apoptosis assay for accurate assessment of cell death. J. Vis. Exp. 2011, 50, 2597. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.O.; Tiffany, M.F.; Qualls, C.R.; Key, C.R. The rising incidence of adenocarcinoma relative to squamous cell carcinoma of the uterine cervix in the United States-a 24-year population-based study. Gynecol. Oncol. 2000, 78, 97–105. [Google Scholar] [CrossRef]
- Chang, T.W.; Chen, C.C.; Chen, K.Y.; Su, J.H.; Chang, J.H.; Chang, M.C. Ribosomal phosphoprotein P0 interacts with GCIP and overexpression of P0 is associated with cellular proliferation in breast and liver carcinoma cells. Oncogene 2008, 27, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Akanuma, G.; Nanamiya, H.; Natori, Y.; Yano, K.; Suzuki, S.; Omata, S. Inactivation of ribosomal protein genes in Bacillus subtilis reveals importance of each ribosomal protein for cell proliferation and cell differentiation. J. Bacteriol. 2012, 194, 6282–6291. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Ito, S.; Ihara, T.; Tamura, H.; Tanaka, S.; Ikeda, T.; Kajihara, H. A-tomatine, the major saponin in tomato, induces programmed cell death mediated by reactive oxygen species in the fungal pathogen Fusarium oxysporum. FEBS. Lett. 2007, 581, 3217–3222. [Google Scholar] [CrossRef]
- Shi, M.; Chen, L.; Wang, X.W.; Zhang, T.; Zhao, P.B.; Song, X.Y. Antimicrobial peptaibols from Trichoderma pseudokoningii induce programmed cell death in plant fungal pathogens. Microbiology 2012, 158, 166–175. [Google Scholar] [CrossRef]
- Wang, H.M.; Lei, Y.; Yan, L.Y.; Wan, L.Y.; Ren, X.P.; Chen, S.L. Functional genomic analysis of Aspergillus flavus interacting with resistant and susceptible peanut. Toxins 2016, 8, 46. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K. The Aspergillus parasiticus protein AFLJ interacts with the aflatoxin pathway-specific regulator AFLR. Mol. Genet. Genom. 2003, 268, 711–719. [Google Scholar]
- Calvo, A.M.; Cary, J.W. Association of fungal secondary metabolism and sclerotial biology. Front. Microbiol. 2015, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.S.; Yue, Y.W.; Fu, Y.S.; Fang, Z.; Xu, Z.L.; Ma, G.L. Exploration of the regulatory mechanism of secondary metabolism by comparative transcriptomics in Aspergillus flavus. Front. Microbiol. 2018, 9, 1568–1583. [Google Scholar]
- Benvenuto, M.; Sileri, P.; Rossi, P.; Masuelli, L.; Fantini, M.; Nanni, M. Natural humoral immune response to ribosomal P0 protein in colorectal cancer patients. J. Trans. Med. 2015, 13, 101. [Google Scholar] [CrossRef] [PubMed]
- Remacha, M.; Jimenez-Diaz, A.; Santos, C.; Briones, E.; Zambrano, R.; Rodriguez-Gabriel, M.A. Proteins P1, P2, and P0, components of the eukaryotic ribosome stalk. New structural and functional aspects. Biochem. Cell Biol. 1995, 73, 959–968. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Zhang, B.C.; Zhang, A.R.; Wu, T.T.; Liu, J.; Yu, L.F.; Wang, W.-X.; Gao, J.-F.; Fang, D.-C.; Rao, Z.-G. Co-transduction of ribosomal protein L23 enhances the therapeutic efficacy of adenoviral-mediated p53 gene transfer in human gastric cancer. Oncol. Rep. 2013, 30, 1989–1995. [Google Scholar] [CrossRef]
- Qi, Y.K.; Li, X.; Chang, C.K.; Xu, F.; He, Q.; Zhao, Y.S. Ribosomal protein L23 negatively regulates cellular apoptosis via the RPL23/Miz-1/c-Myc circuit in higher-risk myelodysplastic syndrome. Sci. Rep. 2016, 7, 2323. [Google Scholar] [CrossRef]
- Pidutti, P.; Federici, F.; Brandi, J.; Manna, L.; Rizzi, E.; Marini, U. Purification and characterization of ribosomal proteins L27 and L30 having antimicrobial activity produced by the Lactobacillus salivarius SGL03. J. Appl. Microbiol. 2017, 124, 398–407. [Google Scholar] [CrossRef]
- Smolock, E.M.; Korshunov, V.A.; Glazko, G.; Qiu, X.; Gerloff, J.; Berk, B.C. Ribosomal Protein L17, RpL17, is an Inhibitor of Vascular Smooth Muscle Growth and Carotid Intima Formation. Circulation 2012, 126, 2418–2427. [Google Scholar] [CrossRef]
- Mina, H.; Kim, H.R.; Kim, I. Ribosomal protein L19 overexpression activates the unfolded protein response and sensitizes MCF7 breast cancer cells to endoplasmic reticulum stress-induced cell death. Biochem. Biophs. Res. Commun. 2014, 450, 673–678. [Google Scholar]
- Bee, A.; Brewer, D.; Beesley, C.; Dodson, A.; Forootan, S.; Dickinson, T. SiRNA knockdown of ribosomal protein gene RPL19 abrogates the aggressive phenotype of human prostate cancer. PLoS ONE 2011, 6, e22672. [Google Scholar] [CrossRef]
- Cao, B.; Fang, Z.L.; Liao, P.; Zhou, X.; Xiong, J.P.; Zeng, S. Cancer-mutated ribosome protein L22 (RPL22/eL22) suppresses cancer cell survival by blocking p53-MDM2 circuit. Oncotarget 2017, 8, 90651–90661. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.J.; Li, J.; Jia, Y.X.; Wu, J.J.; Yuan, L.; Li, M.J. Silencing of ribosomal protein L34 (RPL34) inhibits the proliferation and invasion of esophageal cancer cells. Oncol. Res. 2017, 25, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Ding, L.J.; Wei, Z.T.; Zhang, Y.D.; Li, Y.; Luo, Q.H. Ribosomal protein L34 promotes the proliferation, invasion and metastasis of pancreatic cancer cells. Oncotarget 2016, 7, 85259–85272. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Gou, Y.; Wang, Q.; Jin, H.; Cui, L.; Zhang, Y. Downregulation of RPL6 by siRNA inhibits proliferation and cell cycle progression of human gastric cancer cell lines. PLoS ONE 2011, 6, e26401. [Google Scholar] [CrossRef]
- Guo, X.; Shi, Y.; Gou, Y.; Li, J.; Han, S.; Zhang, Y. Human ribosomal protein S13 promotes gastric cancer growth through down-regulating p27(Kip1). J. Cell Mol. Med. 2011, 15, 296–306. [Google Scholar] [CrossRef]
- Li, Y.; Dong, W.; Shi, Y.; Deng, F.; Chen, X.; Wan, C.; Zhou, M.; Zhao, L.; Fu, Z.F.; Peng, G. Rabies virus phosphoprotein interacts with ribosomal protein L9 and affects rabies virus replication. Virology 2016, 488, 216–224. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Word, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 7, 621–628. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Length | Means-CK | Means-QT | log2(Fold Change) | Probability | Description |
|---|---|---|---|---|---|---|
| AFLA_020210 | 1362 | 370.1433333 | 64.65666667 | −2.517213007 | 0.885380738 | sexual development transcription factor NsdD |
| AFLA_048650 | 2100 | 156.1633333 | 38.95 | −2.00336052 | 0.844491613 | sexual development transcription factor SteA |
| AFLA_018540 | 1068 | 67.02666667 | 14.44666667 | −2.213998536 | 0.81499661 | G protein complex alpha subunit GpaB |
| AFLA_046990 | 2370 | 200.3033333 | 75.34333333 | −1.410634662 | 0.8049291 | APSES transcription factor StuA |
| AFLA_083110 | 255 | 766.0933333 | 24.60333333 | −4.960594472 | 0.950709 | conidiation-specific protein Con-10 |
| AFLA_044790 | 309 | 525.2933333 | 124.2433333 | −2.079954833 | 0.873234869 | conidiation-specific family protein |
| NO. | Gene Name | Means-CK | Means-QT | log2(Fold Change) | Up-Down-Regulation (QT/CK) | Description |
|---|---|---|---|---|---|---|
| 1 | AFLA_047870 | 2358.77 | 2.84 | −1.920818 | Down | 40S ribosomal protein S1 |
| 2 | AFLA_034340 | 976.7166667 | 6.739345 | −2.16111509 | Down | 40S ribosomal protein S10b |
| 3 | AFLA_050800 | 723.8966667 | 1.4697534 | −1.69897 | Down | 40S ribosomal protein S12 |
| 4 | AFLA_075030 | 802.26 | 9.62712 | −1.92081875 | Down | 40S ribosomal protein S13 |
| 5 | AFLA_044170 | 765.6133333 | 3.8280665 | −2.30102999 | Down | 40S ribosomal protein S14 |
| 6 | AFLA_044530 | 2185.433333 | 34.966928 | −1.795880017 | Down | 40S ribosomal protein S15 |
| 7 | AFLA_044110 | 763.1666667 | 9.9211671 | −1.886056647 | Down | 40S ribosomal protein S16 |
| 8 | AFLA_125890 | 2183.286667 | 43.66574 | −1.69897 | Down | 40S ribosomal protein S17 |
| 9 | AFLA_009000 | 1360.496667 | 34.012425 | −1.602059991 | Down | 40S ribosomal protein S18 |
| 10 | AFLA_050650 | 898.7666667 | 20.6716341 | −1.638272164 | Down | 40S ribosomal protein S19 |
| 11 | AFLA_117990 | 651.8966667 | 23.4682812 | −1.443697016 | Down | 40S ribosomal protein S2 |
| 12 | AFLA_043150 | 1318.843333 | 44.840662 | −1.468521082 | Down | 40S ribosomal protein S20 |
| 13 | AFLA_092120 | 8890.06 | 515.62348 | −1.236572001 | Down | 40S ribosomal protein S21 |
| 14 | AFLA_083740 | 680.39 | 6.8039 | −2.067506415 | Down | 40S ribosomal protein S22 |
| 15 | AFLA_021730 | 846.6866667 | 3.3867468 | −2.399868277 | Down | 40S ribosomal protein S23 |
| 16 | AFLA_071320 | 1901.14 | 7.60456 | −2.397940008 | Down | 40S ribosomal protein S24 |
| 17 | AFLA_083050 | 5390.12 | 29.64566 | −2.259637311 | Down | 40S ribosomal protein S25 |
| 17 | AFLA_127800 | 4272.09 | 26.059749 | −2.214670164 | Down | 40S ribosomal protein S26 |
| 18 | AFLA_083470 | 140.9033333 | 2.5362594 | −1.744727494 | Down | 40S ribosomal protein S28 |
| 19 | AFLA_101020 | 1328.043333 | 55.77806 | −1.37675071 | Down | 40S ribosomal protein S4 |
| 20 | AFLA_084620 | 1535.576667 | 16.89347 | −1.958607314 | Down | 40S ribosomal protein S5 |
| 21 | AFLA_026830 | 2783.25 | 125.24625 | −1.346787486 | Down | 40S ribosomal protein S6 |
| 22 | AFLA_029020 | 3209.673333 | 8954.783333 | −1.48148606 | Down | 40S ribosomal protein S8 |
| 23 | AFLA_101160 | 832.2533333 | 1.747731999 | −2.677780753 | Down | 40S ribosomal protein S9 |
| 24 | AFLA_030140 | 259.6833333 | 929.63 | 1.839903203 | Down | 60S acidic ribosomal protein P0 |
| 25 | AFLA_127860 | 264.4766667 | 10.05011333 | −1.420216403 | Down | 60S acidic ribosomal protein P1 |
| 26 | AFLA_044520 | 984.8466667 | 2.659086 | −2.568636235 | Down | 60S acidic ribosomal protein P2 |
| 27 | AFLA_068000 | 765.0433333 | 1.606590999 | −2.677780705 | Down | 60S ribosomal protein L1 |
| 28 | AFLA_112090 | 1100.716667 | 7.1546583 | −2.187086643 | Down | 60S ribosomal protein L11 |
| 29 | AFLA_080140 | 1298.606667 | 37.659593 | −1.537602002 | Down | 60S ribosomal protein L12 |
| 30 | AFLA_115110 | 943.4 | 9.24532 | −2.008773924 | Down | 60S ribosomal protein L13 |
| 31 | AFLA_056250 | 879.56 | 22.86856 | −1.585026652 | Down | 60S ribosomal protein L14 |
| 32 | AFLA_029260 | 633.1733333 | 16.14591999 | −1.593459819 | Down | 60S ribosomal protein L15, putative |
| 33 | AFLA_050000 | 1516.85 | 32.612275 | −1.66756154 | Down | 60S ribosomal protein L16 |
| 34 | AFLA_041990 | 1251.423333 | 4533.676667 | 1.857111595 | Up | 60S ribosomal protein L17 |
| 35 | AFLA_047440 | 2671.72 | 35.266704 | −1.879426068 | Down | 60S ribosomal protein L18 |
| 36 | AFLA_046970 | 2952.823333 | 8912.88 | 1.593796639 | Up | 60S ribosomal protein L19 |
| 37 | AFLA_048810 | 2272.563333 | 163.6245599 | −1.136677139 | Down | 60S ribosomal protein L2 |
| 38 | AFLA_029450 | 670.29 | 7.37319 | −1.958607314 | Down | 60S ribosomal protein L20 |
| 39 | AFLA_101150 | 2427.973333 | 41.2755466 | −1.769551078 | Down | 60S ribosomal protein L21 |
| 40 | AFLA_079880 | 233.58 | 1312.223333 | 2.490024624 | Up | 60S ribosomal protein L22 |
| 41 | AFLA_092370 | 1086.093333 | 13.25033866 | −1.913640169 | Down | 60S ribosomal protein L23 |
| 42 | AFLA_092370 | 1387.563333 | 70.76572998 | −1.292429823 | Down | 60S ribosomal protein L24 |
| 43 | AFLA_048140 | 2383.7 | 165.66715 | −1.155522824 | Down | 60S ribosomal protein L25 |
| 44 | AFLA_110470 | 188.4966667 | 9.481382335 | −1.298432014 | Down | 60S ribosomal protein L25, putative |
| 45 | AFLA_060150 | 2232.186667 | 13.39312 | −2.221848749 | Down | 60S ribosomal protein L26 |
| 46 | AFLA_127220 | 862.0833333 | 13.44849999 | −1.806875401 | Down | 60S ribosomal protein L27 |
| 47 | AFLA_054760 | 1163.723333 | 8.0296909 | −2.161150909 | Down | 60S ribosomal protein L28 |
| 48 | AFLA_103770 | 1127.98 | 10.039022 | −2.050609993 | Down | 60S ribosomal protein L28 |
| 49 | AFLA_134740 | 1447.586667 | 50.66553335 | −1.455931955 | Down | 60S ribosomal protein L3 |
| 50 | AFLA_045790 | 2011.47 | 43.246605 | −1.66756154 | Down | 60S ribosomal protein L30 |
| 51 | AFLA_045790 | 1371.076667 | 4046.166667 | −1.561246503 | Down | 60S ribosomal protein L31 |
| 52 | AFLA_003480 | 11,949.94667 | 33,519.81333 | 1.488009936 | Up | 60S ribosomal protein L32 |
| 53 | AFLA_086630 | 1485.253333 | 2.673455999 | −2.744727494 | Down | 60S ribosomal protein L33 |
| 54 | AFLA_086630 | 2061.426667 | 18.55284 | −2.04575749 | Down | 60S ribosomal protein L34 |
| 55 | AFLA_086630 | 1270.083333 | 5.715374999 | −2.346787486 | Down | 60S ribosomal protein L35 |
| 56 | AFLA_112390 | 832.9033333 | 4.164516667 | −2.301029995 | Down | 60S ribosomal protein L36 |
| 57 | AFLA_112390 | 3332.77 | 39.99324 | −1.920818753 | Down | 60S ribosomal protein L37 |
| 58 | AFLA_112390 | 1112.88 | 14.244864 | −1.89279003 | Down | 60S ribosomal protein L38 |
| 59 | AFLA_112390 | 1971.986667 | 29.77699867 | −1.821023052 | Down | 60S ribosomal protein L42 |
| 60 | AFLA_112390 | 3816.61 | 106.483419 | −1.554395796 | Down | 60S ribosomal protein L43 |
| 61 | AFLA_018700 | 1022.05 | 17.885875 | −1.732828271 | Down | 60S ribosomal protein L5 |
| 62 | AFLA_068420 | 3303.43 | 87.540895 | −1.576754126 | Down | 60S ribosomal protein L6 |
| 63 | AFLA_041710 | 1208.193333 | 5.255640998 | −2.361510743 | Down | 60S ribosomal protein L7 |
| 64 | AFLA_033980 | 1874.96 | 88.12312 | −1.327902142 | Down | 60S ribosomal protein L8 |
| 65 | AFLA_088370 | 686.0866667 | 3323.653333 | 2.276307178 | Up | 60S ribosomal protein L9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.-M.; Li, Z.-Y.; Wang, Y.-D.; Wang, J.-Q.; Yang, P.-L. Quercetin Inhibits the Proliferation and Aflatoxins Biosynthesis of Aspergillus flavus. Toxins 2019, 11, 154. https://doi.org/10.3390/toxins11030154
Li X-M, Li Z-Y, Wang Y-D, Wang J-Q, Yang P-L. Quercetin Inhibits the Proliferation and Aflatoxins Biosynthesis of Aspergillus flavus. Toxins. 2019; 11(3):154. https://doi.org/10.3390/toxins11030154
Chicago/Turabian StyleLi, Xiu-Mei, Zhong-Yuan Li, Ya-Dong Wang, Jin-Quan Wang, and Pei-Long Yang. 2019. "Quercetin Inhibits the Proliferation and Aflatoxins Biosynthesis of Aspergillus flavus" Toxins 11, no. 3: 154. https://doi.org/10.3390/toxins11030154
APA StyleLi, X.-M., Li, Z.-Y., Wang, Y.-D., Wang, J.-Q., & Yang, P.-L. (2019). Quercetin Inhibits the Proliferation and Aflatoxins Biosynthesis of Aspergillus flavus. Toxins, 11(3), 154. https://doi.org/10.3390/toxins11030154