Modulation of Mucin (MUC2, MUC5AC and MUC5B) mRNA Expression and Protein Production and Secretion in Caco-2/HT29-MTX Co-Cultures Following Exposure to Individual and Combined Aflatoxin M1 and Ochratoxin A



Abstract
1. Introduction
2. Results
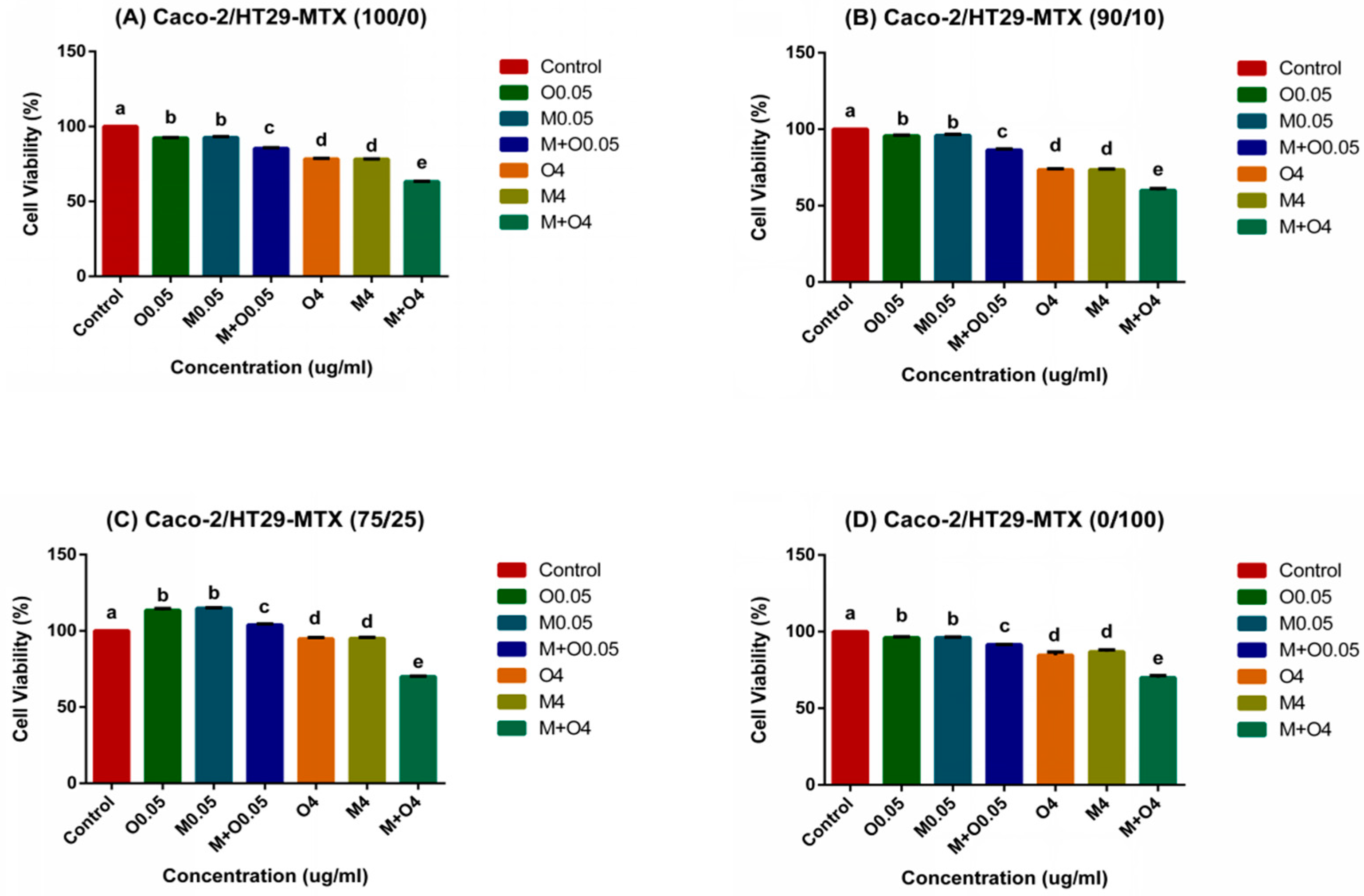
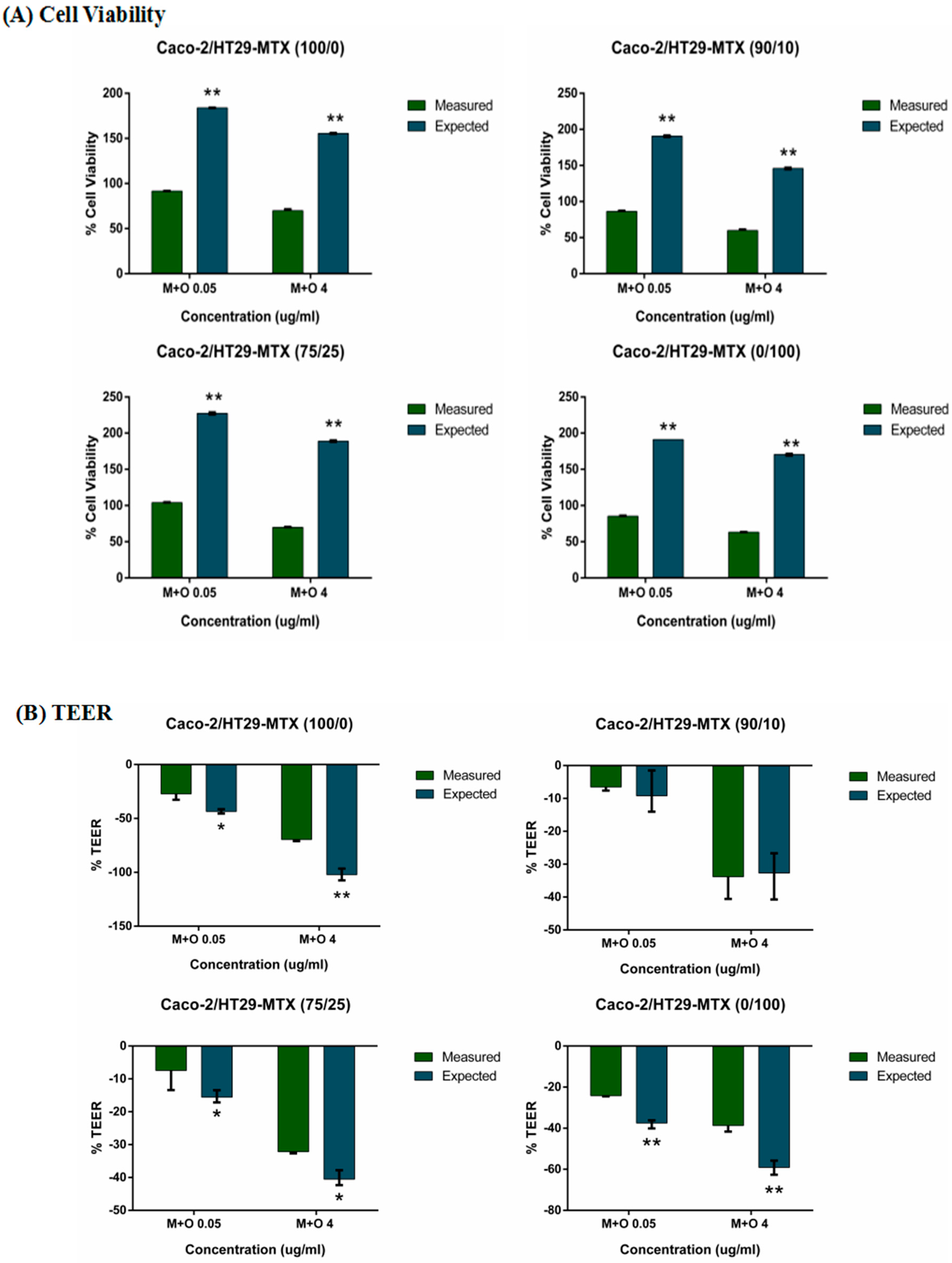
2.1. Effects of AFM1 and OTA Individually Or in Combination on Caco-2/HT29-MTX Cell Viability (100/0, 90/10, 75/25, and 0/100)
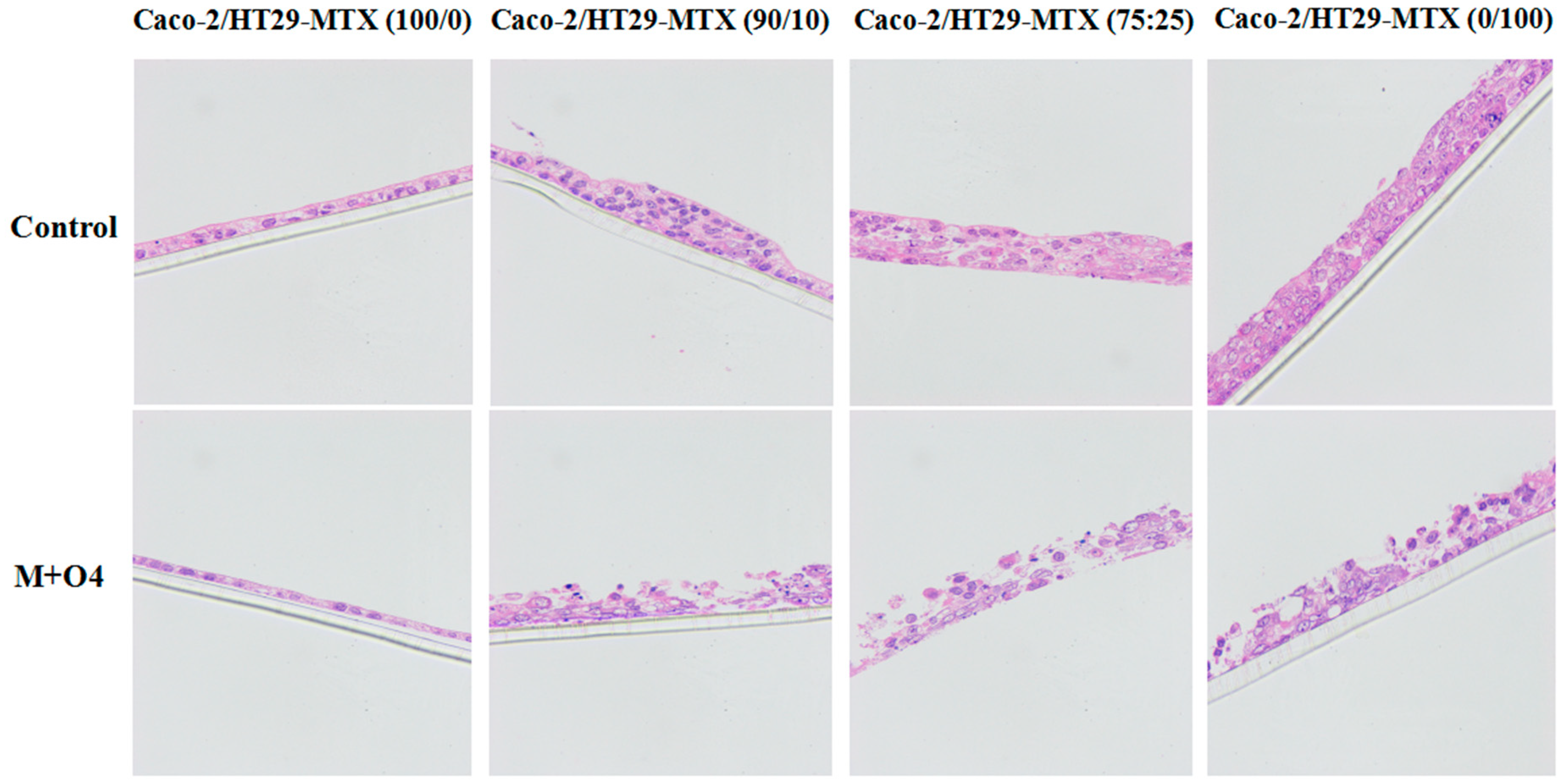
2.2. Effect of AFM1 and OTA in Combination on Caco-2/HT29-MTX Cell Layer Structures (100/0, 90/10, 75/25, and 0/100)
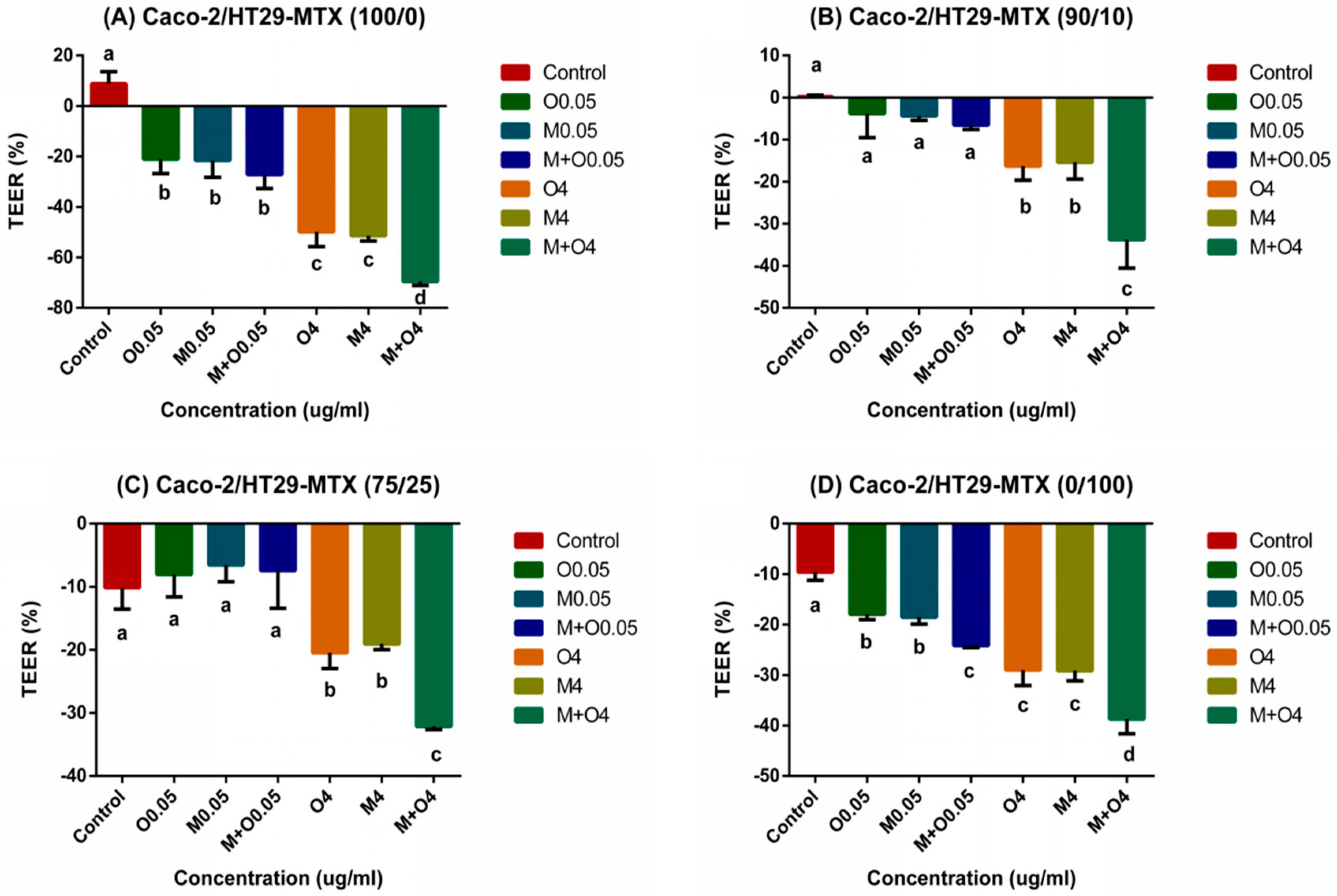
2.3. Effects of AFM1 and OTA Individually Or Collectively on Caco-2/HT29-MTX TEER Values (100/0, 90/10, 75/25, and 0/100)
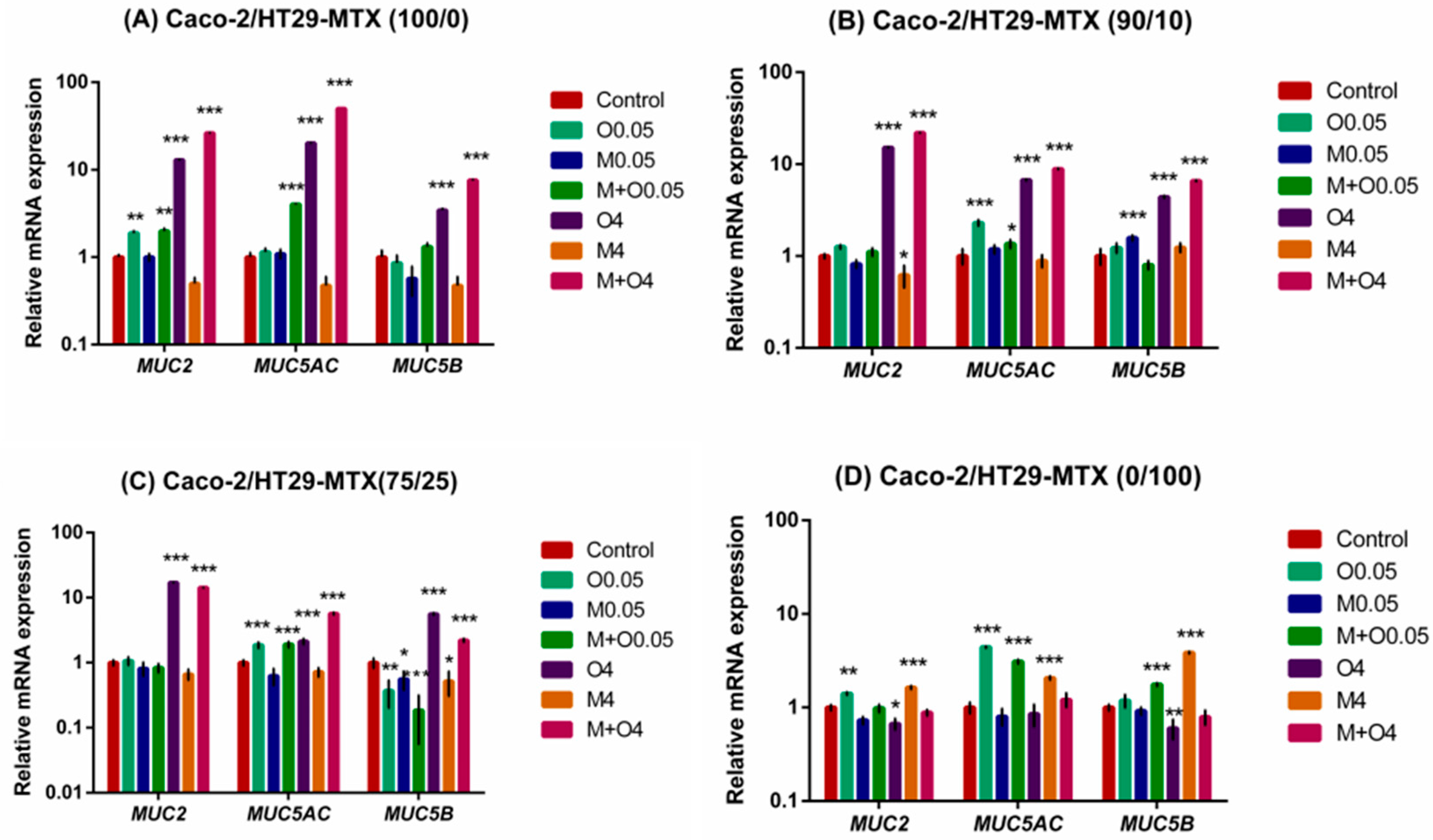
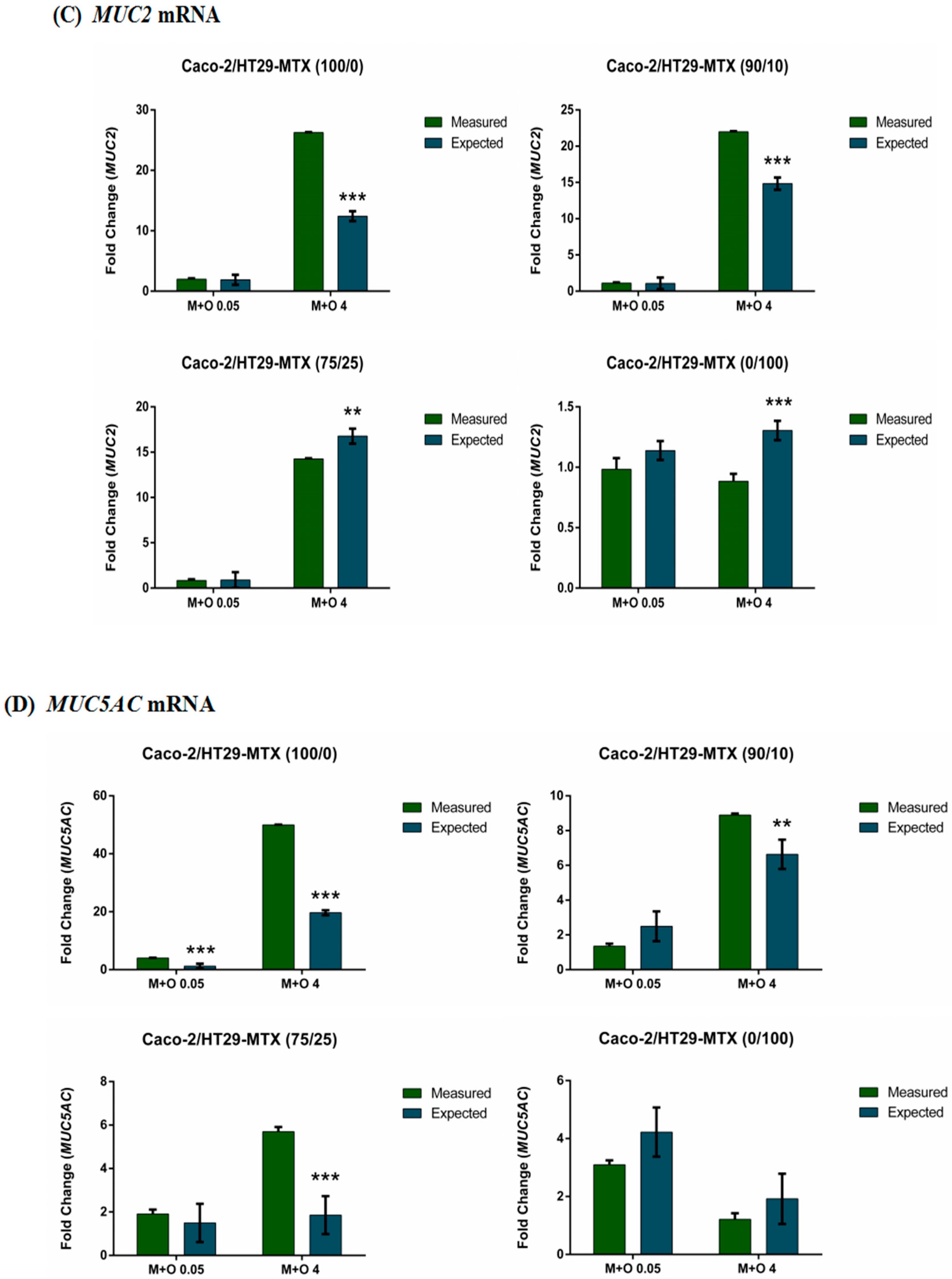
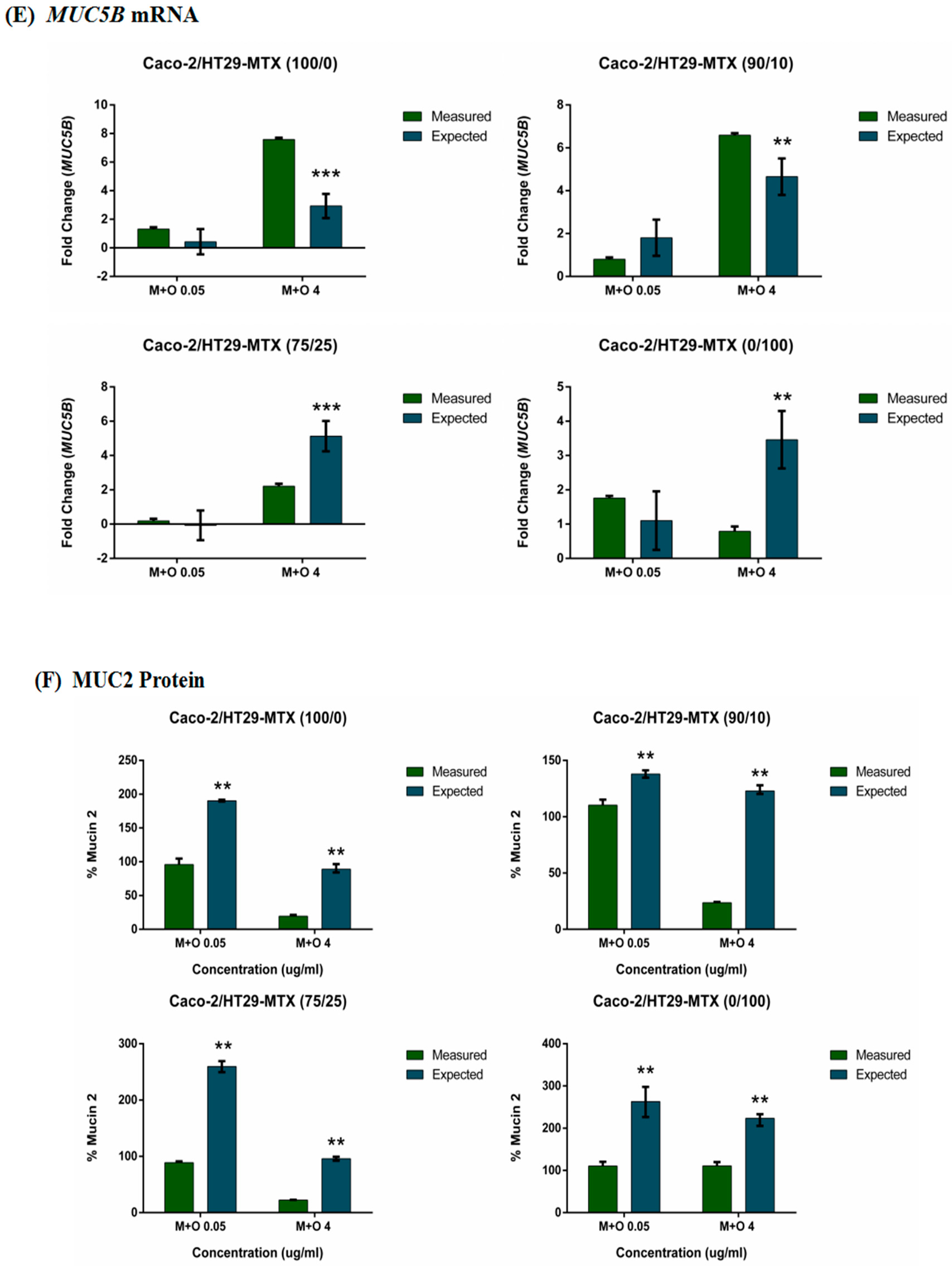
2.4. Effects of AFM1 and OTA Individually Or in Combination on Mucin Gene Expression of Caco-2/HT29-MTX Cultures (100/0, 90/10, 75/25, and 0/100)
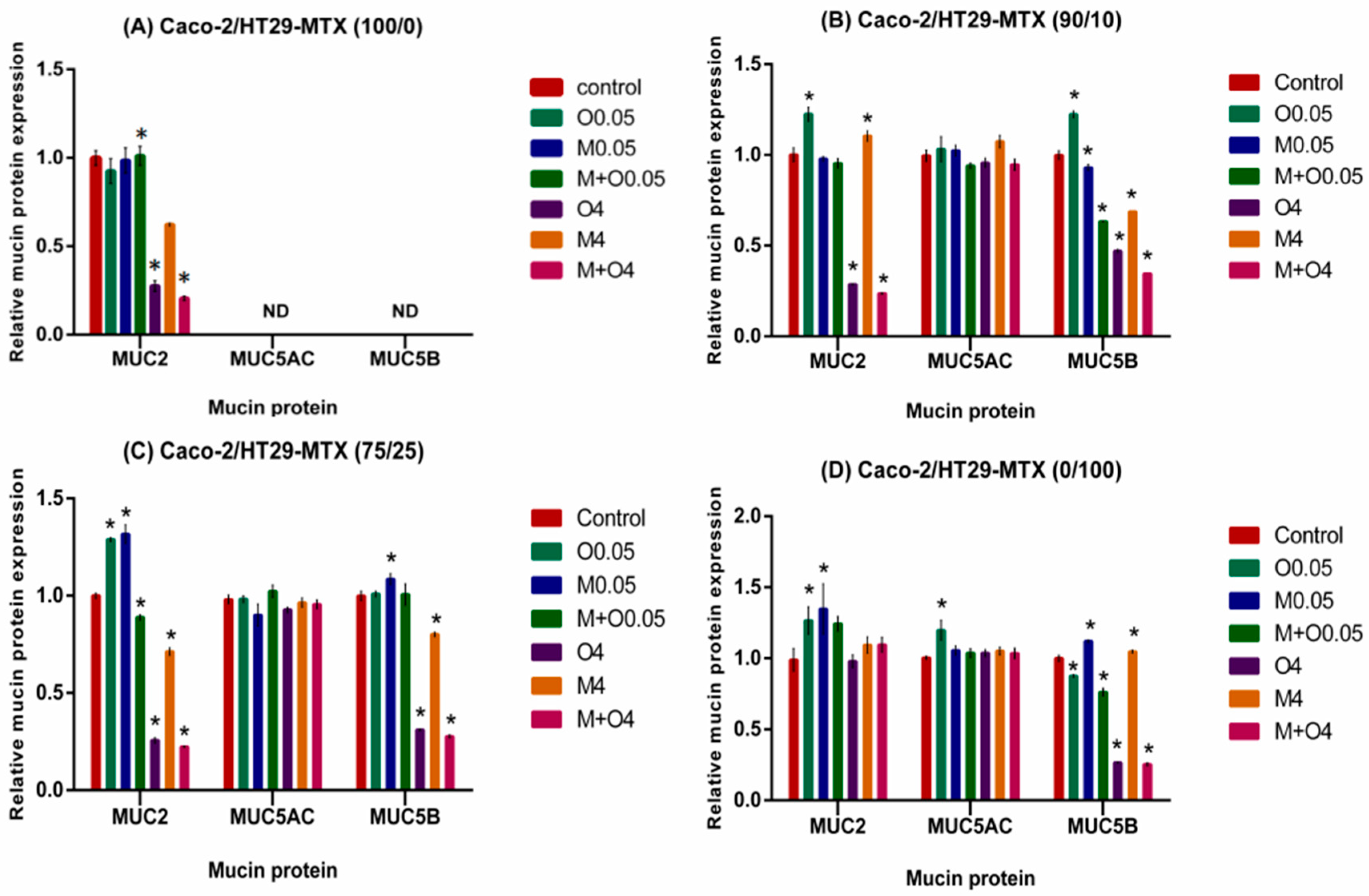
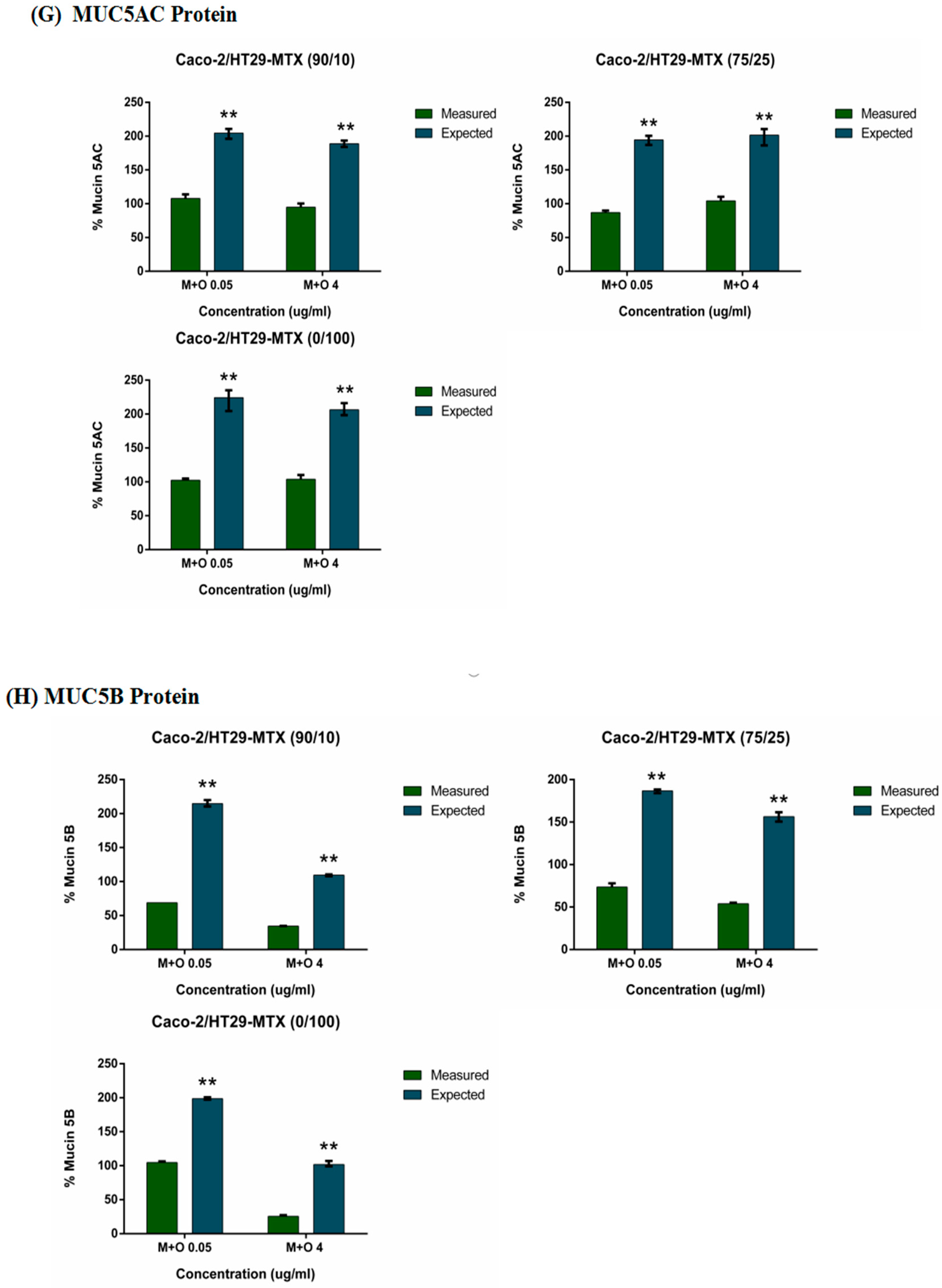
2.5. Effects of AFM1 and OTA Individually Or in Combination on Mucin Protein Abundance of Caco-2/HT29-MTX Cultures (100/0, 90/10, 75/25, and 0/100)
2.6. The Interactive Effects of the Combination of AFM1 and OTA on Caco-2/HT29-MTX Single Cultures and Co-Cultures (100/0, 90/10, 75/25, and 0/100)
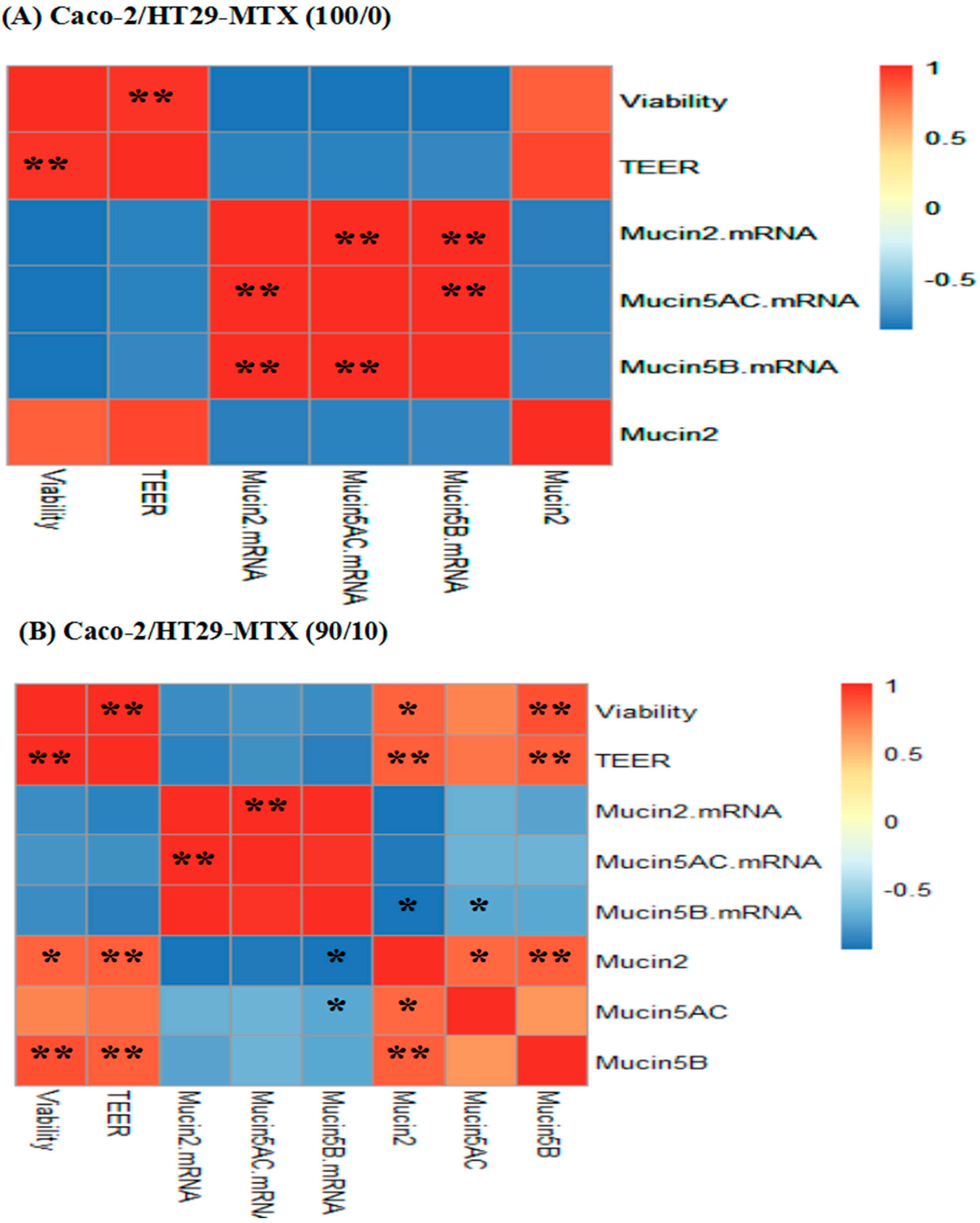
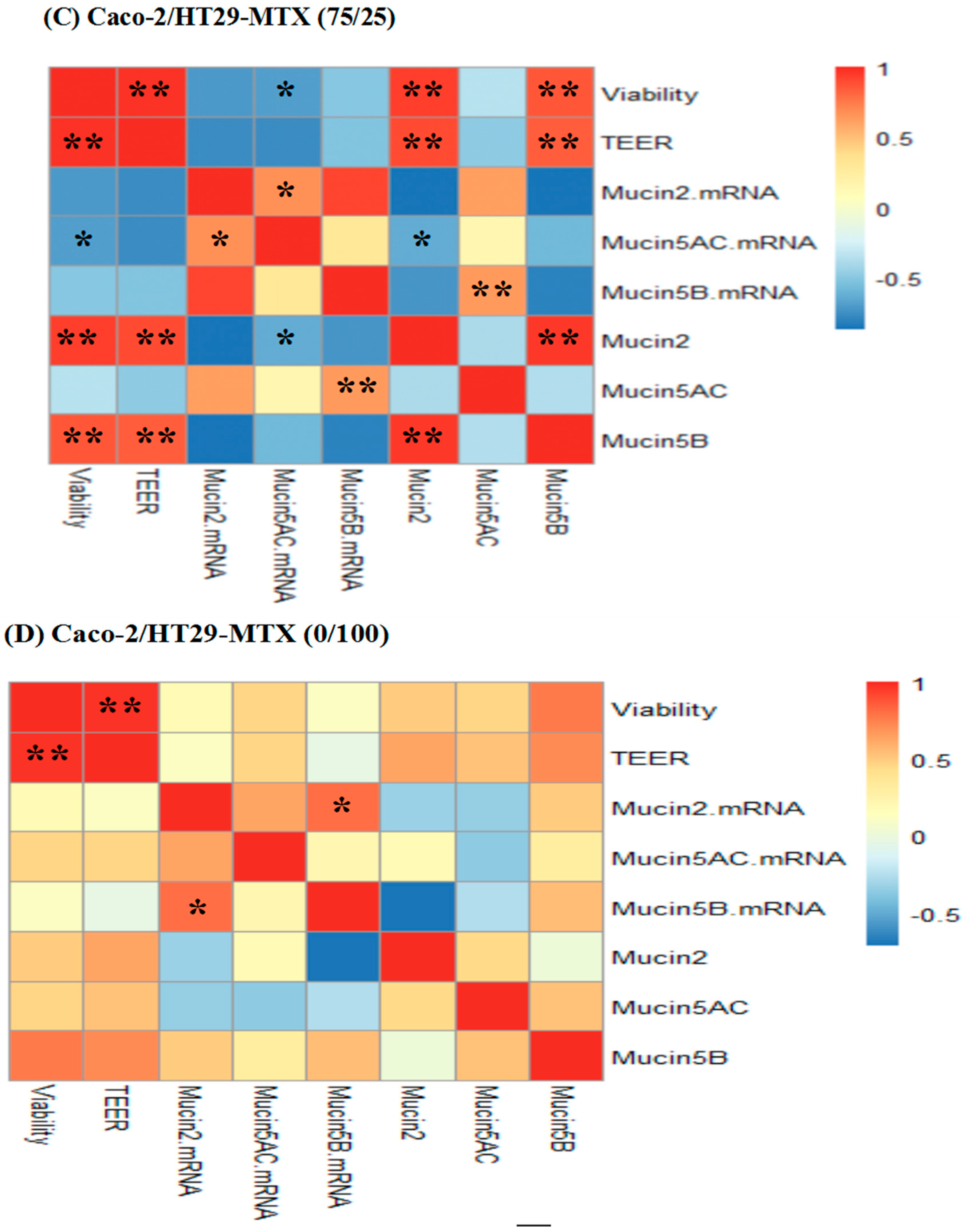
2.7. Correlations between Cell Viability, TEER, Mucin mRNA Expression and Mucin Protein Levels
3. Discussion
4. Materials and Methods
4.1. Mycotoxin Treatment
4.2. Cell Lines and Culture Conditions
4.3. Cell Viability Assay
4.4. Cell Layer Staining
4.5. TEER Measurement
4.6. Quantification of Mucin Gene Expression
4.7. Mucin Protein Abundance
4.8. Analysis of Interactions and Correlations
- Additive effects were defined as the measured values for endpoints that were not significantly above or below the expected values (p > 0.05).
- Synergistic effects were defined as measured values that were significantly lower than the expected values.
- Antagonistic effects were defined as measured values that were significantly higher than the expected values.
- Correlations among cell viability, TEER values, and the levels of mucin mRNA and protein in Caco-2/HT29-MTX co-cultures treated with AFM1 and OTA individually or collectively were assessed by Spearman’s correlations (nonparametric) and R v3.5.2. (TUNA Team, Tsinghua University, Beijing, China) was used for drawing.
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Raiola, A.; Tenore, G.C.; Manyes, L.; Meca, G.; Ritieni, A. Risk analysis of main mycotoxins occurring in food for children: An overview. Food Chem. Toxicol. 2015, 84, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Bouhet, S.; Oswald, I.P. The effects of mycotoxins, fungal food contaminants, on the intestinal epithelial cell-derived innate immune response. Vet. Immunol. Immunopathol. 2005, 108, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.W.; Subrahmanyam, S.; Piletsky, S.A. Analytical methods for determination of mycotoxins: A review. Anal. Chim. Acta 2009, 632, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Arnich, N.; Sirot, V.; Riviere, G.; Jean, J.; Noel, L.; Guerin, T.; Leblanc, J.C. Dietary exposure to trace elements and health risk assessment in the 2nd french total diet study. Food Chem. Toxicol. 2012, 50, 2432–2449. [Google Scholar] [CrossRef] [PubMed]
- Flores-Flores, M.E.; Lizarraga, E.; Cerain, A.L.D.; González-Peñas, E. Presence of mycotoxins in animal milk: A review. Food Control 2015, 53, 163–176. [Google Scholar] [CrossRef]
- Sakin, F.; Tekeli, İ.O.; Yipel, M.; Kürekci, C. Occurrence and health risk assessment of aflatoxins and ochratoxin a in sürk, a Turkish dairy food, as studied by HPLC. Food Control 2018, 90, 317–323. [Google Scholar] [CrossRef]
- Huang, L.C.; Zheng, N.; Zheng, B.Q.; Wen, F.; Cheng, J.B.; Han, R.W.; Xu, X.M.; Li, S.L.; Wang, J.Q. Simultaneous determination of aflatoxin m1, ochratoxin a, zearalenone and α-zearalenol in milk by uhplc-ms/ms. Food Chem. 2014, 146, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Wang, J.Q.; Han, R.W.; Zhen, Y.P.; Xu, X.M.; Sun, P. Survey of aflatoxin m1 in raw milk in the five provinces of china. Food Addit. Contam. 2013, 6, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Aslam, N.; Wynn, P. Aflatoxin contamination of the milk supply: A pakistan perspective. Agriculture 2015, 5, 1172–1182. [Google Scholar] [CrossRef]
- Gao, Y.N.; Wang, J.Q.; Li, S.L.; Zhang, Y.D.; Zheng, N. Aflatoxin m1 cytotoxicity against human intestinal caco-2 cells is enhanced in the presence of other mycotoxins. Food Chem. Toxicol. 2016, 96, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Akbari, P.; Braber, S.; Varasteh, S.; Alizadeh, A.; Garssen, J.; Fink-Gremmels, J. The intestinal barrier as an emerging target in the toxicological assessment of mycotoxins. Arch. Toxicol. 2017, 91, 1007–1029. [Google Scholar] [CrossRef] [PubMed]
- Boudra, H.; Bars, P.L.; Bars, J.L. Thermostability of ochratoxin a in wheat under two moisture conditions. Appl. Environ. Microbiol. 1995, 61, 1156–1158. [Google Scholar] [PubMed]
- Solcan, C.; Pavel, G.; Floristean, V.C.; Chiriac, I.S.; BG, Ş.; Solcan, G. Effect of ochratoxin a on the intestinal mucosa and mucosa-associated lymphoid tissues in broiler chickens. Acta Vet. Hung. 2015, 63, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Jbj, K.; Mercier-Bonin, M.; Eutamène, H.; Theodorou, V. Mucus organisation is shaped by colonic content; a new view. Sci. Rep. 2017, 7, 8527. [Google Scholar]
- Tarabova, L.; Makova, Z.; Piesova, E.; Szaboova, R.; Faixova, Z. Intestinal mucus layer and mucins (a review). Folia Vet. 2016, 60, 21–25. [Google Scholar] [CrossRef]
- Rose, M.C.; Voynow, J.A. Respiratory tract mucin genes and mucin glycoproteins in health and disease. Physiol. Rev. 2006, 86, 245–278. [Google Scholar] [CrossRef] [PubMed]
- Kufe, D.W. Mucins in cancer: Function, prognosis and therapy. Nat. Rev. Cancer 2009, 9, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Ahmed Adam, M.A.; Tabana, Y.M.; Musa, K.B.; Sandai, D.A. Effects of different mycotoxins on humans, cell genome and their involvement in cancer (review). Oncol. Rep. 2017, 37, 1321–1336. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Applegate, T. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; Garbetta, A.; D’Antuono, I.; Cardinali, A.; Martino, N.A.; Debellis, L.; Visconti, A. Toxic mechanisms induced by fumonisin b1 mycotoxin on human intestinal cell line. Arch. Environ. Contam. Toxicol. 2014, 67, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Wikman-Larhed, A.; Artursson, P. Co-cultures of human intestinal goblet (ht29-h) and absorptive (caco-2) cells for studies of drug and peptide absorption. Eur. J. Pharm. Sci. 1995, 3, 171–183. [Google Scholar] [CrossRef]
- Mahler, G.J.; Shuler, M.L.; Glahn, R.P. Characterization of caco-2 and ht29-mtx cocultures in an in vitro digestion/cell culture model used to predict iron bioavailability. J. Nutr. Biochem. 2009, 20, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.Y.; Cedergreen, N.; Yiannikouris, A.; Swamy, H.V.; Karrow, N.A. Assessing interactions of binary mixtures of penicillium mycotoxins (pms) by using a bovine macrophage cell line (bomacs). Toxicol. Appl. Pharmacol. 2017, 318, 33. [Google Scholar] [CrossRef] [PubMed]
- Szabo, A.; Szabo-Fodor, J.; Febel, H.; Mezes, M.; Balogh, K.; Bazar, G.; Kocso, D.; Ali, O.; Kovacs, M. Individual and combined effects of fumonisin b(1), deoxynivalenol and zearalenone on the hepatic and renal membrane lipid integrity of rats. Toxins 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Hilgendorf, C.; Spahn-Langguth, H.; Regardh, C.G.; Lipka, E.; Amidon, G.L.; Langguth, P. Caco-2 versus caco-2/ht29-mtx co-cultured cell lines: Permeabilities via diffusion, inside- and outside-directed carrier-mediated transport. J. Pharm. Sci. 2015, 89, 63–75. [Google Scholar] [CrossRef]
- Xu, Q.; Fan, H.; Yu, W.; Hui, H.; Wu, J. Transport study of egg-derived antihypertensive peptides (lkp and iqw) using caco-2 and ht29 coculture monolayers. J. Agric. Food. Chem. 2017, 65, 7406–7414. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.Y.; Allen, K.J.; Turner, P.C.; El-Nezami, H. Modulation of mucin mrna (muc5ac and muc5b) expression and protein production and secretion in caco-2/ht29-mtx co-cultures following exposure to individual and combined fusarium mycotoxins. Toxicol. Sci. 2014, 139, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.C.; Madec, S.; Troadec, S.; Coton, E.; Hymery, N. Effects of fusariotoxin co-exposure on thp-1 human immune cells. Cell Biol. Toxicol. 2017, 34, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Janssens, G.P.; De Baere, S.; Mountzouris, K.C.; Su, S.; Wong, E.A.; De Meulenaer, B.; et al. Mycotoxins deoxynivalenol and fumonisins alter the extrinsic component of intestinal barrier in broiler chickens. J. Agric. Food. Chem. 2015, 63, 10846–10855. [Google Scholar] [CrossRef] [PubMed]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing protein abundance and mrna expression levels on a genomic scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.; Perrais, M.; Desseyn, J.L.; Aubert, J.P.; Pigny, P.; Van Seuningen, I. Epigenetic regulation (DNA methylation, histone modifications) of the 11p15 mucin genes (muc2, muc5ac, muc5b, muc6) in epithelial cancer cells. Oncogene 2007, 26, 6566–6576. [Google Scholar] [CrossRef] [PubMed]
- Barnett, A.M.; Roy, N.C.; McNabb, W.C.; Cookson, A.L. Effect of a semi-purified oligosaccharide-enriched fraction from caprine milk on barrier integrity and mucin production of co-culture models of the small and large intestinal epithelium. Nutrients 2016, 8, 267. [Google Scholar] [CrossRef] [PubMed]
- Lesuffleur, T.; Porchet, N.; Aubert, J.P.; Swallow, D.; Gum, J.R.; Kim, Y.S.; Real, F.X.; Zweibaum, A. Differential expression of the human mucin genes muc1 to muc5 in relation to growth and differentiation of different mucus-secreting ht-29 cell subpopulations. J. Cell Sci. 1993, 106, 771. [Google Scholar] [PubMed]
- Wan, M.L.Y.; Turner, P.C.; Allen, K.J.; El-Nezami, H. Lactobacillus rhamnosus gg modulates intestinal mucosal barrier and inflammation in mice following combined dietary exposure to deoxynivalenol and zearalenone. J. Funct. Foods 2016, 22, 34–43. [Google Scholar] [CrossRef]
- Pinton, P.; Graziani, F.; Pujol, A.; Nicoletti, C.; Paris, O.; Ernouf, P.; Di Pasquale, E.; Perrier, J.; Oswald, I.P.; Maresca, M. Deoxynivalenol inhibits the expression by goblet cells of intestinal mucins through a pkr and map kinase dependent repression of the resistin-like molecule beta. Mol. Nutr. Food Res. 2015, 59, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, C.; Doohan, F.M. Trichothecene toxicity in eukaryotes: Cellular and molecular mechanisms in plants and animals. Toxicol. Lett. 2013, 217, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Yahi, N.; Younã¨S-Sakr, L.; Boyron, M.; Caporiccio, B.; Fantini, J. Both direct and indirect effects account for the pro-inflammatory activity of enteropathogenic mycotoxins on the human intestinal epithelium: Stimulation of interleukin-8 secretion, potentiation of interleukin-1beta effect and increase in the transepitheli. Toxicol. Appl. Pharmacol. 2008, 228, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Perrais, M.; Pigny, P.; Copin, M.C.; Aubert, J.P.; Van, S.I. Induction of muc2 and muc5ac mucins by factors of the epidermal growth factor (egf) family is mediated by egf receptor/ras/raf/extracellular signal-regulated kinase cascade and sp1. J. Biol. Chem. 2002, 277, 32258–32267. [Google Scholar] [CrossRef] [PubMed]
- Rodenburg, W.; Heidema, A.G.; Boer, J.M.A.; Boveeoudenhoven, I.M.J.; Feskens, E.J.M.; Mariman, E.C.M.; Keijer, J. A framework to identify physiological responses in microarray-based gene expression studies: Selection and interpretation of biologically relevant genes. Physiol. Genom. 2008, 33, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Candille, S.I.; Choi, Y.; Xie, D.; Jiang, L.; Lipookthan, J.; Tang, H.; Snyder, M. Variation and genetic control of protein abundance in humans. Nature 2013, 499, 79–82. [Google Scholar] [CrossRef] [PubMed]
- De Sousa Abreu, R.; Penalva, L.O.; Marcotte, E.M.; Vogel, C. Global signatures of protein and mrna expression levels. Mol. Biosyst. 2009, 5, 1512–1526. [Google Scholar] [CrossRef] [PubMed]
- Schwanhausser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Speijers, G.J.; Speijers, M.H. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, S.; Wang, J.; Luo, C.; Zhao, S.; Zheng, N. Modulation of intestinal epithelial permeability in differentiated caco-2 cells exposed to aflatoxin m1 and ochratoxin a individually or collectively. Toxins 2018, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Feudjio, F.T.; Dornetshuber, R.; Lemmens, M.; Hoffmann, O.; Lemmensgruber, R.; Berger, W.; Shephard, G.S. Beauvericin and enniatin: Emerging toxins and/or remedies? World Mycotoxin J. 2010, 3, 415–430. [Google Scholar] [CrossRef]
- Tozlovanu, M.; Canadas, D.; Pfohlleszkowicz, A.; Frenette, C.; Paugh, R.J.; Manderville, R.A. Glutationski konjugati okratoksina a kao biomarkeri izloženosti. Arhiv za Higijenu Rada i Toksikologiju 2012, 63, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Walter, E.; Janich, S.; Roessler, B.J.; Hilfinger, J.M.; Amidon, G.L. Ht29-mtx/caco-2 cocultures as an in vitro model for the intestinal epithelium: In vitro–in vivo correlation with permeability data from rats and humans. J. Pharm. Sci. 1996, 85, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Gill, N.; Wlodarska, M.; Finlay, B.B. Roadblocks in the gut: Barriers to enteric infection. Cell. Microbiol. 2011, 13, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.H.; Hasnain, S.Z.; Florin, T.H.J.; Mcguckin, M.A. Mucins in inflammatory bowel diseases and colorectal cancer. J. Gastroenterol. Hepatol. 2015, 27, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Grootjans, J.; Hundscheid, I.H.; Lenaerts, K.; Boonen, B.; Renes, I.B.; Verheyen, F.K.; Dejong, C.H.; von Meyenfeldt, M.F.; Beets, G.L.; Buurman, W.A. Ischaemia-induced mucus barrier loss and bacterial penetration are rapidly counteracted by increased goblet cell secretory activity in human and rat colon. Gut Microbes 2013, 62, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Connolly, L.; Frizzell, C.; Elliott, C.T. Cytotoxic assessment of the regulated, co-existing mycotoxins aflatoxin b1, fumonisin b1 and ochratoxin, in single, binary and tertiary mixtures. Toxicon 2014, 90, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; Dietrich, D.R.; O’Brien, E. In vitro investigation of individual and combined cytotoxic effects of ochratoxin a and other selected mycotoxins on renal cells. Toxicol. In Vitro 2006, 20, 332–341. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Set | Product Length (bp) | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|---|
| MUC2 | 238 | AAGACGGCACCTACCTCG | TTGGAGGAATAAACTGGAGAACC |
| MUC5AC | 278 | GTTTGACGGGAAGCAATACA | CGATGATGAAGAAGGTTGAGG |
| MUC5B | 171 | GTGACAACCGTGTCGTCCTG | TGCCGTCAAAGGTGGAATAG |
| GADPH | 235 | GGAGTCCACTGGCGTCTT | GAGTCCTTCCACGATACCAAA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Gao, Y.; Li, S.; Wu, C.; Wang, J.; Zheng, N. Modulation of Mucin (MUC2, MUC5AC and MUC5B) mRNA Expression and Protein Production and Secretion in Caco-2/HT29-MTX Co-Cultures Following Exposure to Individual and Combined Aflatoxin M1 and Ochratoxin A. Toxins 2019, 11, 132. https://doi.org/10.3390/toxins11020132
Huang X, Gao Y, Li S, Wu C, Wang J, Zheng N. Modulation of Mucin (MUC2, MUC5AC and MUC5B) mRNA Expression and Protein Production and Secretion in Caco-2/HT29-MTX Co-Cultures Following Exposure to Individual and Combined Aflatoxin M1 and Ochratoxin A. Toxins. 2019; 11(2):132. https://doi.org/10.3390/toxins11020132
Chicago/Turabian StyleHuang, Xin, Yanan Gao, Songli Li, Chenqing Wu, Jiaqi Wang, and Nan Zheng. 2019. "Modulation of Mucin (MUC2, MUC5AC and MUC5B) mRNA Expression and Protein Production and Secretion in Caco-2/HT29-MTX Co-Cultures Following Exposure to Individual and Combined Aflatoxin M1 and Ochratoxin A" Toxins 11, no. 2: 132. https://doi.org/10.3390/toxins11020132
APA StyleHuang, X., Gao, Y., Li, S., Wu, C., Wang, J., & Zheng, N. (2019). Modulation of Mucin (MUC2, MUC5AC and MUC5B) mRNA Expression and Protein Production and Secretion in Caco-2/HT29-MTX Co-Cultures Following Exposure to Individual and Combined Aflatoxin M1 and Ochratoxin A. Toxins, 11(2), 132. https://doi.org/10.3390/toxins11020132