Pharmacokinetic Properties of the Nephrotoxin Orellanine in Rats

 , ,
, ,
Abstract
1. Introduction
2. Results
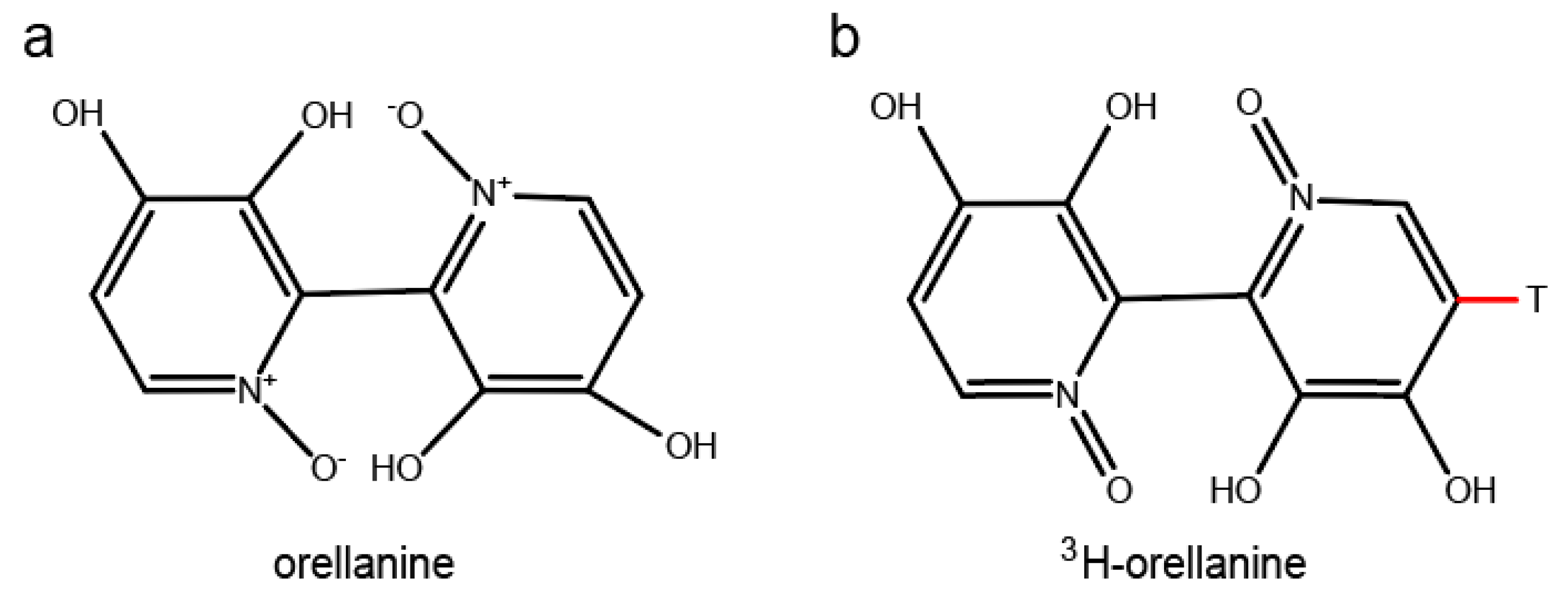
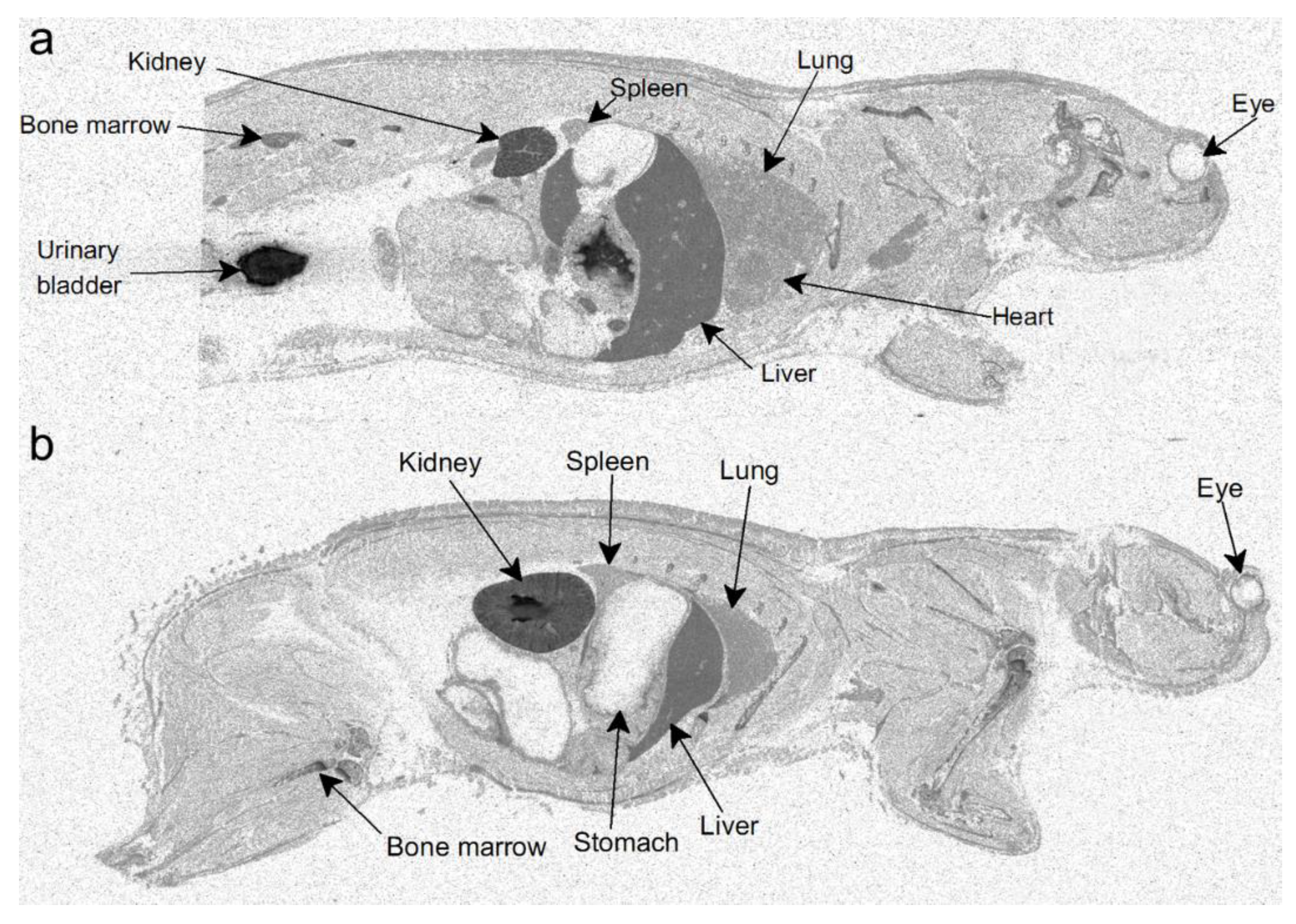
2.1. Radioluminography
2.2. Pharmacokinetics Study: General Condition of Animals
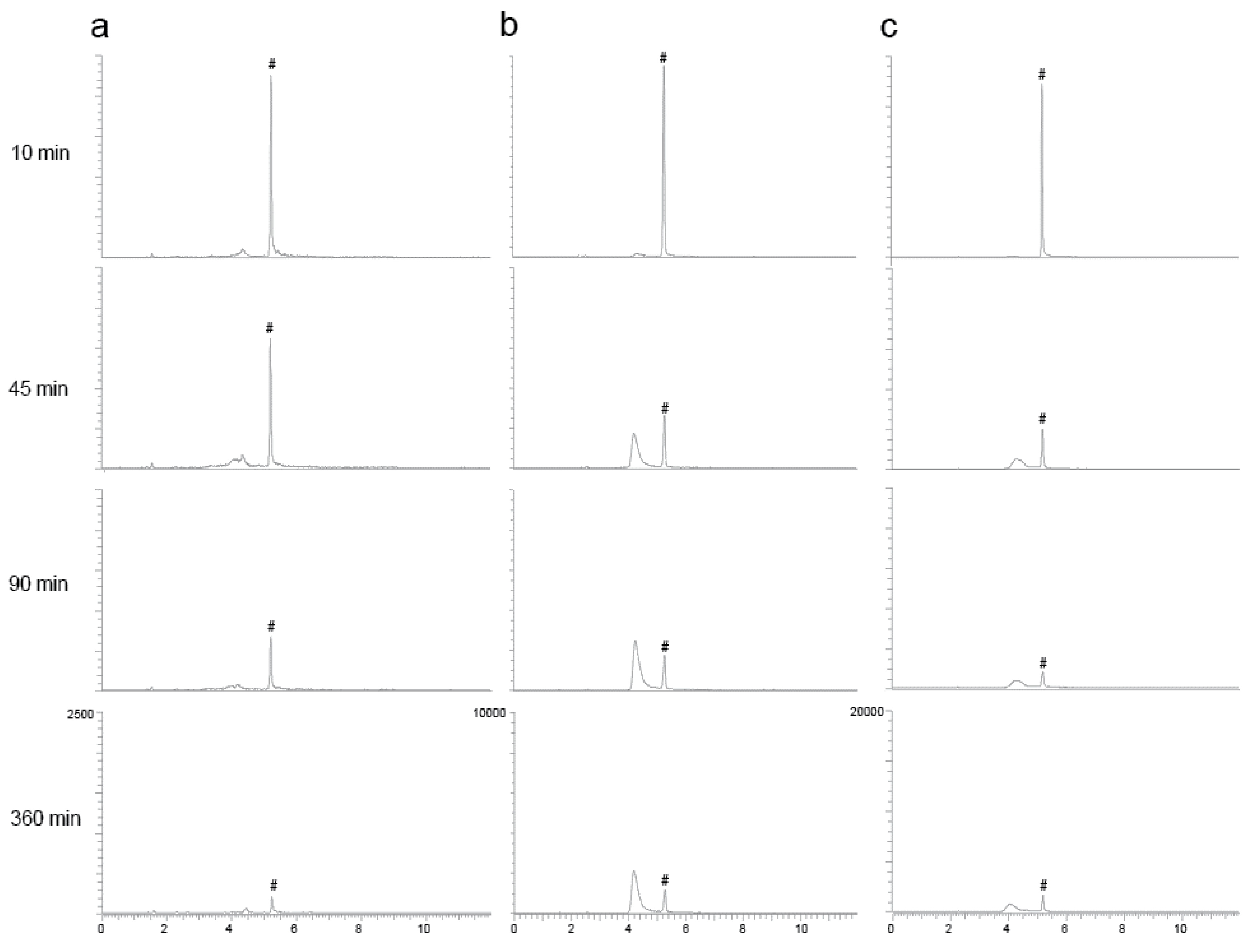
2.3. Parallel Reaction Monitoring (PRM) of Orellanine
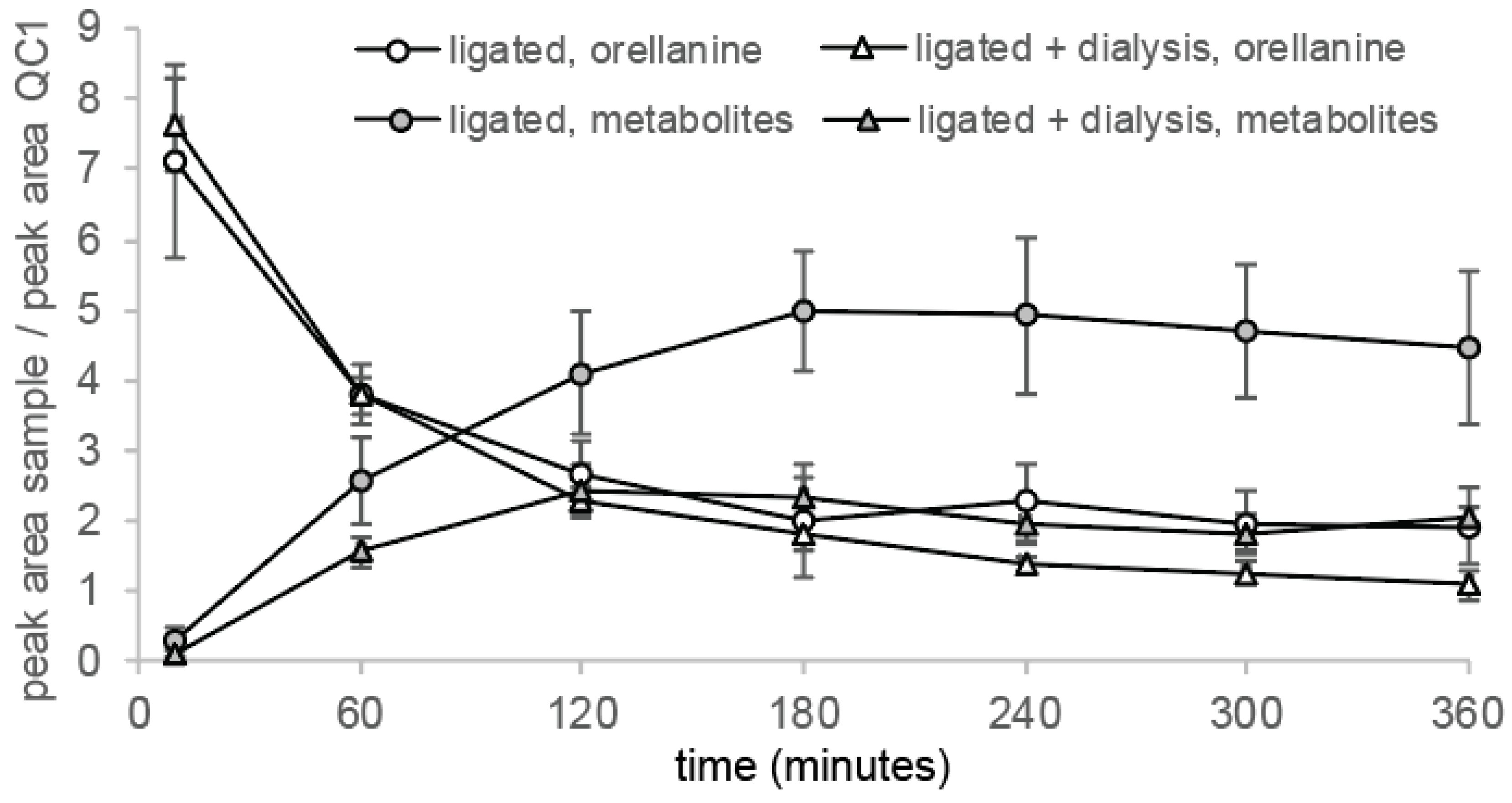
2.4. Plasma Concentration of Orellanine Versus Time and Half-Life
2.5. Clearance of Orellanine
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Test Solution of Orellanine
5.2. Rat Experiments
5.3. Radioluminography
5.4. Pharmacokinetic Studies
5.5. Beta Scintillation of 3H
5.6. Bioanalysis of Orellanine
5.7. Pharmacokinetic Analysis
5.8. Statistics
5.9. Ethical Approval
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grzymala, S. Massenvergiftungen durch den orangefuchsigen hautkopf. Z. Pilzkd 1957, 23, 139–142. [Google Scholar]
- Mottonen, M.; Nieminen, L.; Heikkila, H. Damage caused by two finnish mushrooms, cortinarius speciosissimus and cortinarius gentilis on the rat kidney. Z. Naturforsch C 1975, 30, 668–671. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, U.A.; Nystrom, J.; Buvall, L.; Ebefors, K.; Bjornson-Granqvist, A.; Holmdahl, J.; Haraldsson, B. The fungal nephrotoxin orellanine simultaneously increases oxidative stress and down-regulates cellular defenses. Free Radic. Biol. Med. 2008, 44, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, T.; Hoiland, K. Mushroom poisoning caused by species of the genus cortinarius fries. Arch. Toxicol. 1983, 53, 87–106. [Google Scholar] [PubMed]
- Cantin-Esnault, D.; Richard, J.M.; Jeunet, A. Generation of oxygen radicals from iron complex of orellanine, a mushroom nephrotoxin; preliminary esr and spin-trapping studies. Free Radic. Res. 1998, 28, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.M.; Cantin-Esnault, D.; Jeunet, A. First electron spin resonance evidence for the production of semiquinone and oxygen free radicals from orellanine, a mushroom nephrotoxin. Free Radic. Biol. Med. 1995, 19, 417–429. [Google Scholar] [CrossRef]
- Herrmann, A.; Hedman, H.; Rosen, J.; Jansson, D.; Haraldsson, B.; Hellenas, K.E. Analysis of the mushroom nephrotoxin orellanine and its glucosides. J. Nat. Prod. 2012, 75, 1690–1696. [Google Scholar] [CrossRef] [PubMed]
- Buvall, L.; Hedman, H.; Khramova, A.; Najar, D.; Bergwall, L.; Ebefors, K.; Sihlbom, C.; Lundstam, S.; Herrmann, A.; Wallentin, H.; et al. Orellanine specifically targets renal clear cell carcinoma. Oncotarget 2017, 8, 91085–91098. [Google Scholar] [CrossRef] [PubMed]
- Antkowiak, W.Z.; Gessner, W.P. The structures of orellanine and orelline. Tetrahedron Lett. 1979, 20, 1931–1934. [Google Scholar] [CrossRef]
- Antkowiak, W.Z.; Gessner, W.P. Photodecomposition of orellanine and orellinine, the fungal toxins ofcortinarius orellanus fries andcortinarius speciossimus. Cell Mol. Life Sci. 1985, 41, 769–771. [Google Scholar] [CrossRef]
- Dehmlow, E.V.; Schulz, H.-J. Synthesis of orellanine the lethal poison of a toadstool. Tetrahedron Lett. 1985, 26, 4903–4906. [Google Scholar] [CrossRef]
- Spiteller, P.; Spiteller, M.; Steglich, W. Occurrence of the fungal toxin orellanine as a diglucoside and investigation of its biosynthesis. Angew. Chem. Int. Ed. Engl. 2003, 42, 2864–2867. [Google Scholar] [CrossRef] [PubMed]
- Dinis-Oliveira, R.J.; Soares, M.; Rocha-Pereira, C.; Carvalho, F. Human and experimental toxicology of orellanine. Hum. Exp. Toxicol. 2016, 35, 1016–1029. [Google Scholar] [CrossRef] [PubMed]
- Oubrahim, H.; Richard, J.M.; Cantin-Esnault, D.; Seigle-Murandi, F.; Trecourt, F. Novel methods for identification and quantification of the mushroom nephrotoxin orellanine. Thin-layer chromatography and electrophoresis screening of mushrooms with electron spin resonance determination of the toxin. J. Chromatogr. A 1997, 758, 145–157. [Google Scholar] [CrossRef]
- Calvino, J.; Romero, R.; Pintos, E.; Novoa, D.; Guimil, D.; Cordal, T.; Mardaras, J.; Arcocha, V.; Lens, X.M.; Sanchez-Guisande, D. Voluntary ingestion of cortinarius mushrooms leading to chronic interstitial nephritis. Am. J. Nephrol. 1998, 18, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Delpech, N.; Rapior, S.; Cozette, A.P.; Ortiz, J.P.; Donnadieu, P.; Andary, C.; Huchard, G. Outcome of acute renal failure caused by voluntary ingestion of cortinarius orellanus. Presse Med. 1990, 19, 122–124. [Google Scholar] [PubMed]
- Prast, H.; Pfaller, W. Toxic properties of the mushroom cortinarius orellanus (fries). II. Impairment of renal function in rats. Arch. Toxicol. 1988, 62, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Hedman, H.; Holmdahl, J.; Mölne, J.; Ebefors, K.; Haraldsson, B.; Nyström, J. Long-term clinical outcome for patients poisoned by the fungal nephrotoxin orellanine. BMC Nephrol. 2017, 18, 121. [Google Scholar] [CrossRef] [PubMed]
- Rohrmoser, M.; Kirchmair, M.; Feifel, E.; Valli, A.; Corradini, R.; Pohanka, E.; Rosenkranz, A.; Poder, R. Orellanine poisoning: Rapid detection of the fungal toxin in renal biopsy material. J. Toxicol. Clin. Toxicol. 1997, 35, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Rapior, S.; Delpech, N.; Andary, C.; Huchard, G. Intoxication by cortinarius orellanus: Detection and assay of orellanine in biological fluids and renal biopsies. Mycopathologia 1989, 108, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Danel, V.C.; Saviuc, P.F.; Garon, D. Main features of cortinarius spp. poisoning: A literature review. Toxicon 2001, 39, 1053–1060. [Google Scholar] [CrossRef]
- Capitanio, U.; Montorsi, F. Renal cancer. Lancet 2016, 387, 894–906. [Google Scholar] [CrossRef]
- Thorstenson, A.; Bergman, M.; Scherman-Plogell, A.H.; Hosseinnia, S.; Ljungberg, B.; Adolfsson, J.; Lundstam, S. Tumour characteristics and surgical treatment of renal cell carcinoma in sweden 2005–2010: A population-based study from the national swedish kidney cancer register. Scand. J. Urol. 2014, 48, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.L.; Chang, A.; Perazella, M.A.; Okusa, M.D.; Jaimes, E.A.; Weiss, R.H.; American Society of Nephrology Onco-Nephrology Forum. The nephrologist's tumor: Basic biology and management of renal cell carcinoma. J. Am. Soc. Nephrol. 2016, 27, 2227–2237. [Google Scholar] [CrossRef] [PubMed]
- Albiges, L.; Choueiri, T.; Escudier, B.; Galsky, M.; George, D.; Hofmann, F.; Lam, T.; Motzer, R.; Mulders, P.; Porta, C.; et al. A systematic review of sequencing and combinations of systemic therapy in metastatic renal cancer. Eur. Urol. 2015, 67, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.; Morris, T. Physiological parameters in humans and laboratory animals. Pharm. Res. 1993, 10, 1093–1095. [Google Scholar] [CrossRef] [PubMed]
- Mercatello, A. Changes in renal function induced by anesthesia. Ann. Fr. Anesth. Reanim. 1990, 9, 507–524. [Google Scholar] [CrossRef]
- Burchardi, H.; Kaczmarczyk, G. The effect of anaesthesia on renal function. Eur. J. Anaesthesiol. 1994, 11, 163–168. [Google Scholar] [PubMed]
- Pfaller, W.; Gstraunthaler, G.; Prast, H.; Rupp, L.; Ruedl, C.; Michelitsch, S.; Moser, M. Effects of the Fungal Toxin Orellanine on Renal Epithelium; Marcel Dekker: New York, NY, USA, 1991; pp. 63–69. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Point (h) | 0.5 | 1 | 6 | 12 | 24 |
|---|---|---|---|---|---|
| Adrenal gland | 5.2 | 2.6 | 1.1 | 1.6 | <LOQ |
| Bone marrow | 15.0 | 4.7 | 3.9 | 4.0 | 3.0 |
| Brain | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Brown fat | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Connective Tissue | 9.7 | 3.2 | 1.6 | 2.0 | <LOQ |
| Dermis | 3.2 | 1.4 | <LOQ | <LOQ | <LOQ |
| Epidermis | 4.0 | 1.9 | <LOQ | <LOQ | <LOQ |
| Gastric mucosa | 3.2 | 1.8 | 1.1 | <LOQ | <LOQ |
| Heart blood | 8.6 | 4.9 | 2.0 | 1.7 | 1.3 |
| Intestinal mucosa | 5.3 | 4.2 | 0.8 | 0.8 | 1.0 |
| Kidney cortex | 76.0 | 73.0 | 19.0 | 18.0 | 15.0 |
| Lens (eye) | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Liver | 35.0 | 25.0 | 7.8 | 6.1 | 4.0 |
| Lung | 8.0 | 4.0 | 1.6 | 1.4 | 0.9 |
| Lymph node | 3.9 | 2.1 | 0.8 | 14.0 | <LOQ |
| Myocardium | 6.0 | 2.6 | 1.1 | 0.8 | 0.6 |
| Pancreas | 3.9 | 1.9 | 1.6 | 1.0 | <LOQ |
| Salivary gland | 4.5 | 2.6 | 1.2 | 1.0 | 0.7 |
| Skeletal muscle | 1.8 | 1.6 | <LOQ | <LOQ | <LOQ |
| Spleen | 6.5 | 5.6 | 4.3 | 3.8 | 3.3 |
| Testicle | 1.9 | 1.3 | <LOQ | <LOQ | <LOQ |
| Thymus | 4.9 | 2.5 | 1.2 | 1.1 | 1.1 |
| Thyroid gland | 5.2 | 2.5 | 1.7 | 0.9 | 0.6 |
| Urinary bladder | 560.0 | 2600.0 | 200.0 | 37.0 | 18.0 |
| Limit of Quantification (LOQ) | 0.6 | 0.6 | 0.7 | 0.52–0.67 | 0.6 |
| Time Point (h) | 0.5 | 1 | 6 | 12 |
|---|---|---|---|---|
| Adrenal gland | 5.1 | 8.0 | 2.1 | 0.7 |
| Bone marrow | 10.0 | 5.8 | 6.1 | 4.4 |
| Brain | <LOQ | <LOQ | <LOQ | <LOQ |
| Brown fat | <LOQ | 1.3 | <LOQ | <LOQ |
| Connective tissue (skin) | 10.0 | 12.0 | 1.4 | 0.8 |
| Dermis | 2.0 | 2.0 | <LOQ | <LOQ |
| Epidermis | 4.5 | 6.9 | 1.9 | <LOQ |
| Gastric mucosa | 3.8 | 4.8 | 1.9 | 1.0 |
| Heart blood | 9.0 | 12.0 | 3.5 | 1.8 |
| Intestinal mucosa | 3.3 | 9.2 | 3.2 | 2.1 |
| Kidney cortex | 63.0 | 90.0 | 23.0 | 22.0 |
| Lens (eye) | <LOQ | <LOQ | <LOQ | <LOQ |
| Liver | 29.0 | 37.0 | 18.0 | 7.2 |
| Lung | 8.8 | 10.5 | 3.0 | 1.3 |
| Lymph node | 2.7 | 7.0 | 3.7 | 2.6 |
| Myocardium | 4.7 | 6.3 | 2.6 | 1.8 |
| Pancreas | 2.9 | 5.1 | 2.5 | 1.7 |
| Pituitary | 3.1 | 5.6 | 2.2 | 2.1 |
| Retina | 8.3 | 11.0 | 2.4 | 1.0 |
| Salivary gland | 3.8 | 6.2 | 3.3 | 2.3 |
| Skeletal muscle | 1.9 | 2.1 | 2.0 | 0.9 |
| Spleen | 4.6 | 8.8 | 6.8 | 3.5 |
| Testicle | 2.4 | 2.7 | 1.0 | <LOQ |
| Thymus | 4.0 | 6.2 | 4.3 | 2.2 |
| Thyroid gland | 4.5 | 6.6 | 2.4 | 1.2 |
| Urinary bladder | 590.0 | 600.0 | 93.0 | 15.0 |
| LOQ | 0.6 | 0.6 | 0.43–0.65 | 0.54–0.70 |
| Parameter | LC-MS/MS, Orellanine | Beta Scintillation, 3H-Orellanine | ||||
|---|---|---|---|---|---|---|
| Intact Renal Function | Ligated Kidneys | Ligated Kidney Dialysis | Intact Renal Function | Ligated Kidneys | Ligated Kidney Dialysis | |
| Intercept (nmol/L) | 2115 ± 342 | 3028 ± 610 | 2521 ± 149 | 1471 ± 71 | 2538 ± 115 | 2773 ± 135 |
| Rate of elimination (nmol/mL/min) | 0.00651 ± 0.00032 | 0.000101 ± 0.00013 | 0.00338 ± 0.00063 | 0.0031 ± 0.00012 | 0.00085 ± 0.00019 | 0.00122 ± 0.00006 |
| Half-life (t½, min) | 109 ± 60 | 756 ± 98 | 238 ± 28 | 225 ± 10 | 1033 ± 183 | 583 ± 30 |
| Dose injected (nmol) | 87.3 ± 1.5 | 85.4 ± 1.3 | 86.9 ± 1.2 | 87.6 ± 1.2 | 84.9 ± 1.3 | 87.2 ± 1.1 |
| Area under the curve (AUC, nmol × min/L) | 503,870 ± 70,670 | 3,494,674 ± 783,934 | 645,606 ± 99,420 | 821,601 ± 52,808 | 3,223,824 ± 230,783 | 2,823,938 ± 123,739 |
| Volume of Distribution (VD, mL) | 31.7 ± 4.8 | 35.1 ± 8.6 | 49.1 ± 4.7 | 35.3 ± 1.7 | 37.7 ± 4.9 | 26.3 ± 1.7 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Najar, D.; Haraldsson, B.; Thorsell, A.; Sihlbom, C.; Nyström, J.; Ebefors, K. Pharmacokinetic Properties of the Nephrotoxin Orellanine in Rats. Toxins 2018, 10, 333. https://doi.org/10.3390/toxins10080333
Najar D, Haraldsson B, Thorsell A, Sihlbom C, Nyström J, Ebefors K. Pharmacokinetic Properties of the Nephrotoxin Orellanine in Rats. Toxins. 2018; 10(8):333. https://doi.org/10.3390/toxins10080333
Chicago/Turabian StyleNajar, Deman, Börje Haraldsson, Annika Thorsell, Carina Sihlbom, Jenny Nyström, and Kerstin Ebefors. 2018. "Pharmacokinetic Properties of the Nephrotoxin Orellanine in Rats" Toxins 10, no. 8: 333. https://doi.org/10.3390/toxins10080333
APA StyleNajar, D., Haraldsson, B., Thorsell, A., Sihlbom, C., Nyström, J., & Ebefors, K. (2018). Pharmacokinetic Properties of the Nephrotoxin Orellanine in Rats. Toxins, 10(8), 333. https://doi.org/10.3390/toxins10080333