Human Poisoning from Marine Toxins: Unknowns for Optimal Consumer Protection

 , , ,
, , , 
Abstract
1. Introduction
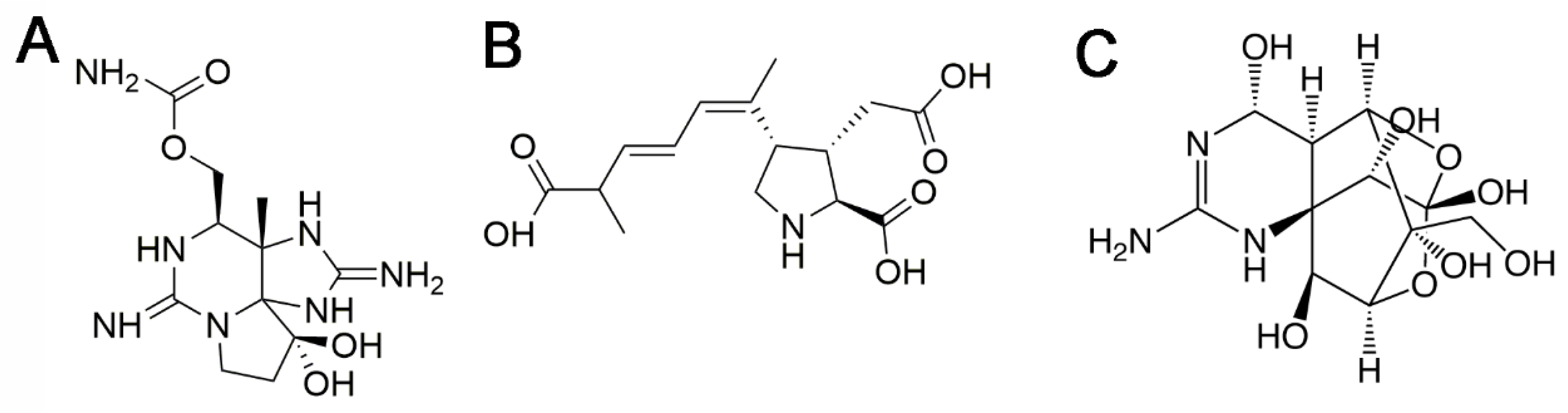
2. Saxitoxins
2.1. Acute Human Intoxication: Paralytic Shellfish Poisoning
2.2. Chronic Toxicity of Saxitoxins
2.3. Relative Toxic Potency of Saxitoxin Analogs
2.4. Limitations and Implications of Current Toxicological Information on Saxitoxins
3. Domoic Acid
3.1. Acute Human Intoxication: Amnesic Shellfish Poisoning
3.2. Chronic Toxicity of Domoic Acid
3.3. Relative Toxic Potency of Domoic Acid Analogs
3.4. Limitations and Implications of Current Toxicological Information on Domoic Acid
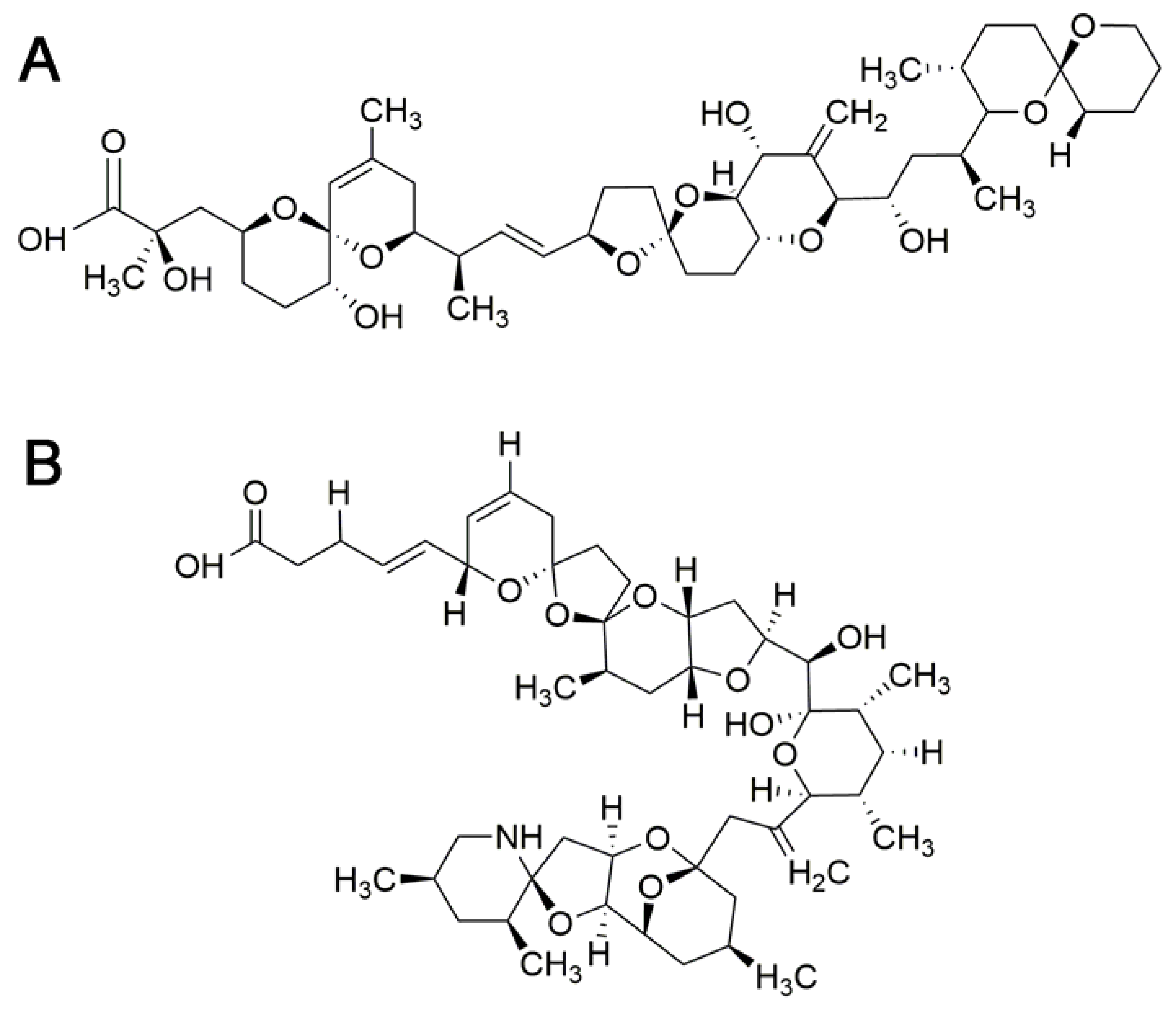
4. Ciguatoxins
4.1. Acute Human Intoxication: Ciguatera
4.2. Ciguatoxin Chronic Toxicity
4.3. Relative Toxic Potency of Ciguatoxin Analogs
4.4. Limitations and Implications of Current Ciguatoxin Toxicity Data
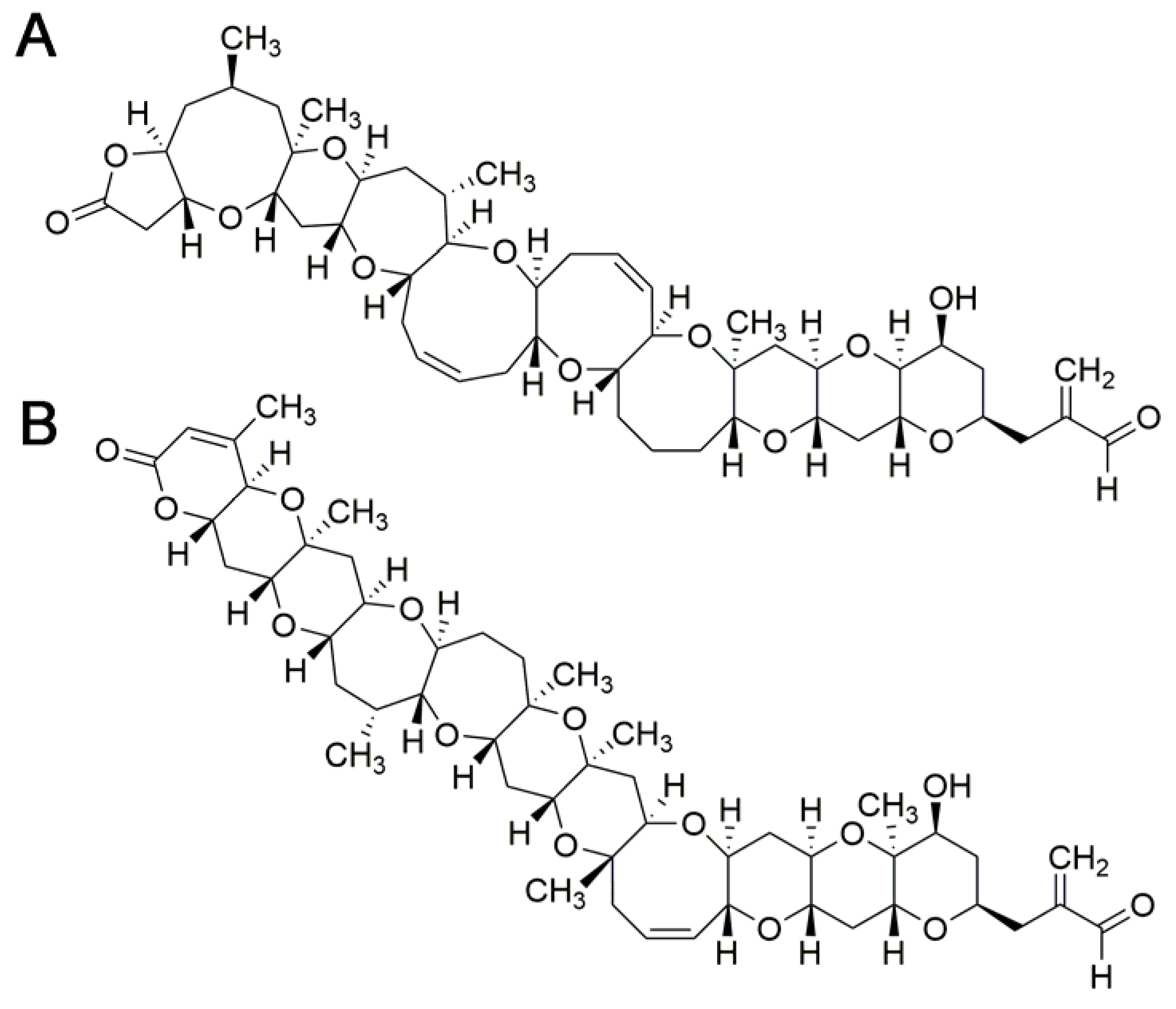
5. Brevetoxins
5.1. Acute Human Intoxication: Neurotoxin Shellfish Poisoning
5.2. Chronic Toxicity of Brevetoxins
5.3. Relative Toxic Potency of Brevetoxin Analogs
5.4. Limitations and Implications of Current Toxicological Information on Brevetoxins
6. Tetrodotoxins
6.1. Acute Human Intoxication
6.2. Chronic Toxicity of Tetrodotoxins
6.3. Relative Toxic Potency of Tetrodotoxin Analogs
6.4. Limitations and Implications of Current Toxicological Information on Tetrodotoxin
7. Okadaic Acid Group
7.1. Acute Human Intoxication: Diarrheic Shellfish Poisoning
7.2. Chronic Toxicity of Okadaic Acid
7.3. Relative Toxic Potency of Okadaic Acid Analogs
7.4. Limitations and Implications of Current Toxicological Data
8. Azaspiracids
8.1. Acute Human Intoxication: Azaspiracid Poisoning
8.2. Azaspiracid Chronic Toxicity
8.3. Relative Toxic Potency of Azaspiracids
8.4. Limitations and Implications of Current Azaspiracid Toxicity Data
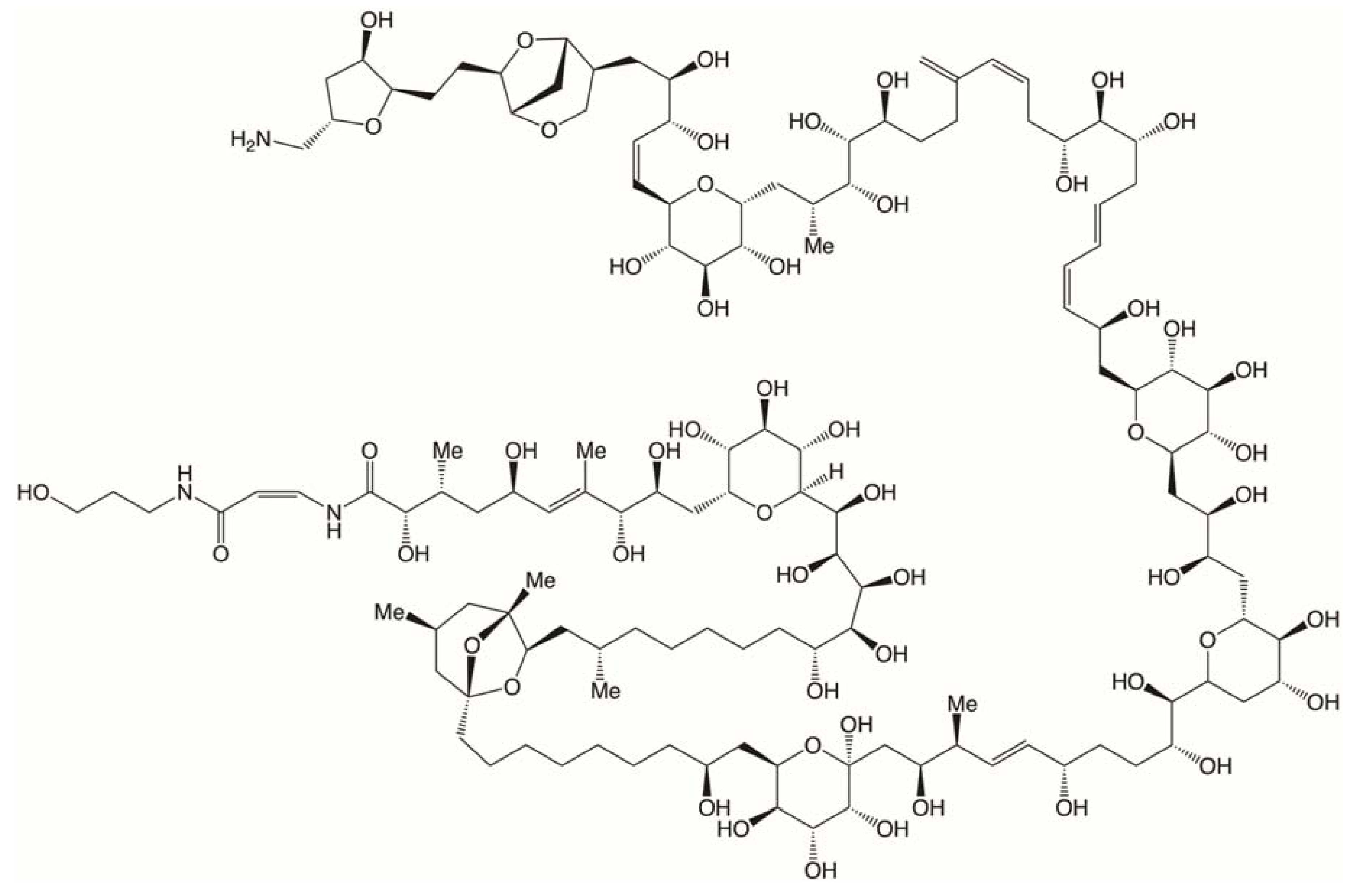
9. Palytoxin
9.1. Human Acute Toxicity of Palytoxins
9.2. Palytoxin Chronic Toxicity
9.3. Relative Toxic Potency of Palytoxins
9.4. Limitations and Implications of Current Palytoxin Toxicity Data
10. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Deeds, J.R.; Landsberg, J.H.; Etheridge, S.M.; Pitcher, G.C.; Longan, S.W. Non-traditional vectors for paralytic shellfish poisoning. Mar. Drugs 2008, 6, 308–348. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Scientific Opinion of the Panel on Contaminants in the Food Chain on a request from the European Commission on Marine biotoxins in shellfish–saxitoxin group. EFSA J. 2009, 1019, 1–76. [Google Scholar]
- Etheridge, S.M. Paralytic shellfish poisoning: Seafood safety and human health perspectives. Toxicon 2010, 56, 108–122. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, K.; Musgrave, I.F.; Humpage, A. Low dose extended exposure to saxitoxin and its potential neurodevelopmental effects: A review. Environ. Toxicol. Pharmacol. 2016, 48, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Negri, A.P.; Jones, G.J.; Hindmarsh, M. Sheep mortality associated with paralytic shellfish poisons from the cyanobacterium Anabaena circinalis. Toxicon 1995, 33, 1321–1329. [Google Scholar] [CrossRef]
- Testai, E.; Scardala, S.; Vichi, S.; Buratti, F.M.; Funari, E. Risk to human health associated with the environmental occurrence of cyanobacterial neurotoxic alkaloids anatoxins and saxitoxins. Crit. Rev. Toxicol. 2016, 46, 385–419. [Google Scholar] [CrossRef] [PubMed]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed]
- Hallegraeff, G.M. Harmful algal blooms: A global overview. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; Imprimerie Landais: Paris, France, 2005; pp. 25–49. [Google Scholar]
- Prakash, A.; Medcof, J.C.; Tennant, A.D. Paralytic shellfish poisoning in eastern Canada. Fish. Board Res. Can. Ott. Bull. 1971, 177, 1–87. [Google Scholar]
- Duran-Riveroll, L.M.; Cembella, A.D. Guanidinium Toxins and Their Interactions with Voltage-Gated Sodium Ion Channels. Mar. Drugs 2017, 15, 303. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.; del Carmen Bravo, M.; Lagos, M.; Lagos, N. Paralytic shellfish poisoning: Post-mortem analysis of tissue and body fluid samples from human victims in the Patagonia fjords. Toxicon 2004, 43, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, L.E.; Dodd, M.J.; Robertson, A.; Ericson, G.; de Koning, C.; Negri, A.P. Post-mortem analysis of samples from a human victim of a fatal poisoning caused by the xanthid crab, Zosimus aeneus. Toxicon 2002, 40, 1463–1469. [Google Scholar] [CrossRef]
- Garcia, C.; Rodriguez-Navarro, A.; Diaz, J.C.; Torres, R.; Lagos, N. Evidence of in vitro glucuronidation and enzymatic transformation of paralytic shellfish toxins by healthy human liver microsomes fraction. Toxicon 2009, 53, 206–213. [Google Scholar] [CrossRef] [PubMed]
- FAO/IOC/WHO. Report of the Joint FAO/IOC/WHO ad Hoc Expert Consultation on Biotoxins in Bivalve Molluscs; IOC: Oslo, Norway, 2004. [Google Scholar]
- Arnich, N.; Thebault, A. Dose-Response Modelling of Paralytic Shellfish Poisoning (PSP) in Humans. Toxins 2018, 10, 141. [Google Scholar] [CrossRef] [PubMed]
- Zepeda, R.J.; Candiracci, M.; Lobos, N.; Lux, S.; Miranda, H.F. Chronic toxicity study of neosaxitoxin in rats. Mar. Drugs 2014, 12, 5055–5071. [Google Scholar] [CrossRef] [PubMed]
- Ramos, P.B.; Diehl, F.; dos Santos, J.M.; Monserrat, J.M.; Yunes, J.S. Oxidative stress in rats induced by consumption of saxitoxin contaminated drink water. Harmful Algae 2014, 37, 68–74. [Google Scholar] [CrossRef]
- Diehl, F.; Ramos, P.B.; Dos Santos, J.M.; Barros, D.M.; Yunes, J.S. Behavioral alterations induced by repeated saxitoxin exposure in drinking water. J. Venom. Anim. Toxins Incl. Trop. Dis. 2016, 22, 18. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.Z.; Lam, P.K.; Hsieh, D.P. Interactions of paralytic shellfish toxins with xenobiotic-metabolizing and antioxidant enzymes in rodents. Toxicon 2003, 42, 425–431. [Google Scholar] [CrossRef]
- Botana, L.; Hess, P.; Munday, R.; Nathalie, A.; DeGrasse, S.; Feeley, M.; Suzuki, T.; Van den Berg, M.; Fattori, V.; Gamarro, E.; et al. Derivation of toxicity equivalency factors for marine biotoxins associated with Bivalve Molluscs. Trends Food Sci. Technol. 2017, 59, 15–24. [Google Scholar] [CrossRef]
- Botana, L.; Vilariño, N.; Alfonso, A.; Vale, C.; Louzao, M.; Elliott, C.; Campbell, K.; Botana, A. The problem of toxicity equivalent factors in developing alternative methods to animal bioassays for marine-toxin detection. Trends Anal. Chem. 2010, 29, 1316–1325. [Google Scholar] [CrossRef]
- EC. Comission Regulation (EC) No. 2074/2005 of 5 December 2005 laying down implementing measures for certain products under Regulation (EC) No. 853/2004 of the European Parliament and of the Council and for the organisation of official controls under Regulation (EC) No. 854/2204 of the European Parliament and of the Council and Regulation (EC) No. 882/2204 of the European Parliament and of the Council, derogating from Regulation (EC) No. 852/2004 of the European Parliament and of the Council and amending Regulations (EC) No. 853/2004 and (EC) No. 854/2004. Off. J. Eur. Comm. 2005, L338, 27. [Google Scholar]
- EU. Commission Regulation 2017/1980 of 31 October 2017 amending Annex III to Regulation (EC) No. 2074/2005 as regards paralytic shellfish poison (PSP) detection method. Off. J. Eur. Union 2017, L285, 8. [Google Scholar]
- AOAC. Official method 2005.06. Quantitative determination of Paralytic Shellfish Poisoning Toxins in shellfish using pre-chromatographic oxidation and liquid chromatography with fluorescence detection. In Official Methods of Analysis of AOAC International, 18th ed.; AOAC: Gaithersburg, MD, USA, 2006. [Google Scholar]
- AOAC. Official Method 2011.02. Paralytic Shellfish Toxins in Mussels, Clams, Oysters, and Scallops. Post-Column Oxidation (PCOX) Method. In Official Methods of Analysis of AOAC International, 19th ed.; AOAC: Gaithersburg, MD, USA, 2012. [Google Scholar]
- Vale, C.; Alfonso, A.; Vieytes, M.R.; Romaris, X.M.; Arevalo, F.; Botana, A.M.; Botana, L.M. In vitro and in vivo evaluation of paralytic shellfish poisoning toxin potency and the influence of the pH of extraction. Anal. Chem. 2008, 80, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO. Technical Paper on Toxicity Equivalency Factors for Marine Biotoxins Associated with Bivalve Molluscs; FAO: Rome, Italy, 2016. [Google Scholar]
- Munday, R.; Thomas, K.; Gibbs, R.; Murphy, C.; Quilliam, M.A. Acute toxicities of saxitoxin, neosaxitoxin, decarbamoyl saxitoxin and gonyautoxins 1&4 and 2&3 to mice by various routes of administration. Toxicon 2013, 76, 77–83. [Google Scholar] [PubMed]
- Selwood, A.I.; Waugh, C.; Harwood, D.T.; Rhodes, L.L.; Reeve, J.; Sim, J.; Munday, R. Acute Toxicities of the Saxitoxin Congeners Gonyautoxin 5, Gonyautoxin 6, Decarbamoyl Gonyautoxin 2&3, Decarbamoyl Neosaxitoxin, C-1&2 and C-3&4 to Mice by Various Routes of Administration. Toxins 2017, 9, 73. [Google Scholar]
- Vilariño, N.; Louzao, M.C.; Fraga, M.; Botana, L.M. From science to policy: Dynamic adaptation of legal regulations on aquatic biotoxins. In Climate Change and Marine and Freshwater Toxins; Botana, L.M., Louzao, M.C., Vilariño, N., Eds.; De Gruyter: Berlin, Germany, 2015; pp. 441–482. [Google Scholar]
- EC. Commission regulation (EC) No. 853/2004 of the European parliament and of the council of 29 April 2004 laying down specific hygiene rules for on the hygiene of foodstuffs. Off. J. Eur. Union 2004, 139, 55–205. [Google Scholar]
- Alimentarius, C. Standard for Live and Raw Bivalve Molluscs. In Codex Alimentarius; Food and Agriculture Organization of the United Nations: Rome, Italy, 2008. [Google Scholar]
- Dhar, B.C.; Cimarelli, L.; Singh, K.S.; Brandi, L.; Brandi, A.; Puccinelli, C.; Marcheggiani, S.; Spurio, R. Molecular detection of a potentially toxic diatom species. Int. J. Environ. Res. Public Health 2015, 12, 4921–4941. [Google Scholar] [CrossRef] [PubMed]
- Maeno, Y.; Kotaki, Y.; Terada, R.; Cho, Y.; Konoki, K.; Yotsu-Yamashita, M. Six domoic acid related compounds from the red alga, Chondria armata, and domoic acid biosynthesis by the diatom, Pseudo-nitzschia multiseries. Sci. Rep. 2018, 8, 356. [Google Scholar] [CrossRef] [PubMed]
- Mos, L. Domoic acid: A fascinating marine toxin. Environ. Toxicol. Pharmacol. 2001, 9, 79–85. [Google Scholar] [CrossRef]
- Quilliam, M.A.; Wright, J.L. The amnesic shellfish poisoning mystery. Anal. Chem. 1989, 61, 1053A–1106A. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rivera, A.; Pinto, M.; Insinilla, A.; Suarez Isla, B.; Uribe, E.; Alvarez, G.; Lehane, M.; Furey, A.; James, K.J. The occurrence of domoic acid linked to a toxic diatom bloom in a new potential vector: The tunicate Pyura chilensis (piure). Toxicon 2009, 54, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Ferriss, B.E.; Marcinek, D.J.; Ayres, D.; Borchert, J.; Lefebvre, K.A. Acute and chronic dietary exposure to domoic acid in recreational harvesters: A survey of shellfish consumption behavior. Environ. Int. 2017, 101, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Maucher Fuquay, J.; Muha, N.; Wang, Z.; Ramsdell, J.S. Elimination kinetics of domoic acid from the brain and cerebrospinal fluid of the pregnant rat. Chem. Res. Toxicol. 2012, 25, 2805–2809. [Google Scholar] [CrossRef] [PubMed]
- Pulido, O.M. Domoic acid toxicologic pathology: A review. Mar. Drugs 2008, 6, 180–219. [Google Scholar] [CrossRef] [PubMed]
- Vale, C. Domoic acid: Chemistry and Pharmacology. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection, 3rd ed.; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 875–894. [Google Scholar]
- Hogberg, H.T.; Bal-Price, A.K. Domoic Acid-Induced Neurotoxicity Is Mainly Mediated by the AMPA/KA Receptor: Comparison between Immature and Mature Primary Cultures of Neurons and Glial Cells from Rat Cerebellum. J. Toxicol. 2011, 2011, 543512. [Google Scholar] [CrossRef] [PubMed]
- Andjelkovic, M.; Vandevijvere, S.; Van Klaveren, J.; Van Oyen, H.; Van Loco, J. Exposure to domoic acid through shellfish consumption in Belgium. Environ. Int. 2012, 49, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, J.S.; Zatorre, R.J.; Carpenter, S.; Gendron, D.; Evans, A.C.; Gjedde, A.; Cashman, N.R. Neurologic sequelae of domoic acid intoxication due to the ingestion of contaminated mussels. N. Engl. J. Med. 1990, 322, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- Doucette, T.A.; Tasker, R.A. Domoic acid: Detection methods, pharmacology and toxicology. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection, 2nd ed.; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 397–429. [Google Scholar]
- Grattan, L.M.; Boushey, C.J.; Liang, Y.; Lefebvre, K.A.; Castellon, L.J.; Roberts, K.A.; Toben, A.C.; Morris, J.G. Repeated Dietary Exposure to Low Levels of Domoic Acid and Problems with Everyday Memory: Research to Public Health Outreach. Toxins 2018, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Ramsdell, J.S.; Gulland, F.M. Domoic acid epileptic disease. Mar. Drugs 2014, 12, 1185–1207. [Google Scholar] [CrossRef] [PubMed]
- Perl, T.M.; Bedard, L.; Kosatsky, T.; Hockin, J.C.; Todd, E.C.; McNutt, L.A.; Remis, R.S. Amnesic shellfish poisoning: A new clinical syndrome due to domoic acid. Can. Dis. Wkly. Rep. 1990, 16, 7–8. [Google Scholar] [PubMed]
- Suzuki, C.A.; Hierlihy, S.L. Renal clearance of domoic acid in the rat. Food Chem. Toxicol. 1993, 31, 701–706. [Google Scholar] [CrossRef]
- Truelove, J.; Mueller, R.; Pulido, O.; Iverson, F. Subchronic toxicity study of domoic acid in the rat. Food Chem. Toxicol. 1996, 34, 525–529. [Google Scholar] [CrossRef]
- Jeffery, B.; Barlow, T.; Moizer, K.; Paul, S.; Boyle, C. Amnesic shellfish poison. Food Chem. Toxicol. 2004, 42, 545–557. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific Opinion of the Panel on Contaminants in the Food Chain on a request from the European Commission on Marine biotoxins in shellfish–domoic acid. EFSA J. 2009, 1181, 1–61. [Google Scholar]
- CODEX. Joint Fao/Who Food Standards Programme. In Proceedings of the Codex Committee on Fish and Fishery Products, Twenty-Eighth Session, Beijing, China, 18–22 September 2006. [Google Scholar]
- Vieira, A.C.; Martinez, J.M.; Pose, R.B.; Queijo, A.A.; Posadas, N.A.; Lopez, L.M. Dose-response and histopathological study, with special attention to the hypophysis, of the differential effects of domoic acid on rats and mice. Microsc. Res. Tech. 2015, 78, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, T.; Mazet, J.A.; Zabka, T.S.; Langlois, G.; Colegrove, K.M.; Silver, M.; Bargu, S.; Van Dolah, F.; Leighfield, T.; Conrad, P.A.; et al. Novel symptomatology and changing epidemiology of domoic acid toxicosis in California sea lions (Zalophus californianus): An increasing risk to marine mammal health. Proc. Biol. Sci. 2008, 275, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Doucette, T.A.; Andrew Tasker, R. Perinatal Domoic Acid as a Neuroteratogen. Curr. Top. Behav. Neurosci. 2016, 29, 87–110. [Google Scholar] [PubMed]
- Grattan, L.M.; Holobaugh, S.; Morris, J.G., Jr. Harmful Algal Blooms and Public Health. Harmful Algae 2016, 57, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Grattan, L.M.; Boushey, C.; Tracy, K.; Trainer, V.; Roberts, S.M.; Schluterman, N.; Morris, J.G., Jr. The association between razor clam consumption and memory in the CoASTAL Cohort. Harmful Algae 2016, 57, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Bjornebekk, A.; Westlye, L.T.; Walhovd, K.B.; Fjell, A.M. Everyday memory: Self-perception and structural brain correlates in a healthy elderly population. J. Int. Neuropsychol. Soc. 2010, 16, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Louzao, M.C.; Abal, P.; Vilariño, N. Toxicity equivalence factor for regulated and non-regulated marine toxins. Curr. Opin. Food Sci. 2017, 18, 64–70. [Google Scholar] [CrossRef]
- Bagnis, R.; Chanteau, S.; Chungue, E.; Hurtel, J.M.; Yasumoto, T.; Inoue, A. Origins of ciguatera fish poisoning: A new dinoflagellate, Gambierdiscus toxicus Adachi and Fukuyo, definitively involved as a causal agent. Toxicon 1980, 18, 199–208. [Google Scholar] [CrossRef]
- Dickey, R.W.; Plakas, S.M. Ciguatera: A public health perspective. Toxicon 2010, 56, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Litaker, R.W.; Holland, W.C.; Hardison, D.R.; Pisapia, F.; Hess, P.; Kibler, S.R.; Tester, P.A. Ciguatoxicity of Gambierdiscus and Fukuyoa species from the Caribbean and Gulf of Mexico. PLoS ONE 2017, 12, e0185776. [Google Scholar] [CrossRef] [PubMed]
- Gatti, C.M.I.; Lonati, D.; Darius, H.T.; Zancan, A.; Roue, M.; Schicchi, A.; Locatelli, C.A.; Chinain, M. Tectus niloticus (Tegulidae, Gastropod) as a Novel Vector of Ciguatera Poisoning: Clinical Characterization and Follow-Up of a Mass Poisoning Event in Nuku Hiva Island (French Polynesia). Toxins 2018, 10, 102. [Google Scholar] [CrossRef] [PubMed]
- Lehane, L.; Lewis, R.J. Ciguatera: Recent advances but the risk remains. Int. J. Food Microbiol. 2000, 61, 91–125. [Google Scholar] [CrossRef]
- Skinner, M.P.; Brewer, T.D.; Johnstone, R.; Fleming, L.E.; Lewis, R.J. Ciguatera fish poisoning in the Pacific Islands (1998 to 2008). PLoS Negl. Trop. Dis. 2011, 5, e1416. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific Opinion on Marine biotoxins in shellfish–Emerging toxins: Ciguatoxin group. EFSA J. 2010, 8, 1–38. [Google Scholar]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An Updated Review of Ciguatera Fish Poisoning: Clinical, Epidemiological, Environmental, and Public Health Management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Bagnis, R.; Kuberski, T.; Laugier, S. Clinical observations on 3009 cases of ciguatera (fish poisoning) in the South Pacific. Am. J. Trop. Med. Hyg. 1979, 28, 1067–1073. [Google Scholar] [CrossRef] [PubMed]
- Quod, J.P.; Turquet, J. Ciguatera in Reunion Island (SW Indian Ocean): Epidemiology and clinical patterns. Toxicon 1996, 34, 779–785. [Google Scholar] [CrossRef]
- Tester, P.A.; Feldman, R.L.; Nau, A.W.; Kibler, S.R.; Wayne Litaker, R. Ciguatera fish poisoning and sea surface temperatures in the Caribbean Sea and the West Indies. Toxicon 2010, 56, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Bentur, Y.; Spanier, E. Ciguatoxin-like substances in edible fish on the eastern Mediterranean. Clin. Toxicol. 2007, 45, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Boada, L.D.; Zumbado, M.; Luzardo, O.P.; Almeida-Gonzalez, M.; Plakas, S.M.; Granade, H.R.; Abraham, A.; Jester, E.L.; Dickey, R.W. Ciguatera fish poisoning on the West Africa Coast: An emerging risk in the Canary Islands (Spain). Toxicon 2010, 56, 1516–1519. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.Y. Ciguatera fish poisoning in East Asia and Southeast Asia. Mar. Drugs 2015, 13, 3466–3478. [Google Scholar] [CrossRef] [PubMed]
- Otero, P.; Perez, S.; Alfonso, A.; Vale, C.; Rodriguez, P.; Gouveia, N.N.; Gouveia, N.; Delgado, J.; Vale, P.; Hirama, M.; et al. First toxin profile of ciguateric fish in Madeira Arquipelago (Europe). Anal. Chem. 2010, 82, 6032–6039. [Google Scholar] [CrossRef] [PubMed]
- Perez-Arellano, J.L.; Luzardo, O.P.; Perez Brito, A.; Hernandez Cabrera, M.; Zumbado, M.; Carranza, C.; Angel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera fish poisoning, Canary Islands. Emerg. Infect. Dis. 2005, 11, 1981–1982. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Sato, S.; Tawong, W.; Sakanari, H.; Uehara, K.; Shah, M.M.; Suda, S.; Yasumoto, T.; Taira, Y.; Yamaguchi, H.; et al. Genetic diversity and distribution of the ciguatera-causing dinoflagellate Gambierdiscus spp. (Dinophyceae) in coastal areas of Japan. PLoS ONE 2013, 8, e60882. [Google Scholar] [CrossRef] [PubMed]
- Kibler, S.R.; Tester, P.A.; Kunkel, K.E.; Moore, S.K.; Litaker, R.W. Effects of ocean warming on growth and distribution of dinoflagellates associated with ciguatera fish poisoning in the Caribbean. Ecol. Model. 2015, 316, 194–210. [Google Scholar] [CrossRef]
- Bidard, J.N.; Vijverberg, H.P.; Frelin, C.; Chungue, E.; Legrand, A.M.; Bagnis, R.; Lazdunski, M. Ciguatoxin is a novel type of Na+ channel toxin. J. Biol. Chem. 1984, 259, 8353–8357. [Google Scholar] [PubMed]
- Nicholson, G.M.; Lewis, R.J. Ciguatoxins: Cyclic polyether modulators of voltage-gated ion channel function. Mar. Drugs 2006, 4, 82–118. [Google Scholar] [CrossRef]
- Gillespie, N.C.; Lewis, R.J.; Pearn, J.H.; Bourke, A.T.; Holmes, M.J.; Bourke, J.B.; Shields, W.J. Ciguatera in Australia. Occurrence, clinical features, pathophysiology and management. Med. J. Aust. 1986, 145, 584–590. [Google Scholar] [PubMed]
- Lewis, R.J. The changing face of ciguatera. Toxicon 2001, 39, 97–106. [Google Scholar] [CrossRef]
- Ruff, T.A.; Lewis, R.J. Clinical aspects of ciguatera: An overview. Pac. Health Dialog 1994, 4, 119–127. [Google Scholar]
- Boucaud-Maitre, D.; Vernoux, J.P.; Pelczar, S.; Daudens-Vaysse, E.; Aubert, L.; Boa, S.; Ferracci, S.; Garnier, R. Incidence and clinical characteristics of ciguatera fish poisoning in Guadeloupe (French West Indies) between 2013 and 2016: A retrospective cases-series. Sci. Rep. 2018, 8, 3095. [Google Scholar] [CrossRef] [PubMed]
- DeFusco, D.J.; O’Dowd, P.; Hokama, Y.; Ott, B.R. Coma due to ciguatera poisoning in Rhode Island. Am. J. Med. 1993, 95, 240–243. [Google Scholar] [CrossRef]
- Karalis, T.; Gupta, L.; Chu, M.; Campbell, B.A.; Capra, M.F.; Maywood, P.A. Three clusters of ciguatera poisoning: Clinical manifestations and public health implications. Med. J. Aust. 2000, 172, 160–162. [Google Scholar] [PubMed]
- Chateau-Degat, M.L.; Dewailly, E.; Cerf, N.; Nguyen, N.L.; Huin-Blondey, M.O.; Hubert, B.; Laudon, F.; Chansin, R. Temporal trends and epidemiological aspects of ciguatera in French Polynesia: A 10-year analysis. Trop. Med. Int. Health 2007, 12, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Katz, A.R.; Terrell-Perica, S.; Sasaki, D.M. Ciguatera on Kauai: Investigation of factors associated with severity of illness. Am. J. Trop. Med. Hyg. 1993, 49, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Baumann, F.; Bourrat, M.B.; Pauillac, S. Prevalence, symptoms and chronicity of ciguatera in New Caledonia: Results from an adult population survey conducted in Noumea during 2005. Toxicon 2010, 56, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Pearn, J. Neurology of ciguatera. J. Neurol. Neurosurg. Psychiatry 2001, 70, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Pierre, O.; Misery, L.; Talagas, M.; Le Garrec, R. Immune effects of the neurotoxins ciguatoxins and brevetoxins. Toxicon 2018, 149, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.A.; Fleming, L.E.; Fernandez, M.; Bienfang, P.; Schrank, K.; Dickey, R.; Bottein, M.Y.; Backer, L.; Ayyar, R.; Weisman, R.; et al. Ciguatera fish poisoning: Treatment, prevention and management. Mar. Drugs 2008, 6, 456–479. [Google Scholar] [CrossRef] [PubMed]
- Bottein, M.Y.; Wang, Z.; Ramsdell, J.S. Toxicokinetics of the ciguatoxin P-CTX-1 in rats after intraperitoneal or oral administration. Toxicology 2011, 284, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ledreux, A.; Ramsdell, J.S. Bioavailability and intravenous toxicokinetic parameters for Pacific ciguatoxin P-CTX-1 in rats. Toxicon 2013, 64, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Mullins, M.E.; Hoffman, R.S. Is mannitol the treatment of choice for patients with ciguatera fish poisoning? Clin. Toxicol. 2017, 55, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, B.; Whittle, N.; Shaw, G.; Eaglesham, G.; Moore, M.R.; Lewis, R.J. Human fatality associated with Pacific ciguatoxin contaminated fish. Toxicon 2010, 56, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Vernoux, J.P.; Lewis, R.J. Isolation and characterisation of Caribbean ciguatoxins from the horse-eye jack (Caranx latus). Toxicon 1997, 35, 889–900. [Google Scholar] [CrossRef]
- CDC. Cluster of ciguatera fish poisoning—North Carolina, 2007. Weekly 2009, 58, 283–285. [Google Scholar]
- Lewis, R.J.; Hoy, A.W. Comparative action of three major ciguatoxins on guinea-pig atria and ilea. Toxicon 1993, 31, 437–446. [Google Scholar] [CrossRef]
- Terao, K.; Ito, E.; Yasumoto, T. Light and electron microscopic studies of the murine heart after repeated administrations of ciguatoxin or ciguatoxin-4c. Nat. Toxins 1992, 1, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Bottein Dechraoui, M.Y.; Rezvani, A.H.; Gordon, C.J.; Levin, E.D.; Ramsdell, J.S. Repeat exposure to ciguatoxin leads to enhanced and sustained thermoregulatory, pain threshold and motor activity responses in mice: Relationship to blood ciguatoxin concentrations. Toxicology 2008, 246, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Terao, K.; Ito, E.; Oarada, M.; Ishibashi, Y.; Legrand, A.M.; Yasumoto, T. Light and electron microscopic studies of pathologic changes induced in mice by ciguatoxin poisoning. Toxicon 1991, 29, 633–643. [Google Scholar] [CrossRef]
- Lewis, R.J.; Sellin, M.; Poli, M.A.; Norton, R.S.; MacLeod, J.K.; Sheil, M.M. Purification and characterization of ciguatoxins from moray eel (Lycodontis javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar] [CrossRef]
- Dickey, R.W. Ciguatera toxins: Chemistry, Toxicology and Detection. In Seafood and Freshwater Toxins; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 479–500. [Google Scholar]
- Satake, M.; Fukui, M.; Legrand, A.M.; Cruchet, P.; Yasumoto, T. Isolation and structures of new ciguatoxin analogs, 2,3-dihydroxyCTX3C and 51-hydroxyCTX3C, accumulated in tropical reef fish. Tetrahedron Lett. 1998, 39, 1197–1198. [Google Scholar] [CrossRef]
- Satake, M.; Ishibashi, Y.; Legrand, A.M.; Yasumoto, T. Isolation and structure of ciguatoxin-4A, a new ciguatoxin precursor, from cultures of dinoflagellate Gambierdiscus toxicus and parrotfish Scarus gibbus. Biosci. Biotechnol. Biochem. 1997, 60, 2103–2105. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Yasumoto, T. Structures and configurations of ciguatoxin from the moray eel Gymnothorax javanicus and its likely precursor from the dinoflaggelate Gambierdiscus toxicus. J. Am. Chem. Soc. 1990, 112, 4380–4386. [Google Scholar] [CrossRef]
- Lewis, R.J.; Vernoux, J.P.; Brereton, I.M. Structure of Caribbean ciguatoxin isolated from Caranx latus. J. Am. Chem. Soc. 1998, 120, 5914–5920. [Google Scholar] [CrossRef]
- FDA. Fish and Fishery Products. Hazards and Controls Guidance, 4th ed.; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2011.
- Norma Oficial Mexicana NOM-242-SSA1-2009. Productos y servicios. Productos de la pesca frescos, refrigerados, congelados y procesados. Especificaciones sanitarias y métodos de prueba. Diario Oficial de la Federación. Mexico. Off. J. Eur. Commun. 2011, L221, 29. [Google Scholar]
- Suzuki, T. Marine Toxin Research and Toxin Monitoring System in Japan: Experiences from Japan. Available online: http://iocwestpac.org/files/upload_manual/L1%20Suzuki.pdf (accessed on 8 August 2018).
- Turner, A.D.; Higgins, C.; Davidson, K.; Veszelovszki, A.; Payne, D.; Hungerford, J.; Higman, W. Potential threats posed by new or emerging marine biotoxins in UK waters and examination of detection methodology used in their control: Brevetoxins. Mar. Drugs 2015, 13, 1224–1254. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific Opinion of the Panel on Contaminants in the Food Chain on a request from the European Commission on Marine biotoxins in shellfish–Emerging toxins: Brevetoxin group. EFSA J. 2010, 8, 1–29. [Google Scholar]
- Watkins, S.M.; Reich, A.; Fleming, L.E.; Hammond, R. Neurotoxic shellfish poisoning. Mar. Drugs 2008, 6, 431–455. [Google Scholar] [CrossRef] [PubMed]
- Pierce, R.H.; Henry, M.S. Harmful algal toxins of the Florida red tide (Karenia brevis): Natural chemical stressors in South Florida coastal ecosystems. Ecotoxicology 2008, 17, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Fleming, L.E.; Kirkpatrick, B.; Backer, L.C.; Walsh, C.J.; Nierenberg, K.; Clark, J.; Reich, A.; Hollenbeck, J.; Benson, J.; Cheng, Y.S.; et al. Review of Florida Red Tide and Human Health Effects. Harmful Algae 2011, 10, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Atchison, W.D.; Luke, V.S.; Narahashi, T.; Vogel, S.M. Nerve membrane sodium channels as the target site of brevetoxins at neuromuscular junctions. Br. J. Pharmacol. 1986, 89, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Poli, M.A.; Musser, S.M.; Dickey, R.W.; Eilers, P.P.; Hall, S. Neurotoxic shellfish poisoning and brevetoxin metabolites: A case study from Florida. Toxicon 2000, 38, 981–993. [Google Scholar] [CrossRef]
- Morabito, S.; Silvestro, S.; Faggio, C. How the marine biotoxins affect human health. Nat. Prod. Res. 2018, 32, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Poli, M.A.; Templeton, C.B.; Thompson, W.L.; Hewetson, J.F. Distribution and elimination of brevetoxin PbTx-3 in rats. Toxicon 1990, 28, 903–910. [Google Scholar] [CrossRef]
- Fleming, L.E.; Backer, L.C.; Baden, D.G. Overview of aerosolized Florida red tide toxins: Exposures and effects. Environ. Health Perspect. 2005, 113, 618–620. [Google Scholar] [CrossRef] [PubMed]
- Bean, J.A.; Fleming, L.E.; Kirkpatrick, B.; Backer, L.C.; Nierenberg, K.; Reich, A.; Cheng, Y.S.; Wanner, A.; Benson, J.; Naar, J.; et al. Florida Red Tide Toxins (Brevetoxins) and Longitudinal Respiratory Effects in Asthmatics. Harmful Algae 2011, 10, 744–748. [Google Scholar] [CrossRef] [PubMed]
- Benson, J.M.; Tischler, D.L.; Baden, D.G. Uptake, tissue distribution, and excretion of brevetoxin 3 administered to rats by intratracheal instillation. J. Toxicol. Environ Health 1999, 57, 345–355. [Google Scholar]
- Bourdelais, A.J.; Campbell, S.; Jacocks, H.; Naar, J.; Wright, J.L.; Carsi, J.; Baden, D.G. Brevenal is a natural inhibitor of brevetoxin action in sodium channel receptor binding assays. Cell. Mol. Neurobiol. 2004, 24, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, H. Joint FAO/WHO/IOC activities to provide scientific advice on marine biotoxins (research report). Mar. Pollut. Bull. 2006, 52, 1735–1745. [Google Scholar] [CrossRef] [PubMed]
- Zaias, J.; Fleming, L.E.; Baden, D.G.; Abraham, W.M. Repeated exposure to aerosolized brevetoxin-3 induces prolonged airway hyperresponsiveness and lung inflammation in sheep. Inhal. Toxicol. 2011, 23, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Morohashi, A.; Satake, M.; Naoki, H.; Kaspar, H.F.; Oshima, Y.; Yasumoto, T. Brevetoxin B4 isolated from greenshell mussels Perna canaliculus, the major toxin involved in neurotoxic shellfish poisoning in New Zealand. Nat. Toxins 1999, 7, 45–48. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on the risks for public health related to the presence of tetrodotoxin (TTX) and TTX analogues in marine bivalves and gastropods. EFSA J. 2017, 15, 4752. [Google Scholar]
- Noguchi, T.; Onuki, K.; Arakawa, O. Tetrodotoxin poisoning due to pufferfish and gastropods, and their intoxication mechanism. ISRN Toxicol. 2011, 2011, 276939. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.; Alfonso, A.; Vale, C.; Alfonso, C.; Vale, P.; Tellez, A.; Botana, L.M. First toxicity report of tetrodotoxin and 5,6,11-trideoxyTTX in the trumpet shell Charonia lampas lampas in Europe. Anal. Chem. 2008, 80, 5622–5629. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Eurosurveillance 2015, 20, 21009. [Google Scholar] [CrossRef] [PubMed]
- Lipkind, G.M.; Fozzard, H.A. A structural model of the tetrodotoxin and saxitoxin binding site of the Na+ channel. Biophys. J. 1994, 66, 1–13. [Google Scholar] [CrossRef]
- Narahashi, T. Chemicals as tools in the study of excitable membranes. Physiol. Rev. 1974, 54, 813–889. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.J.; Schild, L.; Moczydlowski, E.G. Use-dependent block of the voltage-gated Na+ channel by tetrodotoxin and saxitoxin: Effect of pore mutations that change ionic selectivity. J. Gen. Physiol. 2012, 140, 435–454. [Google Scholar] [CrossRef] [PubMed]
- Lago, J.; Rodriguez, L.P.; Blanco, L.; Vieites, J.M.; Cabado, A.G. Tetrodotoxin, an Extremely Potent Marine Neurotoxin: Distribution, Toxicity, Origin and Therapeutical Uses. Mar. Drugs 2015, 13, 6384–6406. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, M.A.; Schneider, J.J.; Isbister, G.K. Use of high performance liquid chromatography to measure tetrodotoxin in serum and urine of poisoned patients. Toxicon 2004, 44, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Fong, B.M.; Tam, S.; Tsui, S.H.; Leung, K.S. Development and validation of a high-throughput double solid phase extraction-liquid chromatography-tandem mass spectrometry method for the determination of tetrodotoxin in human urine and plasma. Talanta 2011, 83, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Abal, P.; Louzao, M.C.; Antelo, A.; Alvarez, M.; Cagide, E.; Vilarino, N.; Vieytes, M.R.; Botana, L.M. Acute Oral Toxicity of Tetrodotoxin in Mice: Determination of Lethal Dose 50 (LD50) and No Observed Adverse Effect Level (NOAEL). Toxins 2017, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Mattei, C. Tetrodotoxin, a Candidate Drug for Nav1.1-Induced Mechanical Pain? Mar. Drugs 2018, 16, 72. [Google Scholar] [CrossRef] [PubMed]
- Valdiglesias, V.; Prego-Faraldo, M.V.; Pasaro, E.; Mendez, J.; Laffon, B. Okadaic acid: More than a diarrheic toxin. Mar. Drugs 2013, 11, 4328–4349. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a New Type of Shellfish Poisoning in the Tohoku District. Nippon Suisan Gakkaishi 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- Braga, A.C.; Alves, R.N.; Maulvault, A.L.; Barbosa, V.; Marques, A.; Costa, P.R. In vitro bioaccessibility of the marine biotoxin okadaic acid in shellfish. Food Chem. Toxicol. 2016, 89, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Doucet, E.; Ross, N.N.; Quilliam, M.A. Enzymatic hydrolysis of esterified diarrhetic shellfish poisoning toxins and pectenotoxins. Anal. Bioanal. Chem. 2007, 389, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Bialojan, C.; Takai, A. Inhibitory effect of a marine-sponge toxin, okadaic acid, on protein phosphatases. Specificity and kinetics. Biochem. J. 1988, 256, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Twiner, M.J.; Doucette, G.J.; Pang, Y.; Fang, C.; Forsyth, C.J.; Miles, C.O. Structure-Activity Relationship Studies Using Natural and Synthetic Okadaic Acid/Dinophysistoxin Toxins. Mar. Drugs 2016, 14, 207. [Google Scholar] [CrossRef] [PubMed]
- MacKintosh, C.; Klumpp, S. Tautomycin from the bacterium Streptomyces verticillatus. Another potent and specific inhibitor of protein phosphatases 1 and 2A. FEBS Lett. 1990, 277, 137–140. [Google Scholar] [CrossRef]
- Munday, R. Is protein phosphatase inhibition responsible for the toxic effects of okadaic Acid in animals? Toxins 2013, 5, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Louzao, M.C.; Fernandez, D.A.; Abal, P.; Fraga, M.; Vilarino, N.; Vieytes, M.R.; Botana, L.M. Diarrhetic effect of okadaic acid could be related with its neuronal action: Changes in neuropeptide Y. Toxicol. Lett. 2015, 237, 151–160. [Google Scholar] [CrossRef] [PubMed]
- James, K.J.; Carey, B.; O’Halloran, J.; van Pelt, F.N.; Skrabakova, Z. Shellfish toxicity: Human health implications of marine algal toxins. Epidemiol. Infect. 2010, 138, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Trainer, V.L.; Moore, L.; Bill, B.D.; Adams, N.G.; Harrington, N.; Borchert, J.; da Silva, D.A.; Eberhart, B.T. Diarrhetic shellfish toxins and other lipophilic toxins of human health concern in Washington State. Mar. Drugs 2013, 11, 1815–1835. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, D.A.; Louzao, M.C.; Fraga, M.; Vilarino, N.; Vieytes, M.R.; Botana, L.M. Experimental basis for the high oral toxicity of dinophysistoxin 1: A comparative study of DSP. Toxins 2014, 6, 211–228. [Google Scholar] [CrossRef] [PubMed]
- Abal, P.; Louzao, M.C.; Suzuki, T.; Watanabe, R.; Vilariño, N.; Carrera, C.; Botana, A.M.; Vieytes, M.R.; Botana, L.M. Toxic action reevaluation of Okadaic Acid, Dinophysistoxin-1 and Dinophysistoxin-2: Toxicity Equivalency Factors based on the oral toxicity study. Cell. Physiol. Biochem. 2018, in press. [Google Scholar]
- Garcia, C.; Truan, D.; Lagos, M.; Santelices, J.P.; Diaz, J.C.; Lagos, N. Metabolic transformation of dinophysistoxin-3 into dinophysistoxin-1 causes human intoxication by consumption of O-acyl-derivatives dinophysistoxins contaminated shellfish. J. Toxicol. Sci. 2005, 30, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Matias, W.G.; Traore, A.; Creppy, E.E. Variations in the distribution of okadaic acid in organs and biological fluids of mice related to diarrhoeic syndrome. Hum. Exp. Toxicol. 1999, 18, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Abal, P.; Louzao, M.C.; Cifuentes, J.M.; Vilarino, N.; Rodriguez, I.; Alfonso, A.; Vieytes, M.R.; Botana, L.M. Characterization of the dinophysistoxin-2 acute oral toxicity in mice to define the Toxicity Equivalency Factor. Food Chem. Toxicol. 2017, 102, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Matias, W.G.; Creppy, E.E. Transplacental passage of [3H]-okadaic acid in pregnant mice measured by radioactivity and high-performance liquid chromatography. Hum. Exp. Toxicol. 1996, 15, 226–230. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Opinion of the Scientific Panel on Contaminants in the Food chain on a request from the European Commission Marine biotoxins in shellfish–okadaic acid and analogues. EFSA J. 2008, 589, 1–62. [Google Scholar]
- Le Hegarat, L.; Jacquin, A.G.; Bazin, E.; Fessard, V. Genotoxicity of the marine toxin okadaic acid, in human Caco-2 cells and in mice gut cells. Environ. Toxicol. 2006, 21, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Valdiglesias, V.; Mendez, J.; Pasaro, E.; Cemeli, E.; Anderson, D.; Laffon, B. Assessment of okadaic acid effects on cytotoxicity, DNA damage and DNA repair in human cells. Mutat. Res. 2010, 689, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Suganuma, M.; Suguri, H.; Yoshizawa, S.; Takagi, K.; Uda, N.; Wakamatsu, K.; Yamada, K.; Murata, M.; Yasumoto, T.; et al. Diarrhetic shellfish toxin, dinophysistoxin-1, is a potent tumor promoter on mouse skin. Jpn. J. Cancer Res. 1988, 79, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Sueoka, E.; Suganuma, M. Tumor promoters: From chemicals to inflammatory proteins. J. Cancer Res. Clin. Oncol. 2013, 139, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, M.; Zhong, T.Y.; Tampe, R.; Garcia, L.; Lagos, N. Sublethal doses of dinophysistoxin-1 and okadaic acid stimulate secretion of inflammatory factors on innate immune cells: Negative health consequences. Toxicon 2017, 126, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, M.; Toledo, H.; Lagos, N. Okadaic acid toxin at sublethal dose produced cell proliferation in gastric and colon epithelial cell lines. Mar. Drugs 2013, 11, 4751–4760. [Google Scholar] [CrossRef] [PubMed]
- Manerio, E.; Rodas, V.; Costas, E.; Hernandez, J. Shellfish consumption: A major risk factor for colorectal cancer. Med. Hypotheses 2008, 70, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Cordier, S.; Monfort, C.; Miossec, L.; Richardson, S.; Belin, C. Ecological analysis of digestive cancer mortality related to contamination by diarrhetic shellfish poisoning toxins along the coasts of France. Environ. Res. 2000, 84, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rodas, V.; Maneiro, E.; Martinez, J.; Navarro, M.; Costas, E. Harmful algal blooms, red tides and human health: Diarrhetic shellfish poisoning and colorectal cancer. Anal. Real Acad. Nac. Farm. 2006, 72, 391–408. [Google Scholar]
- EC. Commission regulation (EU) No. 15/2011 of 10 January 2011 amending Regulation (EC) No. 2074/2005 as regards recognised testing methods for detecting marine biotoxins in live bivalve molluscs. Off. J. Eur. Commun. 2011, L6, 3–9. [Google Scholar]
- Aune, T.; Larsen, S.; Aasen, J.A.; Rehmann, N.; Satake, M.; Hess, P. Relative toxicity of dinophysistoxin-2 (DTX-2) compared with okadaic acid, based on acute intraperitoneal toxicity in mice. Toxicon 2007, 49, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tubaro, A.; Sosa, S.; Bornacin, A.; Jungerford, J. Pharmacology and toxicology of diarrheic shellfish toxins. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection, 2nd ed.; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 229–253. [Google Scholar]
- EFSA. Scientific Opinion of the Panel on Contaminants in the Food Chain on a request from the European Commission on Marine Biotoxins in Shellfish—Summary on regulated marine biotoxins. EFSA J. 2009, 1306, 1–23. [Google Scholar]
- Aune, T.; Espenes, A.; Aasen, J.A.; Quilliam, M.A.; Hess, P.; Larsen, S. Study of possible combined toxic effects of azaspiracid-1 and okadaic acid in mice via the oral route. Toxicon 2012, 60, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.C.; Rubiolo, J.A.; Lopez-Alonso, H.; Cifuentes, J.M.; Alfonso, A.; Bermudez, R.; Otero, P.; Vieytes, M.R.; Vega, F.V.; Botana, L.M. Oral toxicity of okadaic acid in mice: Study of lethality, organ damage, distribution and effects on detoxifying gene expression. Toxins 2013, 5, 2093–2108. [Google Scholar] [CrossRef] [PubMed]
- Hopson, K.P.; Truelove, J.; Chun, J.; Wang, Y.; Waeber, C. S1P activates store-operated calcium entry via receptor- and non-receptor-mediated pathways in vascular smooth muscle cells. Am. J. Physiol. Cell Physiol. 2011, 300, C919–C926. [Google Scholar] [CrossRef] [PubMed]
- BIOTOXINS. EU-Harmonised Standard Operating Procedure for determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS Version 5; Agencia Española de Consumo, Seguridad Alimentaria y Nutrición: Madrid, Spain, 2015. [Google Scholar]
- Kilcoyne, J.; McCarron, P.; Twiner, M.J.; Rise, F.; Hess, P.; Wilkins, A.L.; Miles, C.O. Identification of 21,22-Dehydroazaspiracids in Mussels (Mytilus edulis) and in Vitro Toxicity of Azaspiracid-26. J. Nat. Prod. 2018, 81, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Ofuji, K.; Naoki, H.; James, K.J.; Furey, A.; McMahon, T.; Silke, J.; Yasumoto, T. Azaspiracid, a new marine toxin having unique spiro ring assemblies, isolated from Irish mussels, Mytilus edulis. J. Am. Chem. Soc. 1998, 120, 9967–9968. [Google Scholar] [CrossRef]
- Tillmann, U.; Jaen, D.; Fernandez, L.; Gottschling, M.; Witt, M.; Blanco, J.; Krock, B. Amphidoma languida (Amphidomatacea, Dinophyceae) with a novel azaspiracid toxin profile identified as the cause of molluscan contamination at the Atlantic coast of southern Spain. Harmful Algae 2017, 62, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Tillmann, U.; Borel, C.M.; Barrera, F.; Lara, R.; Krock, B.; Almandoz, G.O.; Witt, M.; Trefault, N. Azadinium poporum from the Argentine Continental Shelf, Southwestern Atlantic, produces azaspiracid-2 and azaspiracid-2 phosphate. Harmful Algae 2016, 51, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Bacchiocchi, S.; Siracusa, M.; Ruzzi, A.; Gorbi, S.; Ercolessi, M.; Cosentino, M.A.; Ammazzalorso, P.; Orletti, R. Two-year study of lipophilic marine toxin profile in mussels of the North-central Adriatic Sea: First report of azaspiracids in Mediterranean seafood. Toxicon 2015, 108, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rivera, A.; O’Callaghan, K.; Moriarty, M.; O’Driscoll, D.; Hamilton, B.; Lehane, M.; James, K.J.; Furey, A. First evidence of azaspiracids (AZAs): A family of lipophilic polyether marine toxins in scallops (Argopecten purpuratus) and mussels (Mytilus chilensis) collected in two regions of Chile. Toxicon 2010, 55, 692–701. [Google Scholar] [CrossRef] [PubMed]
- James, K.J.; Furey, A.; Lehane, M.; Ramstad, H.; Aune, T.; Hovgaard, P.; Morris, S.; Higman, W.; Satake, M.; Yasumoto, T. First evidence of an extensive northern European distribution of azaspiracid poisoning (AZP) toxins in shellfish. Toxicon 2002, 40, 909–915. [Google Scholar] [CrossRef]
- Magdalena, A.B.; Lehane, M.; Krys, S.; Fernandez, M.L.; Furey, A.; James, K.J. The first identification of azaspiracids in shellfish from France and Spain. Toxicon 2003, 42, 105–108. [Google Scholar] [CrossRef]
- Klontz, K.C.; Abraham, A.; Plakas, S.M.; Dickey, R.W. Mussel-associated azaspiracid intoxication in the United States. Ann. Intern. Med. 2009, 150, 361. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Ofuji, K.; James, K.J.; Furey, A.; Yasumoto, T. New toxic event caused by Irish mussels. Harmful Algae 1997, 468–469. [Google Scholar]
- Taleb, H.; Vale, P.; Amanhir, R.; Benhadouch, A.; Sagou, R.; Chafik, A. First detection of azaspiracid in mussels in North West Africa. J. Shellfish Res. 2006, 25, 1067–1070. [Google Scholar]
- Ueoka, R.; Ito, A.; Izumikawa, M.; Maeda, S.; Takagi, M.; Shin-ya, K.; Yoshida, M.; van Soest, R.W.M.; Matsunaga, S. Isolation of azaspiracid-2 from a marine sponge Echinoclathria sp. as a potent cytotoxin. Toxicon 2009, 53, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Tillmann, U.; Adams, N.G.; Krock, B.; Stutts, W.L.; Deeds, J.R.; Han, M.S.; Trainer, V.L. Identification of Azadinium species and a new azaspiracid from Azadinium poporum in Puget Sound, Washington State, USA. Harmful Algae 2017, 68, 152–167. [Google Scholar] [CrossRef] [PubMed]
- McMahon, T.S.J. Winter toxicity of unknown aetiology in mussels. Harmful Algae 1996, 14, 2. [Google Scholar]
- Álvarez, G.; Uribe, E.; Avalos, P.; Marino, C.; Blanco, J. First identification of azaspiracid and spirolides in Mesodesma donacium and Mulinia edulis from Northern Chile. Toxicon 2010, 55, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Furey, A.; Moroney, C.; Braña-Magdalena, A.; Saez, M.J.; Lehane, M.; James, K.J. Geographical, temporal, and species variation of the polyether toxins, azaspiracids, in shellfish. Environ. Sci. Technol. 2003, 37, 3078–3084. [Google Scholar] [CrossRef] [PubMed]
- McMahon, T.; Silke, J. Re-occurrence of winter toxicity. Harmful Algae News, 1998; 17, 12. [Google Scholar]
- EFSA. Opinion of the Scientific Panel on Contaminants in the Food chain on a request from the European Commission Marine biotoxins in shellfish—Azaspiracids. EFSA J. 2008, 723, 1–52. [Google Scholar]
- James, K.J.; Fidalgo Saez, M.J.; Furey, A.; Lehane, M. Azaspiracid poisoning, the food-borne illness associated with shellfish consumption. Food Addit. Contam. 2004, 21, 879–892. [Google Scholar] [CrossRef] [PubMed]
- RASFF-Food and Feed Safety Alert Ref. 2008.0426-add11. European Commission, 2008. Available online: https://ec.europa.eu/food/safety/rasff/portal_en (accessed on 8 August 2018).
- RASFF-Food and Feed Safety Alert Ref. 2013.1596. European Commission, 2013. Available online: https://ec.europa.eu/food/safety/rasff/portal_en (accessed on 8 August 2018).
- RASFF-Food and Feed Safety Alert Ref. 2013.1706. European Commission, 2013. Available online: https://ec.europa.eu/food/safety/rasff/portal_en (accessed on 8 August 2018).
- Abal, P.; Louzao, M.C.; Fraga, M.; Vilarino, N.; Ferreiro, S.; Vieytes, M.R.; Botana, L.M. Absorption and Effect of Azaspiracid-1 Over the Human Intestinal Barrier. Cell. Physiol. Biochem. 2017, 43, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Aasen, J.A.; Espenes, A.; Hess, P.; Aune, T. Sub-lethal dosing of azaspiracid-1 in female NMRI mice. Toxicon 2010, 56, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Amzil, Z.; Sibat, M.; Royer, F.; Savar, V. First report on azaspiracid and yessotoxin groups detection in French shellfish. Toxicon 2008, 52, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mendoza, E.; Sanchez-Bravo, Y.A.; Turner, A.; Blanco, J.; O’Neil, A.; Mancera-Flores, J.; Perez-Brunius, P.; Rivas, D.; Almazan-Becerril, A.; Pena-Manjarrez, J.L. Lipophilic toxins in cultivated mussels (Mytilus galloprovincialis) from Baja California, Mexico. Toxicon 2014, 90, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Elgarch, A.; Vale, P.; Rifai, S.; Fassouane, A. Detection of diarrheic shellfish poisoning and azaspiracid toxins in Moroccan mussels: Comparison of the LC-MS method with the commercial immunoassay kit. Mar. Drugs 2008, 6, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Vale, P. Is there a risk of human poisoning by azaspiracids from shellfish harvested at the Portuguese coast? Toxicon 2004, 44, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Ito, E.; Satake, M.; Ofuji, K.; Higashi, M.; Harigaya, K.; McMahon, T.; Yasumoto, T. Chronic effects in mice caused by oral administration of sublethal doses of azaspiracid, a new marine toxin isolated from mussels. Toxicon 2002, 40, 193–203. [Google Scholar] [CrossRef]
- Doerr, B.; O’Halloran, J.; O’Brien, N.; van Pelt, F. Investigation of the genotoxic potential of the marine biotoxins azaspiracid 1–3. Toxicon 2016, 121, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Ferreiro, S.F.; Vilarino, N.; Carrera, C.; Louzao, M.C.; Cantalapiedra, A.G.; Santamarina, G.; Cifuentes, J.M.; Vieira, A.C.; Botana, L.M. Subacute Cardiovascular Toxicity of the Marine Phycotoxin Azaspiracid-1 in Rats. Toxicol. Sci. 2016, 151, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Ferreiro, S.F.; Vilarino, N.; Carrera, C.; Louzao, M.C.; Santamarina, G.; Cantalapiedra, A.G.; Cifuentes, J.M.; Crespo, A.; Botana, L.M. In vivo cardiomyocyte response to YTX- and AZA-1-induced damage: Autophagy versus apoptosis. Arch. Toxicol. 2017, 91, 1859–1870. [Google Scholar] [CrossRef] [PubMed]
- Pelin, M.; Kilcoyne, J.; Nulty, C.; Crain, S.; Hess, P.; Tubaro, A.; Sosa, S. Toxic equivalency factors (TEFs) after acute oral exposure of azaspiracid 1, −2 and −3 in mice. Toxicol. Lett. 2018, 282, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Jauffrais, T.; Marcaillou, C.; Herrenknecht, C.; Truquet, P.; Sechet, V.; Nicolau, E.; Tillmann, U.; Hess, P. Azaspiracid accumulation, detoxification and biotransformation in blue mussels (Mytilus edulis) experimentally fed Azadinium spinosum. Toxicon 2012, 60, 582–595. [Google Scholar] [CrossRef] [PubMed]
- McCarron, P.; Kilcoyne, J.; Miles, C.O.; Hess, P. Formation of Azaspiracids-3, -4, -6, and -9 via decarboxylation of carboxyazaspiracid metabolites from shellfish. J. Agric. Food Chem. 2009, 57, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Pelin, M.; Brovedani, V.; Sosa, S.; Tubaro, A. Palytoxin-Containing Aquarium Soft Corals as an Emerging Sanitary Problem. Mar. Drugs 2016, 14, 33. [Google Scholar] [CrossRef] [PubMed]
- Brissard, C.; Herve, F.; Sibat, M.; Sechet, V.; Hess, P.; Amzil, Z.; Herrenknecht, C. Characterization of ovatoxin-h, a new ovatoxin analog, and evaluation of chromatographic columns for ovatoxin analysis and purification. J. Chromatogr. A 2015, 1388, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Tartaglione, L.; Mazzeo, A.; Dell’Aversano, C.; Forino, M.; Giussani, V.; Capellacci, S.; Penna, A.; Asnaghi, V.; Faimali, M.; Chiantore, M.; et al. Chemical, molecular, and eco-toxicological investigation of Ostreopsis sp. from Cyprus Island: Structural insights into four new ovatoxins by LC-HRMS/MS. Anal. Bioanal. Chem. 2016, 408, 915–932. [Google Scholar] [CrossRef] [PubMed]
- Deeds, J.R.; Handy, S.M.; White, K.D.; Reimer, J.D. Palytoxin found in Palythoa sp. zoanthids (Anthozoa, Hexacorallia) sold in the home aquarium trade. PLoS ONE 2011, 6, e18235. [Google Scholar] [CrossRef] [PubMed]
- Aligizaki, K.; Katikou, P.; Nikolaidis, G.; Panou, A. First episode of shellfish contamination by palytoxin-like compounds from Ostreopsis species (Aegean Sea, Greece). Toxicon 2008, 51, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Bire, R.; Trotereau, S.; Lemee, R.; Oregioni, D.; Delpont, C.; Krys, S.; Guerin, T. Hunt for Palytoxins in a Wide Variety of Marine Organisms Harvested in 2010 on the French Mediterranean Coast. Mar. Drugs 2015, 13, 5425–5446. [Google Scholar] [CrossRef] [PubMed]
- Taniyama, S.; Arakawa, O.; Terada, M.; Nishio, S.; Takatani, T.; Mahmud, Y.; Noguchi, T. Ostreopsis sp., a possible origin of palytoxin (PTX) in parrotfish Scarus ovifrons. Toxicon 2003, 42, 29–33. [Google Scholar] [CrossRef]
- Amzil, Z.; Sibat, M.; Chomerat, N.; Grossel, H.; Marco-Miralles, F.; Lemee, R.; Nezan, E.; Sechet, V. Ovatoxin-a and palytoxin accumulation in seafood in relation to Ostreopsis cf. ovata blooms on the French Mediterranean coast. Mar. Drugs 2012, 10, 477–496. [Google Scholar] [CrossRef] [PubMed]
- Bire, R.; Trotereau, S.; Lemee, R.; Delpont, C.; Chabot, B.; Aumond, Y.; Krys, S. Occurrence of palytoxins in marine organisms from different trophic levles of the French Mediterranean coast harvested in 2009. Harmful Algae 2013, 28, 10–22. [Google Scholar] [CrossRef]
- Artigas, P.; Gadsby, D.C. Large diameter of palytoxin-induced Na/K pump channels and modulation of palytoxin interaction by Na/K pump ligands. J. Gen. Physiol. 2004, 123, 357–376. [Google Scholar] [CrossRef] [PubMed]
- Ares, I.R.; Cagide, E.; Louzao, M.C.; Espina, B.; Vieytes, M.R.; Yasumoto, T.; Botana, L.M. Ostreocin-d impact on globular actin of intact cells. Chem. Res. Toxicol. 2009, 22, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Bellocci, M.; Ronzitti, G.; Milandri, A.; Melchiorre, N.; Grillo, C.; Poletti, R.; Yasumoto, T.; Rossini, G.P. A cytolytic assay for the measurement of palytoxin based on a cultured monolayer cell line. Anal. Biochem. 2008, 374, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.L.; Yang, C.C.; Deng, J.F.; Wang, K.Y. Hyperkalemia, hyperphosphatemia, acute kidney injury, and fatal dysrhythmias after consumption of palytoxin-contaminated goldspot herring. Ann. Emerg. Med. 2014, 64, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Thakur, L.K.; Jha, K.K. Palytoxin-induced acute respiratory failure. Respir. Med. Case Rep. 2017, 20, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Wieringa, A.; Bertholee, D.; Ter Horst, P.; van den Brand, I.; Haringman, J.; Ciminiello, P. Respiratory impairment in four patients associated with exposure to palytoxin containing coral. Clin. Toxicol. 2014, 52, 150–151. [Google Scholar] [CrossRef] [PubMed]
- Sud, P.; Su, M.K.; Greller, H.A.; Majlesi, N.; Gupta, A. Case series: Inhaled coral vapor-toxicity in a tank. J. Med. Toxicol. 2013, 9, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Deeds, J.R.; Schwartz, M.D. Human risk associated with palytoxin exposure. Toxicon 2010, 56, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Taniyama, S.; Mahmud, Y.; Terada, M.; Takatani, T.; Arakawa, O.; Noguchi, T. Occurrence of a food poisoning incident by palytoxin from a serranid Epinephelus sp. in Japan. J. Nat. Toxins 2002, 11, 277–282. [Google Scholar] [PubMed]
- Onuma, Y.; Satake, M.; Ukena, T.; Roux, J.; Chanteau, S.; Rasolofonirina, N.; Ratsimaloto, M.; Naoki, H.; Yasumoto, T. Identification of putative palytoxin as the cause of clupeotoxism. Toxicon 1999, 37, 55–65. [Google Scholar] [CrossRef]
- Alcala, A.C.; Alcala, L.C.; Garth, J.S.; Yasumura, D.; Yasumoto, T. Human fatality due to ingestion of the crab Demania reynaudii that contained a palytoxin-like toxin. Toxicon 1988, 26, 105–107. [Google Scholar] [CrossRef]
- Bernasconi, M.; Berger, D.; Tamm, M.; Stolz, D. Aquarism: An innocent leisure activity? Palytoxin-induced acute pneumonitis. Respiration 2012, 84, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Hamade, A.K.; Deglin, S.E.; McLaughlin, J.B.; Deeds, J.R.; Handy, S.M.; Knolhoff, A.M. Suspected Palytoxin Inhalation Exposures Associated with Zoanthid Corals in Aquarium Shops and Homes–Alaska, 2012–2014. Morb. Mortal. Wkly. Rep. 2015, 64, 852–855. [Google Scholar] [CrossRef]
- Murphy, L.T.; Charlton, N.P. Prevalence and characteristics of inhalational and dermal palytoxin exposures reported to the National Poison Data System in the U.S. Environ. Toxicol. Pharmacol. 2017, 55, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Tartaglione, L.; Dell’Aversano, C.; Mazzeo, A.; Forino, M.; Wieringa, A.; Ciminiello, P. Determination of Palytoxins in Soft Coral and Seawater from a Home Aquarium. Comparison between Palythoa- and Ostreopsis-Related Inhalatory Poisonings. Environ. Sci. Technol. 2016, 50, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Durando, P.; Ansaldi, F.; Oreste, P.; Moscatelli, P.; Marensi, L.; Grillo, C.; Gasparini, R.; Icardi, G.; Collaborative Group for the Ligurian Syndromic Algal Surveillance. Ostreopsis ovata and human health: Epidemiological and clinical features of respiratory syndrome outbreaks from a two-year syndromic surveillance, 2005-06, in north-west Italy. Eurosurveillance 2007, 12, E070607. [Google Scholar] [CrossRef] [PubMed]
- Gallitelli, M.; Ungaro, N.; Addante, L.M.; Procacci, V.; Silveri, N.G.; Sabba, C. Respiratory illness as a reaction to tropical algal blooms occurring in a temperate climate. JAMA 2005, 293, 2599–2600. [Google Scholar] [PubMed]
- Tichadou, L.; Glaizal, M.; Armengaud, A.; Grossel, H.; Lemee, R.; Kantin, R.; Lasalle, J.L.; Drouet, G.; Rambaud, L.; Malfait, P.; et al. Health impact of unicellular algae of the Ostreopsis genus blooms in the Mediterranean Sea: Experience of the French Mediterranean coast surveillance network from 2006 to 2009. Clin. Toxicol. 2010, 48, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Benedettini, G.; Onorari, M.; Serena, F.; Battocchi, C.; et al. First finding of Ostreopsis cf. ovata toxins in marine aerosols. Environ. Sci. Technol. 2014, 48, 3532–3540. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pezzolesi, L.; Pistocchi, R.; et al. Isolation and structure elucidation of ovatoxin-a, the major toxin produced by Ostreopsis ovata. J. Am. Chem. Soc. 2012, 134, 1869–1875. [Google Scholar] [CrossRef] [PubMed]
- Nordt, S.P.; Wu, J.; Zahller, S.; Clark, R.F.; Cantrell, F.L. Palytoxin poisoning after dermal contact with zoanthid coral. J. Emerg. Med. 2011, 40, 397–399. [Google Scholar] [CrossRef] [PubMed]
- Farooq, A.V.; Gibbons, A.G.; Council, M.D.; Harocopos, G.J.; Holland, S.; Judelson, J.; Shoss, B.L.; Schmidt, E.J.; Md Noh, U.K.; D’Angelo, A.; et al. Corneal Toxicity Associated With Aquarium Coral Palytoxin. Am. J. Ophthalmol. 2017, 174, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Moshirfar, M.; Khalifa, Y.M.; Espandar, L.; Mifflin, M.D. Aquarium coral keratoconjunctivitis. Arch. Ophthalmol. 2010, 128, 1360–1362. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, Y.; Fuchs, J.; Beuschel, R.; Tschopp, M.; Goldblum, D. Dangerous reef aquaristics: Palytoxin of a brown encrusting anemone causes toxic corneal reactions. Toxicon 2015, 106, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Sosa, S.; Del Favero, G.; De Bortoli, M.; Vita, F.; Soranzo, M.R.; Beltramo, D.; Ardizzone, M.; Tubaro, A. Palytoxin toxicity after acute oral administration in mice. Toxicol. Lett. 2009, 191, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Ito, E.; Yasumoto, T. Toxicological studies on palytoxin and ostreocin-d administered to mice by three different routes. Toxicon 2009, 54, 244–251. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific Opinion on marine biotoxin in shellfish–Palytoxin group. EFSA J. 2009, 7, 1–38. [Google Scholar]
- Munday, R. Palytoxin toxicology: Animal studies. Toxicon 2011, 57, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Wiles, J.S.; Vick, J.A.; Christensen, M.K. Toxicological evaluation of palytoxin in several animal species. Toxicon 1974, 12, 427–433. [Google Scholar] [CrossRef]
- Ito, E.; Ohkusu, M.; Terao, K.; Yasumoto, T. Effects of repeated injections of palytoxin on lymphoid tissues in mice. Toxicon 1997, 35, 679–688. [Google Scholar] [CrossRef]
- Del Favero, G.; Beltramo, D.; Sciancalepore, M.; Lorenzon, P.; Coslovich, T.; Poli, M.; Testai, E.; Sosa, S.; Tubaro, A. Toxicity of palytoxin after repeated oral exposure in mice and in vitro effects on cardiomyocytes. Toxicon 2013, 75, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Suganuma, M.; Nakayasu, M.; Hakii, H.; Horiuchi, T.; Takayama, S.; Sugimura, T. Palytoxin is a non-12-O-tetradecanoylphorbol-13-acetate type tumor promoter in two-stage mouse skin carcinogenesis. Carcinogenesis 1986, 7, 707–710. [Google Scholar] [CrossRef] [PubMed]
- Wattenberg, E.V. Palytoxin: Exploiting a novel skin tumor promoter to explore signal transduction and carcinogenesis. Am. J. Physiol. Cell Physiol. 2007, 292, C24–C32. [Google Scholar] [CrossRef] [PubMed]
- Munday, R. Toxicological requirements for risk assessment of shellfish contaminants: A review. Afr. J. Mar. Sci. 2006, 28, 447–449. [Google Scholar] [CrossRef]
- Tubaro, A.; Del Favero, G.; Beltramo, D.; Ardizzone, M.; Forino, M.; De Bortoli, M.; Pelin, M.; Poli, M.; Bignami, G.; Ciminiello, P.; et al. Acute oral toxicity in mice of a new palytoxin analog: 42-hydroxy-palytoxin. Toxicon 2011, 57, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Usami, M.; Satake, M.; Ishida, S. Palytoxin analogs from the Dinoflagellate Ostreopsis siamensis. J. Am. Chem. Soc. 1995, 117, 5389–5390. [Google Scholar] [CrossRef]
- Pelin, M.; Forino, M.; Brovedani, V.; Tartaglione, L.; Dell’Aversano, C.; Pistocchi, R.; Poli, M.; Sosa, S.; Florio, C.; Ciminiello, P.; et al. Ovatoxin-a, A Palytoxin Analogue Isolated from Ostreopsis cf. ovata Fukuyo: Cytotoxic Activity and ELISA Detection. Environ. Sci. Technol. 2016, 50, 1544–1551. [Google Scholar] [CrossRef] [PubMed]
- Fussy, A.; Pommier, P.; Lumbroso, C.; de Haro, L. Chelonitoxism: New case reports in French Polynesia and review of the literature. Toxicon 2007, 49, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Pavlin, B.I.; Musto, J.; Pretrick, M.; Sarofalpiy, J.; Sappa, P.; Shapucy, S.; Kool, J. Mass poisoning after consumption of a hawksbill turtle, Federated States of Micronesia, 2010. West. Pac. Surveill. Resp. J. 2015, 6, 25–32. [Google Scholar] [CrossRef]
- Singh, S.S.; Biswas, A.K.; Shirley, P.; Vijayachari, P. Chelonitoxism in Andaman and Nicobar Island: A report on mass poisoning including a death of an adult. Toxicon 2016, 118, 61–63. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | TEF, 2009 EFSA | Oral LD50 Gavage (nmol/kg) | Oral LD50 Feeding (nmol/kg) | Relative Toxicity Gavage/Feeding | TEF, 2016 FAO/WHO |
|---|---|---|---|---|---|
| STX | 1 | 1190 | 3200 | 1 | 1 |
| NeoSTX | 1 | 700 | 1260 | 1.7/2.54 | 2 |
| GTX1 | 1 | 1 | |||
| GTX4 | 0.7 | 0.7 | |||
| GTX1&4 | 1610 | 3420 | 0.74/0.93 | ||
| GTX2 | 0.4 | 0.4 | |||
| GTX3 | 0.6 | 0.6 | |||
| GTX2&3 | 2230 | 5590 | 0.53/0.57 | ||
| GTX5 | 0.1 | 18,900 | 50,000 | 0.063/0.064 | 0.1 |
| GTX6 | 0.1 | 31,100 | >188,000 | 0.038/<0.017 | 0.05 |
| C1 | 0.01 | ||||
| C2 | 0.1 | 0.1 | |||
| C1&2 | 35,000 | 74,000 | 0.034/0.043 | ||
| C3 | 0.1 | 0.01 | |||
| C4 | 0.1 | 0.1 | |||
| C3&4 | 42,700 | 0.028/- | |||
| dc-STX | 1 | 2600 | 8680 | 0.46/0.37 | 0.5 |
| dc-neoSTX | 0.4 | 5500 | 14,300 | 0.22/0.22 | 0.2 |
| dc-GTX2 | 0.2 | 0.4 | |||
| dc-GTX3 | 0.4 | 0.4 | |||
| dc-GTX2&3 | 7130 | 29600 | 0.17/0.11 | ||
| References | [2] | [28,29] | [28,29] | [28,29] | [27] |
| Ciguatoxin | Mouse i.p. LD50 (µg/kg b.w.) | Reference | Proposed TEF (EFSA) |
|---|---|---|---|
| P-CTX-1 | 0.25 | [103,104] | 1 |
| P-CTX-2 | 0.9 | [82] | 0.3 |
| P-CTX-3 | 0.9 | [103] | 0.3 |
| P-CTX-3C | 2 | [82] | 0.2 |
| 2,3-dihydroxy P-CTX-3C | 1.8 | [105] | 0.1 |
| 51-hydroxy P-CTX-3C | 0.27 | [105] | 1 |
| P-CTX-4A | 2 | [106] | 0.1 |
| P-CTX-4B | 4 | [106,107] | 0.05 |
| C-CTX-1 | 3.6 | [97,108] | 0.1 |
| C-CTX-2 | 1 | [97] | 0.3 |
| Seafood Poisoning | Vectors | Onset Time and Duration | Major Acute Symptoms | Treatment |
|---|---|---|---|---|
| Paralytic shellfish poisoning (PSP) | Bivalve molluscs (clams, mussels, oysters, scallops), crab, lobster, gastropods, cephalopods, Atlantic salmon, herring, mackerel, puffer fish | Symptoms begin from 30 min to a few hours and can persist more than 24 h | Neurologic symptoms (tingling of lips and tongue, paresthesias, weakness, ataxia, dizziness, shortness of breath) | Supportive care: fluid therapy, respiratory support. |
| Amnesic shellfish poisoning (ASP) | Clams, mussels, oysters, scallops, squid, sardines, anchovies, crab and lobster | Symptoms begin from 15 min to 48 h and can persist several months in severe intoxications | Gastrointestinal (nausea, vomiting, diarrhea…) and neurologic symptoms (confusion, disorientation, memory loss, seizures) | Respiratory support and correction of cardiac dysrhythmias and hemodynamic instability |
| Ciguatera fish poisoning (CFP) | Fish, molluscs | Symptoms begin from 0.5 to 12 h. The acute phase lasts for 2–4 days. In the chronic phase symptoms persist weeks or months. | Gastrointestinal (nausea, vomiting, diarrhea…), neurologic symptoms (cold allodynia, itching, dizziness, ataxia, fatigue), cardiovascular (hypotension, bradycardia) | Intravenous mannitol and supportive care |
| Neurotoxin shellfish poisoning (NSP) | Mussels, clams, whelks, conch, coquinas, oysters, scallops, organs of some planktivorous fish | Symptoms begin 15 min to 3 h and last for a few days | Neurological (paresthesias, peripheral tingling, ataxia, myalgias, loss of coordination and coma in severe cases) and gastrointestinal symptoms (nausea, vomiting, abdominal pain and diarrhea). Respiratory problems and eye irritation can occur | General supportive care, bronchodilators, fluid replacement Gastrointestinal decontamination with activated charcoal for patients with recent ingestion of toxin. Administration of sedatives and pain mitigation |
| TTX poisoning | Pufferfish, goby fish, gastropods, crabs and bivalves | Symptoms occur within 10–45 min, sometimes delayed to 3–6 h but human data on recovery are very variable | Numbness, paraesthesia, incoordination, severe respiratory failure, hypotension, cardiac dysrhythmias and death in fatal cases | There is no specific antidote. Treatment is supportive, with removal of unabsorbed toxin. Treatment options included cysteine, cholinesterase inhibitors, naloxone and steroids |
| Diarrheic shellfish poisoning (DSP) | Mussels, oysters, scallops, clams, cockles | Symptoms occur from 30 min to 5 h and continue for about 2–3 days | Diarrhea, gastrointestinal distress, nausea, vomiting, and abdominal pain | Replacement of electrolyte and fluid loss |
| Azaspiracid poisoning (AZP) | Mussels, oysters, scallops, clams, cockles | Onset: 3 h Duration: 15 h | Diarrhea, nausea, vomiting, and abdominal cramps | Replacement of electrolyte and fluid loss |
| Palytoxin poisoning | Fish, sea urchin, molluscs, crabs, octopus | Onset: Minutes to several hours | Bitter taste, oral and limb numbness, dizziness, myalgia, rabdomyolysis | Supportive care |
| Seafood Poisoning | Representative Toxin/s | Regulated Levels in Seafood |
|---|---|---|
| Paralytic shellfish poisoning (PSP) | Saxitoxin (STX (2HCl)) | 0.8 mg STX eq/kg |
| Amnesic shellfish poisoning (ASP) | Domoic acid (DA) and isoDA | 20 mg DA eq/kg |
| Ciguatera fish poisoning (CFP) | Pacific-ciguatoxin-1 (P-CTX-1) Caribbean-ciguatoxin-1 (C-CTX-1) | Absence (EU) 10 ng P-CTX-1 eq/kg (USA) 100 ng C-CTX-1/kg (USA) 0.025 MU/g (Japan, Mexico) |
| Neurotoxin shellfish poisoning (NSP) | _ | 20 MU/100 g |
| TTX poisoning | _ | Absence (EU) |
| Diarrheic shellfish poisoning (DSP) | Okadaic acid (OA) | 0.16 mg OA eq/kg |
| Azaspiracid poisoning (AZP) | Azaspiracid | 0.16 mg AZA eq/kg |
| Palytoxin poisoning | _ | _ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilariño, N.; Louzao, M.C.; Abal, P.; Cagide, E.; Carrera, C.; Vieytes, M.R.; Botana, L.M. Human Poisoning from Marine Toxins: Unknowns for Optimal Consumer Protection. Toxins 2018, 10, 324. https://doi.org/10.3390/toxins10080324
Vilariño N, Louzao MC, Abal P, Cagide E, Carrera C, Vieytes MR, Botana LM. Human Poisoning from Marine Toxins: Unknowns for Optimal Consumer Protection. Toxins. 2018; 10(8):324. https://doi.org/10.3390/toxins10080324
Chicago/Turabian StyleVilariño, Natalia, M. Carmen Louzao, Paula Abal, Eva Cagide, Cristina Carrera, Mercedes R. Vieytes, and Luis M. Botana. 2018. "Human Poisoning from Marine Toxins: Unknowns for Optimal Consumer Protection" Toxins 10, no. 8: 324. https://doi.org/10.3390/toxins10080324
APA StyleVilariño, N., Louzao, M. C., Abal, P., Cagide, E., Carrera, C., Vieytes, M. R., & Botana, L. M. (2018). Human Poisoning from Marine Toxins: Unknowns for Optimal Consumer Protection. Toxins, 10(8), 324. https://doi.org/10.3390/toxins10080324