Characterisation of the Mycobiota on the Shell Surface of Table Eggs Acquired from Different Egg-Laying Hen Breeding Systems


 ,
,  ,
,
Abstract
1. Introduction
2. Results and Discussion
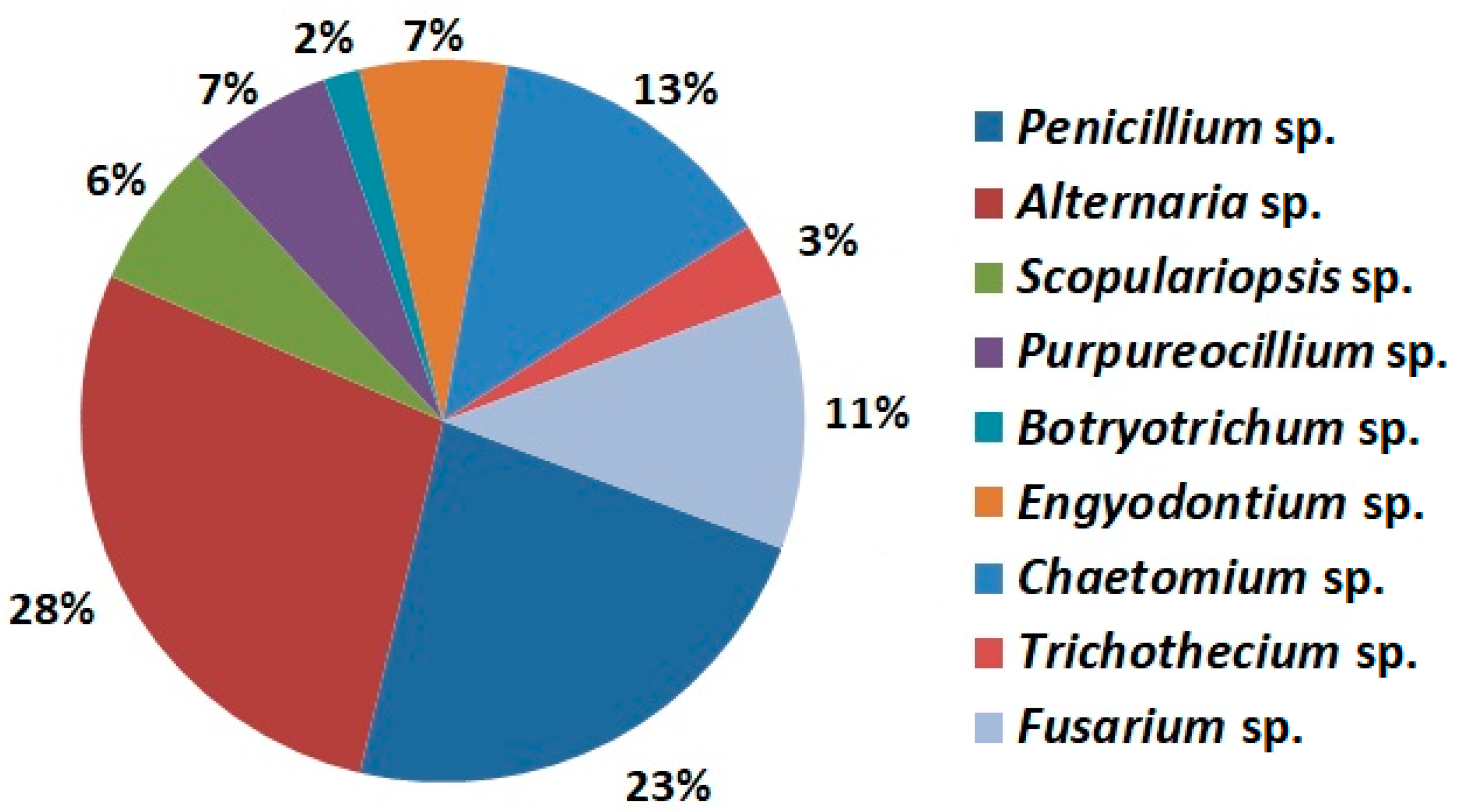
2.1. Mycobiota Species Identification
2.2. Mycobiota Strains Isolated from Eggs Acquired from Different Egg-Laying Hen Breeding System
2.3. Toxin Content Analysis
2.4. Toxin Content Analysis in Eggs Acquired from Different Egg-Laying Hen Breeding System
3. Conclusions
4. Materials and Methods
4.1. Egg Collection
4.2. Mycobiota Isolation, Media and Culture Conditions
4.3. DNA Extraction
4.4. Molecular Species Identification
4.5. Analysis of Mycotoxins in Egg White
Author Contributions
Funding
Conflicts of Interest
References
- Commission Regulation (EC) 1881/2006. Off. J. Eur. Commun. 2006, 558, L364.
- De Reu, K.; Grijspeerdt, W.; Messensa, M.; Heyndrickxa, M.; Uyttendaeleb, J.; Debevereb, L.; Hermana, L. Eggshell factors influencing eggshell penetration and whole egg contamination by different bacteria, including Salmonella enteritidis. Int. J. Food Microbiol. 2006, 112, 2532–2560. [Google Scholar] [CrossRef] [PubMed]
- Gantois, I.; Ducatelle, R.; Pasmans, F.; Haesebrouck, F.; Gast, R.; Humphrey, T.J.; Van Immerseel, F. Mechanisms of egg contamination by Salmonella enteritidis. FEMS Microbiol. Lett. 2009, 33, 718–738. [Google Scholar] [CrossRef] [PubMed]
- Corry, J.E.L. Microbiological analysis of eggs and egg products. In Microbiological Analysis of Red Meat, Poultry and Eggs; Mead Red, G.C., Ed.; CRC Press: Cambridge, UK, 2007; pp. 183–201. [Google Scholar]
- Gros, R.V.; Nichita, I.; Șereș, M.; Ilie, M.S.; Marcu, A.; Cucerzan, A.; Tîrziu, E. Study of the fungi dynamics in a poultry house with permanent litter. Lucr. St. Med. Vet. 2015, 48, 2572–2662. [Google Scholar]
- Harry, E.G.; Cooper, D.M. The treatment of hatching eggs for the control of egg transmitted aspergillosis. Br. Poult. Sci. 1970, 11, 2692–2772. [Google Scholar] [CrossRef] [PubMed]
- Girma, G.; Abebaw, M.; Zemene, M.; Mamuye, Y.; Getaneh, G. A Review on Aspergillosis in Poultry. J. Vet. Sci. Technol. 2016, 7, 6. [Google Scholar] [CrossRef]
- Szablewski, T.; Stuper, K.; Cegielska-Radziejewska, R.; Kijowski, J.; Perkowski, J. Ergosterol as an indicator of the presence of microscopic fungi in eggs for human consumption produced in different husbandry systems. Poult. Sci. 2010, 89, 2491–2493. [Google Scholar] [CrossRef] [PubMed]
- Rohweder, D.; Valenta, H.; Sondermann, S.; Schollenberger, M.; Drochner, W.; Pahlow, G.; Döll, S.; Dänicke, S. Effect of different storage conditions on the mycotoxin contamination of Fusarium culmorum-infected and non-infected wheat straw. Mycotoxin Res. 2011, 27, 1451–1453. [Google Scholar]
- Kokkonen, M.; Ojala, L.; Parikka, P.; Jestoi, M. Mycotoxin production of selected Fusarium species at different culture conditions. Int. J. Food Microbiol. 2010, 30, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Stuper-Szablewska, K.; Szablewski, T.; Cegielska-Radziejewska, R.; Ostrowska, A.; Matysiak, A.; Perkowski, J. Contamination of different kind of litter by microscopic fungi from hen house. Apar. Bad. Dydakt. 2014, 2, 1992–2004. [Google Scholar]
- Skóra, J.; Gutarowska, B.; Pielech-Przybylska, K.; Stępień, Ł.; Pietrzak, K.; Piotrowska, M.; Pietrowski, P. Assessment of microbiological contamination in the work environments of museums, archives and libraries. Aerobiologia 2015, 31, 3894–3901. [Google Scholar] [CrossRef] [PubMed]
- Sypecka, Z.; Kelly, M.; Brereton, P. Deoxynivalenol and Zearalenone Residues in Eggs of Laying Hens Fed with a Naturally Contaminated Diet, Effects on Egg Production and Estimation of Transmission Rates from Feed to Eggs. J. Agric. Food Chem. 2004, 25, 5463–5471. [Google Scholar] [CrossRef] [PubMed]
- Binder, E.M.; Tan, L.M.; Chin, L.J.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Desjardins, A.E. Fusarium Mycotoxins Chemistry, Genetics and Biology; ASP Press: Eagan, MN, USA, 2006. [Google Scholar]
- Zur, G.; Shimoni, E.; Hallerman, E.; Kashi, Y. Detection of Alternaria fungal contamination in cereal grains by a polymerase chain reaction-based assay. J. Food Prot. 2002, 65, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.F.; Gravesen, S.; Nielsen, P.A.; Andersen, B.; Thrane, U.; Frisvad, J.C. Production of mycotoxins on artificially and naturally infested building materials. Mycopathologia 1999, 145, 435–436. [Google Scholar] [CrossRef]
- Fogle, M.R.; Douglas, D.R.; Jumper, C.A.; Straus, D.C. Growth and mycotoxin production by Chaetomium globosum. Mycopathologia 2007, 164, 495–496. [Google Scholar] [CrossRef] [PubMed]
- Provost, N.; Shi, C.; She, Y.; Cyr, T.; Miller, D. Characterization of an antigenic chitosanase from the cellulolytic fungus Chaetomium globosum. Med. Mycol. 2013, 51, 2902–2999. [Google Scholar] [CrossRef] [PubMed]
- Piskorska-Pliszczynska, J.; Mikolajczyk, M.; Warenik-Bany, M.; Maszewski, S.; Strucinski, P. Soil as a source of dioxin contamination in eggs from free-range hens on a Polish farm. Sci. Total Environ. 2014, 145, 435–436. [Google Scholar] [CrossRef] [PubMed]
- Macêdo, D.P.C.; Neves, R.P.; De Souza-Motta, C.M.; Magalhães, O.M.C. Engyodontium album fungaemia, the first reported case. Braz. J. Microbiol. 2007, 38, 1101–1112. [Google Scholar] [CrossRef]
- Holt, P.S.; Davies, R.H.; Dewulf, J.; Gast, R.K.; Huwe, J.K.; Jones, D.R.; Waltman, D.; Willian, K.R. The impact of different housing systems on egg safety and quality. Poult. Sci. 2011, 90, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Wall, H.; Tauson, R.; Sorgjerd, S. Bacterial contamination of eggshells in furnished and conventional cages. J. Appl. Poult. Res. 2008, 17, 11–16. [Google Scholar] [CrossRef]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum, causal agent of foot and root rot and head blight on wheat. Mol. Plant Pathol. 2013, 14, 3233–3241. [Google Scholar] [CrossRef] [PubMed]
- Kuzdraliński, A.; Nowak, M.; Szczerba, H.; Dudziak, K.; Muszyńska, M.; Leśniowska-Nowak, J. The composition of Fusarium species in wheat husks and grains in south-eastern Poland. J. Integr. Agric. 2017, 16, 1530–1536. [Google Scholar] [CrossRef]
- Stępień, Ł.; Waśkiewicz, A.; Urbaniak, M. Wildly growing asparagus (Asparagus officinalis L.) hosts pathogenic Fusarium species and accumulates their mycotoxins. Microb. Ecol. 2016, 71, 9279–9337. [Google Scholar] [CrossRef] [PubMed]
- Ciegler, A. Fungi that produce mycotoxins, Conditions and occurrence. Mycopathologia 1978, 65, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Stępień, Ł.; Gromadzka, K.; Chełkowski, J. Polymorphism of mycotoxin biosynthetic genes among Fusarium equiseti isolates from Italy and Poland. J. Appl. Genet. 2012, 53, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Prelusky, D.B.; Hamilton, R.M.G.; Trenholm, H.L. Transmission of Residues to Eggs Following Long-Term Administration of 14C-Labelled Deoxynivalenol to Laying Hens. Poult. Sci. 1989, 68, 744–748. [Google Scholar] [CrossRef] [PubMed]
- Prelusky, D.B.; Trenholm, H.L.; Hamilton, R.M.G.; Miller, J.D. Transmission of [14C] Deoxynivalenol to Eggs following Oral Administration to Laying Hens. J. Agric. Food Chem. 1987, 35, 1821–1886. [Google Scholar] [CrossRef]
- Valenta, H.; Dänicke, S. Carry-over of deoxynivalenol into eggs of laying hens—Preliminary results. Mycotoxin Res. 2005, 21, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Tangni, E.K.; Waegeneersa, N.; Van Overmeireb, I.; Goeyensb, L.; Pussemier, L. Mycotoxin analyses in some home produced eggs in Belgium reveal small contribution to the total daily intake. Sci. Total Environ. 2009, 407, 4411–4418. [Google Scholar] [CrossRef] [PubMed]
- Escrivá, L.; Font, G.; Manyes, L.; Berrada, H. Studies on the Presence of Mycotoxins in Biological Samples: An Overview. Toxins 2017, 9, 251. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006; pp. 1–388. [Google Scholar]
- Stępień, Ł.; Jestoi, M.; Chełkowski, J. Cyclic hexadepsipeptides in wheat field samples and esyn1 gene divergence among enniatin producing Fusarium avenaceum strains. World Mycotoxin J. 2013, 6, 399–409. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Kullnig-Gradinger, C.M.; Szakacs, G.; Kubicek, C.P. Phylogeny and evolution of genus Trichoderma a multigene approach. Mycol. Res. 2002, 106, 757–767. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetic. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Shinsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Stępień, Ł.; Waśkiewicz, A. Sequence divergence of the enniatin synthase gene in relation to production of beauvericin and enniatins in Fusarium species. Toxins 2013, 5, 537–555. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Hen Housing System | Sample Treatment | Identified Fungal Isolates |
|---|---|---|
| Cage | Native mycobiota | Alternaria brassicae Alternaria obovoidea Botryotrichum spirotrichum Chaetomium murorum |
| Surface-sterilised | Alternaria arborescens Alternaria burnsii Alternaria tenuissima Engyodontium album | |
| Deep litter housing | Native mycobiota | Alternaria alternata Alternaria tenuissima Chaetomium globosum Engyodontium album Fusarium culmorum Fusarium equiseti |
| Surface-sterilised | Acrostalagmus luteoalbus Alternaria arborescens Alternaria brassicae Alternaria infectoria Alternaria multiformis Alternaria tenuissima Chaetomium globosum Penicillium chrysogenum Penicillium riseofulvum Scopulariopsis brevicaulis Trichothecium roseum | |
| Free range | Native mycobiota | Alternaria infectoria Alternaria tenuissima Chaetomium funicola Chaetomium globosum Epicoccum nigrum Fusarium culmorum Fusarium tricinctum Trichothecium roseum |
| Surface-sterilised | Alternaria sp. Acrostalagmus luteoalbus Chaetomium globosum Purpureocillium lilacinum | |
| Organic | Native mycobiota | Alternaria alternata Epicoccum nigrum Trichothecium roseum |
| Surface-sterilised | Alternaria infectoria Alternaria multiformis Alternaria brassicae Alternaria tenuissima Chaetomium elatum Chaetomium globosum Epicoccum nigrum Purpureocillium lilacinum Trichothecium roseum |
| Fusarium sp. | Concentration of Type-A Trichothecenes (μg/kg) | |||||
|---|---|---|---|---|---|---|
| Scirpentriol | T-2 Tetraol | T-2 Triol | DAS | HT-2 | T-2 | |
| Fc | 3 ± 1 | 5 ± 1 | 2 ± 1 | 11 ± 9 | 16 ± 8 | 22 ± 7 |
| Fe | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| Ft | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| none | 10 ± 9 | <LOD | <LOD | <LOD | <LOD | 13 ± 9 |
| Fusarium sp. | Concentration of Type-B Trichothecenes (μg/kg) ± SD | ||||
|---|---|---|---|---|---|
| DON | FUS-X | 3-AcDON | 15-AcDON | NIV | |
| Fc | 16 ± 3 | 2 ± 1 | 4 ± 2 | 3 ± 1 | 55 ± 11 |
| Fe | <LOD | <LOD | <LOD | <LOD | <LOD |
| Ft | <LOD | <LOD | <LOD | <LOD | <LOD |
| none | 10 ± 4 | <LOD | <LOD | <LOD | 19 ± 9 |
| Egg Content | Total Toxin Concentrations (μg/kg) | ||
|---|---|---|---|
| Range | Mean | ||
| Native mycobiota | Organic | <LOD | - |
| Free range | 0.1–55 | 36 a | |
| Deep litter | 0.1–38 | 27 a | |
| Cage | <LOD | - | |
| Surface-sterilised | Organic | <LOD | - |
| Free range | <LOD | - | |
| Deep litter | 13–19 | 14 | |
| Cage | <LOD | - | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomczyk, Ł.; Stępień, Ł.; Urbaniak, M.; Szablewski, T.; Cegielska-Radziejewska, R.; Stuper-Szablewska, K. Characterisation of the Mycobiota on the Shell Surface of Table Eggs Acquired from Different Egg-Laying Hen Breeding Systems. Toxins 2018, 10, 293. https://doi.org/10.3390/toxins10070293
Tomczyk Ł, Stępień Ł, Urbaniak M, Szablewski T, Cegielska-Radziejewska R, Stuper-Szablewska K. Characterisation of the Mycobiota on the Shell Surface of Table Eggs Acquired from Different Egg-Laying Hen Breeding Systems. Toxins. 2018; 10(7):293. https://doi.org/10.3390/toxins10070293
Chicago/Turabian StyleTomczyk, Łukasz, Łukasz Stępień, Monika Urbaniak, Tomasz Szablewski, Renata Cegielska-Radziejewska, and Kinga Stuper-Szablewska. 2018. "Characterisation of the Mycobiota on the Shell Surface of Table Eggs Acquired from Different Egg-Laying Hen Breeding Systems" Toxins 10, no. 7: 293. https://doi.org/10.3390/toxins10070293
APA StyleTomczyk, Ł., Stępień, Ł., Urbaniak, M., Szablewski, T., Cegielska-Radziejewska, R., & Stuper-Szablewska, K. (2018). Characterisation of the Mycobiota on the Shell Surface of Table Eggs Acquired from Different Egg-Laying Hen Breeding Systems. Toxins, 10(7), 293. https://doi.org/10.3390/toxins10070293