Development of Time-Resolved Fluoroimmunoassay for Detection of Cylindrospermopsin Using Its Novel Monoclonal Antibodies


Abstract

1. Introduction
2. Results and Discussion
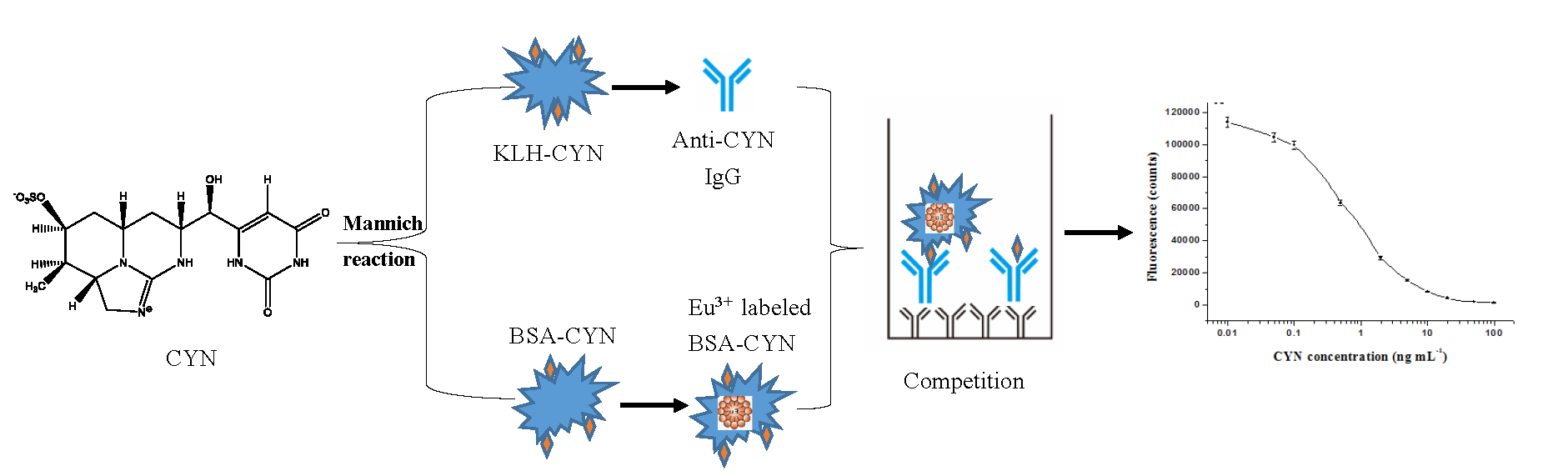
2.1. CYN Conjugate Preparation
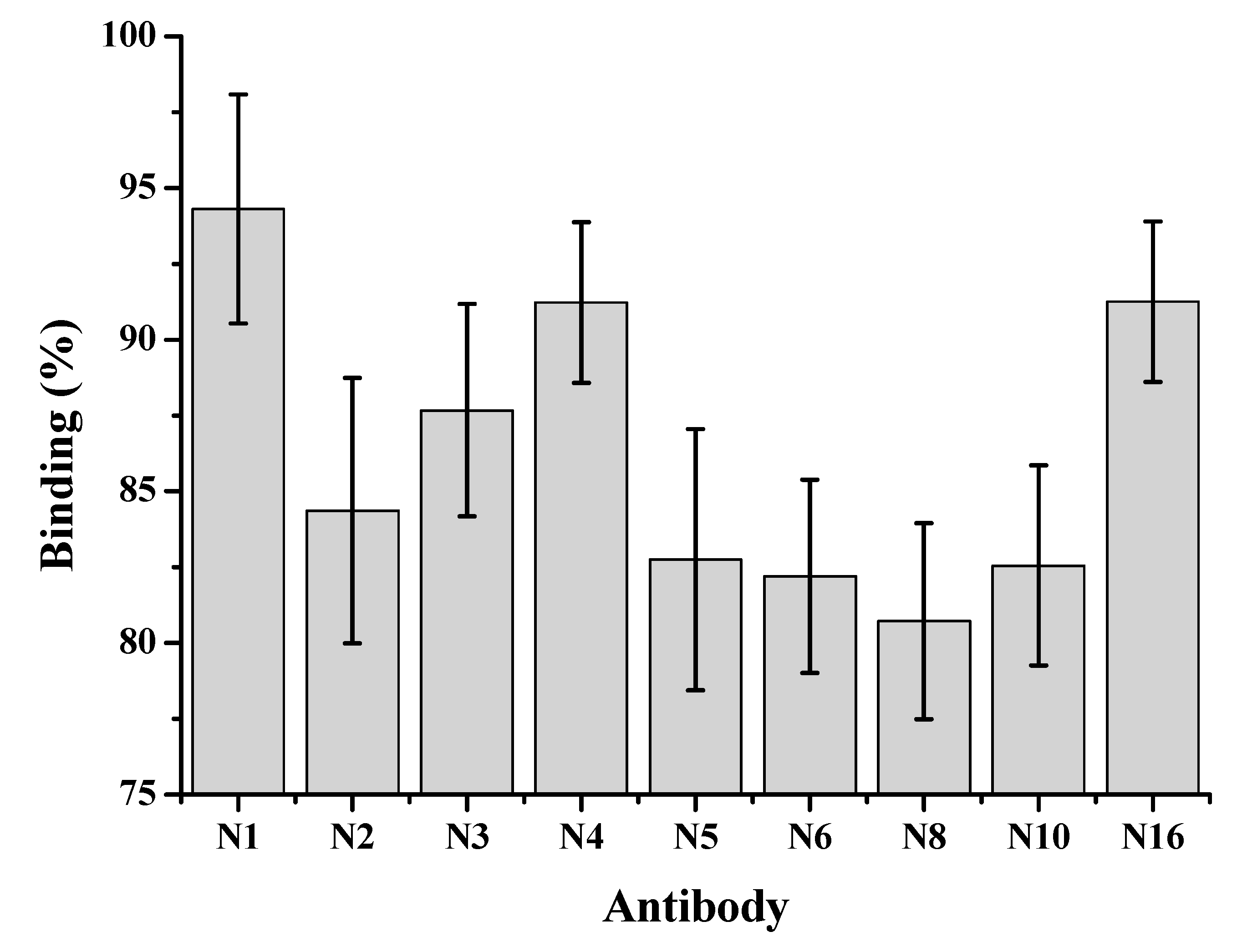
2.2. Monoclonal Antibody Production
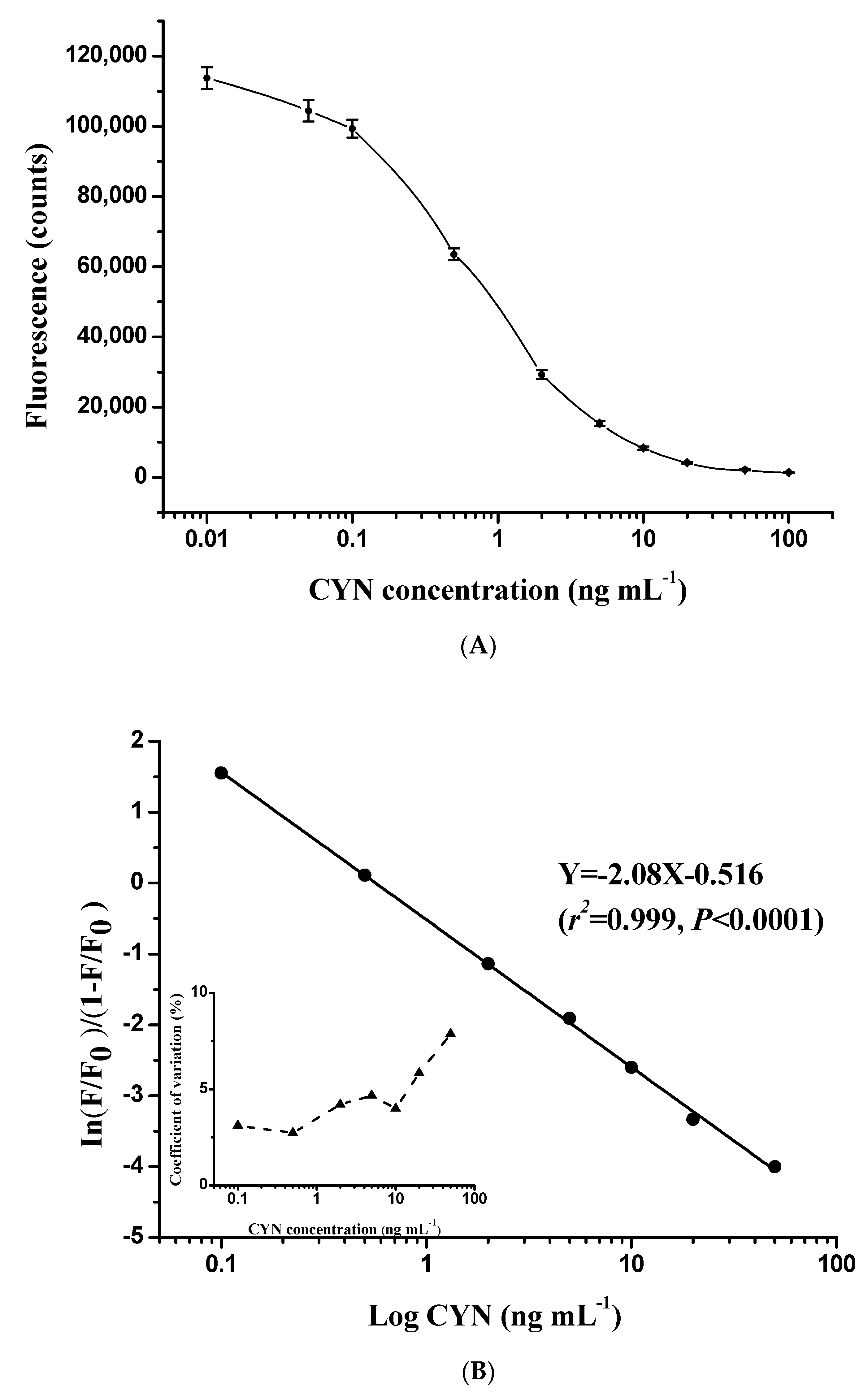
2.3. Establishment of Standard Curves
2.4. Assay Validation
2.4.1. Analytical Sensitivity of the Present Method
2.4.2. Precision and Accuracy of the Present Method
2.4.3. Recovery of the Developed Method
2.4.4. Dilution Linearity for the Present Method
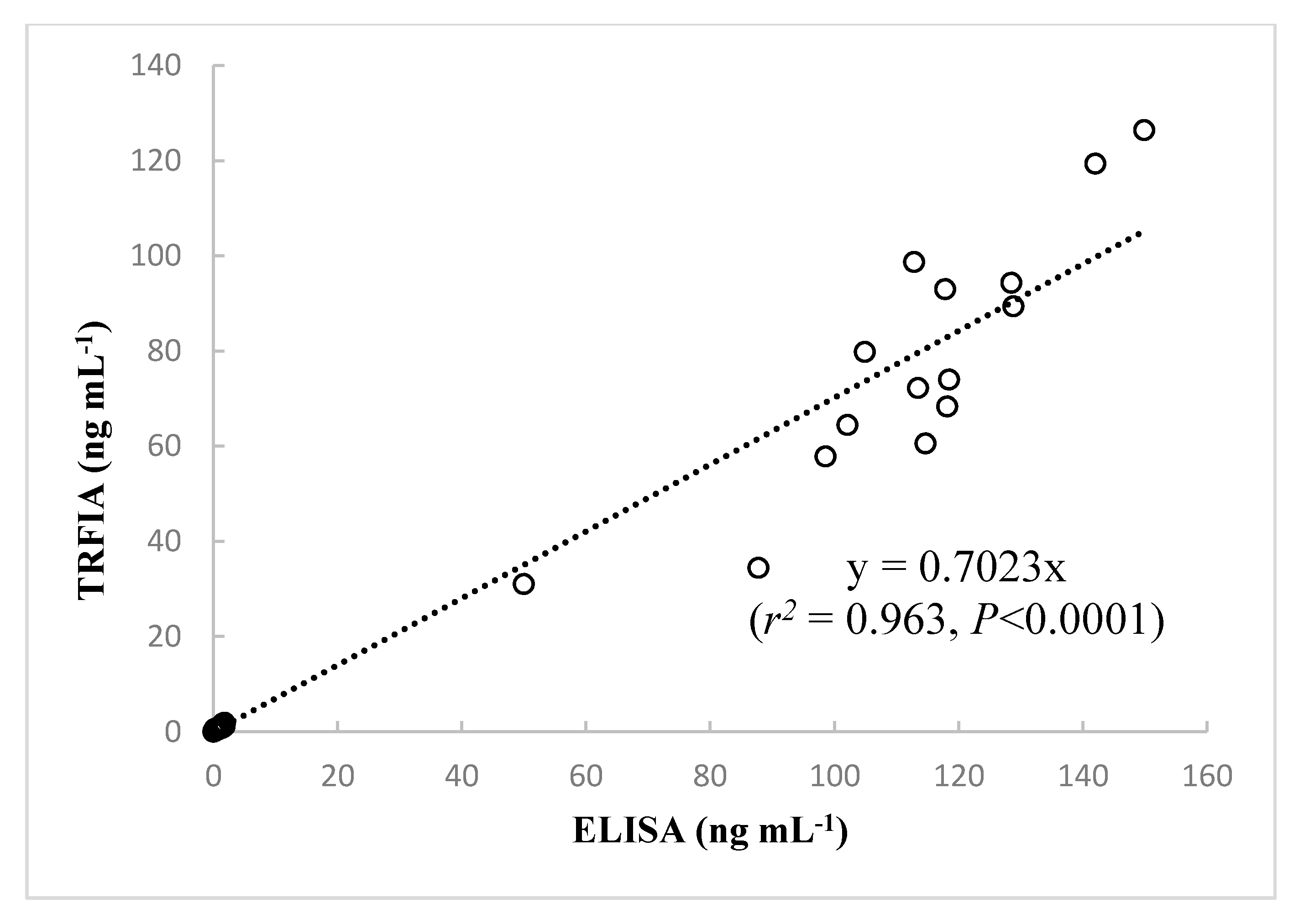
2.5. Comparison of Assay Results and Performance of the Developed TRFIA and ELISA
3. Materials and Method
3.1. Chemicals and Solutions
3.2. Preparation of Protein Conjugates KLH-CYN and BSA-CYN
3.3. Monoclonal Antibody Production
3.4. Eu3+-Labeled BSA-CYN
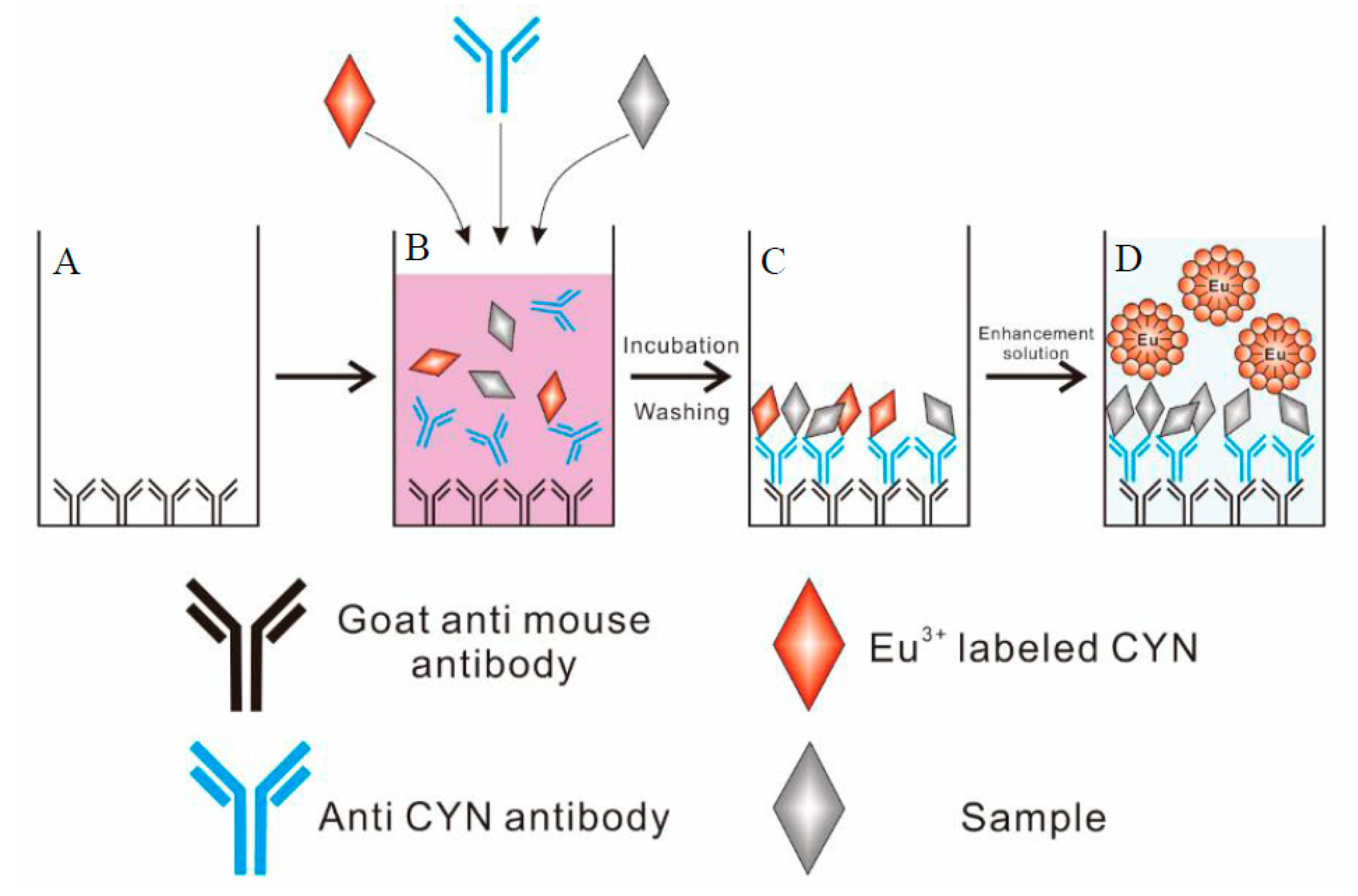
3.5. Coating with Secondary Antibody
3.6. Development of the Direct Competitive TRFIA
3.7. Validation
3.7.1. Analytical Sensitivity
3.7.2. Precision and Accuracy of the Assay
3.7.3. Spiked Sample Analysis
3.7.4. Dilution Linearity Test
3.8. Comparison of Sample Analysis with ELISA Kit
3.9. Data Analysis
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Moreira, C.; Azevedo, J.; Antunes, A.; Vasconcelos, V. Cylindrospermopsin: Occurrence, methods of detection and toxicology. J. Appl. Microbiol. 2013, 114, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Poniedziałek, B. In search of environmental role of cylindrospermopsin: A review on global distribution and ecology of its producers. Water Res. 2014, 66, 320–337. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, I.; Moore, R.E.; Runnegar, M.T.C. Cylindrospermopsin, a potent hepatotoxin from the blue-green alga Cylindrospermopsis raciborskii. J. Am. Chem. Soc. 1992, 114, 7942–7944. [Google Scholar] [CrossRef]
- Banker, R.; Carmeli, S.; Werman, M.; Teltsch, B.; Porat, R.; Sukenik, A. Uracil moiety is required for toxicity of the cyanobacterial hepatotoxin cylindrospermopsin. J. Toxicol. Environ. Health Part A 2001, 62, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Carmichael, W.W.; Brittain, S.; Eaglesham, G.K.; Shaw, G.R.; Liu, Y.D.; Watanabe, M.M. First report of the cyanotoxins cylindrospermopsin and deoxycylindrospermopsin from Rhaphidiopsis curvata (Cyanobacteria). J. Phycol. 2001, 37, 1121–1126. [Google Scholar] [CrossRef]
- Poniedziałek, B.; Rzymski, P.; Kokociński, M. Cylindrospermopsin: Water-linked potential threat to human health in Europe. Environm. Toxicol. Pharmacol. 2012, 34, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Pichardo, S.; Cameán, A.M.; Jos, A. In Vitro Toxicological Assessment of Cylindrospermopsin: A Review. Toxins (Basel) 2017, 9, E402. [Google Scholar] [CrossRef] [PubMed]
- Poniedzialek, B.; Rzymski, P.; Karczewski, J. The role of the enzymatic antioxidant system in cylindrospermopsin-induced toxicity in human lymphocytes. Toxicol. In Vitro 2015, 29, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Poniedzialek, B.; Rzymski, P.; Wiktotowicz, K. First report of cylindrospermopsin effect on human peripheral blood lymphocytes proliferation in vitro. Cent. Eur. J. Immunol. 2012, 37, 314–317. [Google Scholar] [CrossRef]
- Poniedzialek, B.; Rzymski, P.; Wiktorowicz, K. Toxocity of cylindrospermopsin in human lymphocytes: Proliferation, viability and cell cycle studies. Toxicol. In Vitro 2014, 28, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Kinnear, S. Cylindrospermopsin: A Decade of Progress on Bioaccumulation Research. Mar. Drugs 2010, 8, 542–564. [Google Scholar] [CrossRef] [PubMed]
- Adamski, M.; Żmudzki, P.; Chrapusta, E.; Bober, B.; Kaminski, A.; Zabaglo, K.; Latkowska, E.; Bialczyk, J. Effect of pH and temperature on the stability of cylindrospermopsin. Characterization of decomposition products. Algal Res. 2016, 15, 129–134. [Google Scholar] [CrossRef]
- Moreira, C.; Ramos, V.; Azevedo, J.; Vasconcelos, V. Methods to detect cyanobacteria and their toxins in the environment. Appl. Microbiol. Biotechnol. 2014, 98, 8073–8082. [Google Scholar] [CrossRef] [PubMed]
- Eaglesham, G.K.; Norris, R.L.; Shaw, G.R.; Smith, M.J.; Chiswell, R.K.; Davis, B.C.; Neville, G.R.; Seawright, A.A.; Moore, M.R. Use of HPLC-MS/MS to monitor cylindrospermopsin, a blue–green algal toxin, for public health purposes. Environ. Toxicol. 1999, 14, 151–154. [Google Scholar] [CrossRef]
- Welker, M.; Bickel, H.; Fastner, J. HPLC-PDA detection of cylindrospermopsin—Opportunities and limits. Water Res. 2002, 36, 4659–4663. [Google Scholar] [CrossRef]
- Rasmussen, J.P.; Giglio, S.; Monis, P.T.; Campbell, R.J.; Saint, C.P. Development and field testing of a real-time PCR assay for cylindrospermopsin-producing cyanobacteria. J. Appl. Microbiol. 2008, 104, 1503–1515. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Philips, E.D.; Szabo, N.J.; Badylak, S. A comparative study of Florida strains of Cylindrospermopsis and Aphanizomenon for cylindrospermopsin production. Toxicon 2008, 51, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Bláhová, L.; Oravec, M.; Maršálek, B.; Šejnohová, L.; Šimek, Z.; Bláha, L. The first occurrence of the cyanobacterial alkaloid toxin cylindrospermopsin in the Czech Republic as determined by immunochemical and LC/MS methods. Toxicon 2009, 53, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.W.; Orr, P.T.; Boyer, G.L.; Burford, M.A. Investigating the production and release of cylindrospermopsin and deoxy-cylindrospermopsin by Cylindrospermopsis raciborskii over a natural growth cycle. Harmful Algae 2014, 31, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.P.; Lind, O. First evidence of “paralytic shellfish toxins” and cylindrospermopsin in a Mexican freshwater system, Lago Catemaco., and apparent bioaccumulation of the toxins in “tegogolo” snails (Pomacea patula catemacensis). Toxicon 2010, 55, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Al-Tebrineh, J.; Merrick, C.; Ryan, D.; Humpage, A.; Bowling, L.; Neilan, B.A. Community composition, toxigenicity, and environmental conditions during a cyanobacterial bloom occurring along 1,100 kilometers of the Murray River. Appl. Environ. Microbiol. 2012, 78, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Z.A.; Al-Shehri, A.M. Assessment of cylindrospermopsin toxin in an arid Saudi lake containing dense cyanobacterial bloom. Environ. Monit. Assess. 2013, 185, 2157–2166. [Google Scholar] [CrossRef] [PubMed]
- Somdee, T.; Kaewsan, T.; Somdee, A. Monitoring toxic cyanobacteria and cyanotoxins (microcystins and cylindrospermopsins) in four recreational reservoirs (Khon Kaen, Thailand). Environ. Monit. Assess. 2013, 185, 9521–9529. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Peng, L.; Huang, X.; Han, B.-P. Occurrence and dominance of Cylindrospermopsis raciborskii and dissolved cylindrospermopsin in urban reservoirs used for drinking water supply, South China. Environ. Monit. Assess. 2014, 186, 3079–3090. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Young, F.M.; Codd, G.A. Performance assessment of a cylindrospermopsin ELISA with purified compounds and cyanobacterial extracts. Environ. Forensics 2017, 18, 147–152. [Google Scholar] [CrossRef]
- Elliott, C.T.; Redshaw, C.H.; George, S.E.; Campbell, K. First development and characterisation of polyclonal and monoclonal antibodies to the emerging fresh water toxin cylindrospermopsin. Harmful Algae 2013, 24, 10–19. [Google Scholar] [CrossRef]
- Fraga, M.; Vilariño, N.; Louzao, M.C.; Rodríguez, L.P.; Alfonso, A.; Campbell, K.; Elliott, C.T.; Taylor, P.; Ramos, V.; Vasconcelos, V.; et al. Multi-detection method for five common microalgal toxins based on the use of microspheres coupled to a flow-cytometry system. Anal. Chim. Acta 2014, 850, 57–64. [Google Scholar] [CrossRef] [PubMed]
- McNamee, S.E.; Elliott, C.T.; Greer, B.; Lochhead, M.; Campbell, K. Development of a planar waveguide microarray for the monitoring and early detection of five harmful algal toxins in water and cultures. Environ. Sci. Technol. 2014, 48, 13340–13349. [Google Scholar] [CrossRef] [PubMed]
- Dickson, E.F.; Pollak, A.; Diamandis, E.P. Time-resolved detection of lanthanide luminescence for ultrasensitive bioanalytical assays. J. Photochem. Photobiol. B 1995, 27, 3–19. [Google Scholar] [CrossRef]
- Wang, K.; Huang, B.; Zhang, J.; Zhou, B.; Gao, L.; Zhu, L.; Jin, J. A novel and sensitive method for the detection of deoxynivalenol in food by time-resolved fluoroimmunoassay. Toxicol. Mech. Methods 2009, 19, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Wang, Y.; Song, B.; Wang, X.; Ma, H.; Yuan, J. Homogeneous time-resolved fluoroimmunoassay of microcystin-LR using layered WS2 nanosheets as a transducer. Methods Appl. Fluoresc. 2017, 5, 024007. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, P.R.; Chandrasena, N.R.; Jones, G.J.; Humpage, A.R.; Falconer, I.R. Isolation and toxicity of Cylindrospermopsis raciborskii from an ornamental lake. Toxicon 1997, 35, 341–346. [Google Scholar] [CrossRef]
- Humpage, A.R.; Falconer, I.R. Oral toxicity of the cyanobacterial toxin cylindrospermopsin in male Swiss albino mice, determination of no observed adverse effect level for deriving a drinking water guideline value. Environ. Toxicol. 2003, 18, 94–103. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz, A.A.; Hiskia, A.; Kaloudis, T.; Chernoff, N.; Hill, D.; Antoniou, M.G.; He, X.; Loftin, K.; O’Shea, K.; Zhao, C.; et al. A review on cylindrospermopsin: The global occurrence, detection, toxicity and degradation of a potent cyanotoxin. Environ. Sci. Process Impacts 2013, 15, 1979–2003. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Dai, R.; Zhang, H.; Li, C.; Zhang, X.; Shen, J.; Wen, K.; Wang, Z. Production of monoclonal antibodies with broad specificity and development of an immunoassay for microcystins and nodularin in water. Anal. Bioanal. Chem. 2016, 408, 6037–6044. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.; Stewart, L.D.; Doucette, G.J.; Fodey, T.L.; Haughey, S.A.; Vilariño, N.; Kawatsu, K.; Elliott, C.T. Assessment of specific binding proteins suitable for the detection of paralytic shellfish poisons using optical biosensor technology. Anal. Chem. 2007, 79, 5906–5914. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S.; Soutome, H.; Tsutsumi, T.; Hasegawa, A.; Sekijima, M.; Sugamata, M.; Harada, K.; Suganuma, M.; Ueno, Y. Novel monoclonal antibodies against microcystin and their protective activity for hepatotoxicity. Nat. Toxins 1995, 3, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Young, F.M.; Metcalf, J.S.; Meriluoto, J.A.; Spoof, L.; Morrison, L.F.; Codd, G.A. Production of antibodies against microcystin-RR for the assessment of purified microcystins and cyanobacterial environmental samples. Toxicon 2006, 48, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.; Stack, E.; Krivelo, S.; McPartlin, D.A.; Byrne, B.; Greef, C.; Lochhead, M.J.; Husar, G.; Devlin, S.; Elliott, C.T.; et al. Detection of the cyanobacterial toxin, microcystin-LR, using a novel recombinant antibody-based optical-planar waveguide platform. Bios. Bioelectron. 2015, 67, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Zeck, A.; Eikenberg, A.; Weller, M.G.; Niessner, R. Highly sensitive immunoassay based on amonoclonal antibody specific for [4-arginine] microcystins. Anal. Chim. Acta 2001, 441, 1–13. [Google Scholar] [CrossRef]
- Sheng, J.W.; He, M.; Shi, H.C. A highly specific immunoassay for microcystin-LR detection based on a monoclonal antibody. Anal. Chim. Acta 2007, 603, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.G. Immunoassays and biosensors for the detection of cyanobacterial toxins in water. Sensors (Basel) 2013, 13, 15085–15112. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.M.; Wu, Y.S.; Gan, N.Q.; Song, L.R. An ELISA-like time-resolved fluorescence immunoassay for microcystin detection. Clin. Chim. Acta 2004, 348, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Akter, S.; Vehniäinen, M.; Kankaanpää, H.T.; Lamminmäki, U. Rapid and highly sensitive non-competitive immunoassay for specific detection of nodularin. Microorganisms 2017, 5, E58. [Google Scholar] [CrossRef] [PubMed]
- Kokociński, M.; Mankiewicz-Boczek, J.; Jurczak, T.; Spoof, L.; Meriluoto, J.; Rejmonczyk, E.; Hautala, H.; Vehniäinen, M.; Pawełczyk, J.; Soininen, J. Aphanizomenon gracile (Nostocales), a cylindrospermopsin-producing cyanobacterium in Polish lakes. Environ. Sci. Pollut. Res. Int. 2013, 20, 5243–5264. [Google Scholar] [CrossRef] [PubMed]
- Lipman, N.S.; Jackson, L.R.; Trudel, L.J.; Weis-Garcia, F. Monoclonal versus polyclonal antibodies: Distinguishing characteristics, applications, and information resources. ILAR J. 2005, 46, 258–268. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Samples | Nominal Value (ng mL−1) | Mean ± SD (ng mL−1) | CV (%) | Recovery (%) |
|---|---|---|---|---|---|
| Intra-assay (n = 8) | A | 2.0 | 1.91 ± 0.078 | 4.1 | 95.9 |
| B | 5.0 | 5.16 ± 0.24 | 4.6 | 103.2 | |
| C | 20.0 | 20.3 ± 1.18 | 5.8 | 101.5 | |
| Inter-assay (n = 12) | A | 2.0 | 1.96 ± 0.096 | 4.9 | 98.1 |
| B | 5.0 | 5.08 ± 0.24 | 4.8 | 101.7 | |
| C | 20.0 | 19.5 ± 12.7 | 6.5 | 97.4 |
| Spiked Value (ng mL−1) | Measured Value ± SD (ng mL−1) | Recovery (%) | CV (%) |
|---|---|---|---|
| 0.25 | 0.246 ± 0.04 | 98.6 | 16.38 |
| 5.0 | 5.44 ± 0.54 | 108.8 | 9.99 |
| 25 | 23.15 ± 1.91 | 92.6 | 8.24 |
| Sample | Dilution | Expected Value (ng mL−1) | Observed Value (ng mL−1, n = 3) | Recovery (%) |
|---|---|---|---|---|
| A | NA | 10.0 | ||
| 1:2 | 5.0 | 4.86 | 97.2 | |
| 1:4 | 2.5 | 2.41 | 96.4 | |
| 1:8 | 1.25 | 1.29 | 103.2 | |
| 1:16 | 0.62 | 0.66 | 106.5 | |
| B | NA | 50.0 | ||
| 1:2 | 25.0 | 25.4 | 101.6 | |
| 1:4 | 12.5 | 13.0 | 104.6 | |
| 1:8 | 6.25 | 5.91 | 94.5 | |
| 1:16 | 3.12 | 3.39 | 108.7 |
| Method | Recovery | Imprecision | Operating Time | Maximum Quantitative Value |
|---|---|---|---|---|
| TRFIA | 95.9–103.2% | 4.1–6.5% | 1 h | 50 ng mL−1 |
| ELISA (Beacon) | 80–120% | <20% | 1.5 h | 2 ng mL−1 |
| ELISA (Abraxis) | 98–108% | 4.3–8.3% | 1.25–1.5 h | 2 ng mL−1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, L.; Peng, L.; Yang, Y.; Han, B.-p. Development of Time-Resolved Fluoroimmunoassay for Detection of Cylindrospermopsin Using Its Novel Monoclonal Antibodies. Toxins 2018, 10, 255. https://doi.org/10.3390/toxins10070255
Lei L, Peng L, Yang Y, Han B-p. Development of Time-Resolved Fluoroimmunoassay for Detection of Cylindrospermopsin Using Its Novel Monoclonal Antibodies. Toxins. 2018; 10(7):255. https://doi.org/10.3390/toxins10070255
Chicago/Turabian StyleLei, Lamei, Liang Peng, Yang Yang, and Bo-ping Han. 2018. "Development of Time-Resolved Fluoroimmunoassay for Detection of Cylindrospermopsin Using Its Novel Monoclonal Antibodies" Toxins 10, no. 7: 255. https://doi.org/10.3390/toxins10070255
APA StyleLei, L., Peng, L., Yang, Y., & Han, B.-p. (2018). Development of Time-Resolved Fluoroimmunoassay for Detection of Cylindrospermopsin Using Its Novel Monoclonal Antibodies. Toxins, 10(7), 255. https://doi.org/10.3390/toxins10070255