High Production of LukMF’ in Staphylococcus aureus Field Strains Is Associated with Clinical Bovine Mastitis

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Prevalence of lukM-lukF’ Genes among Bovine Mastitis Isolates
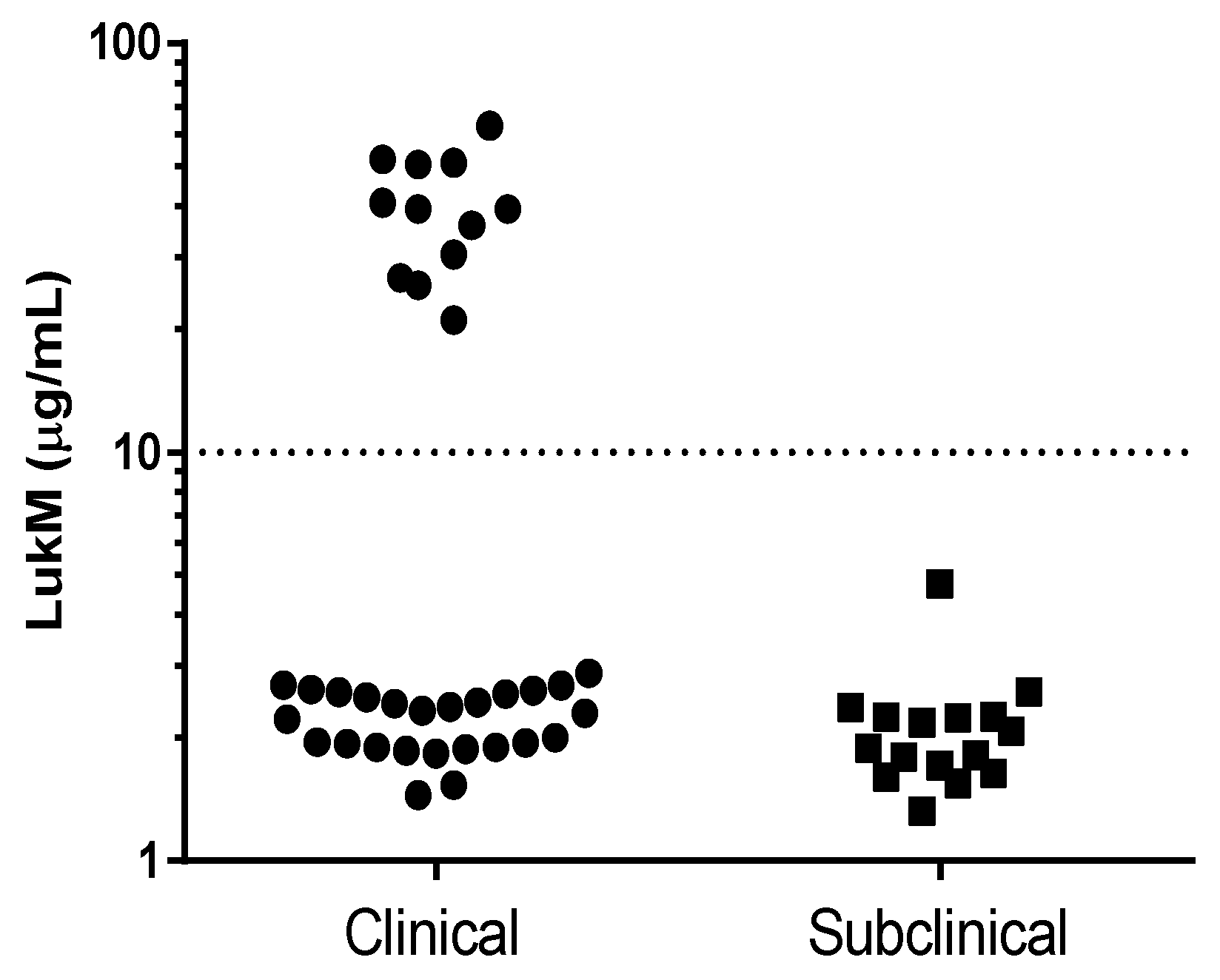
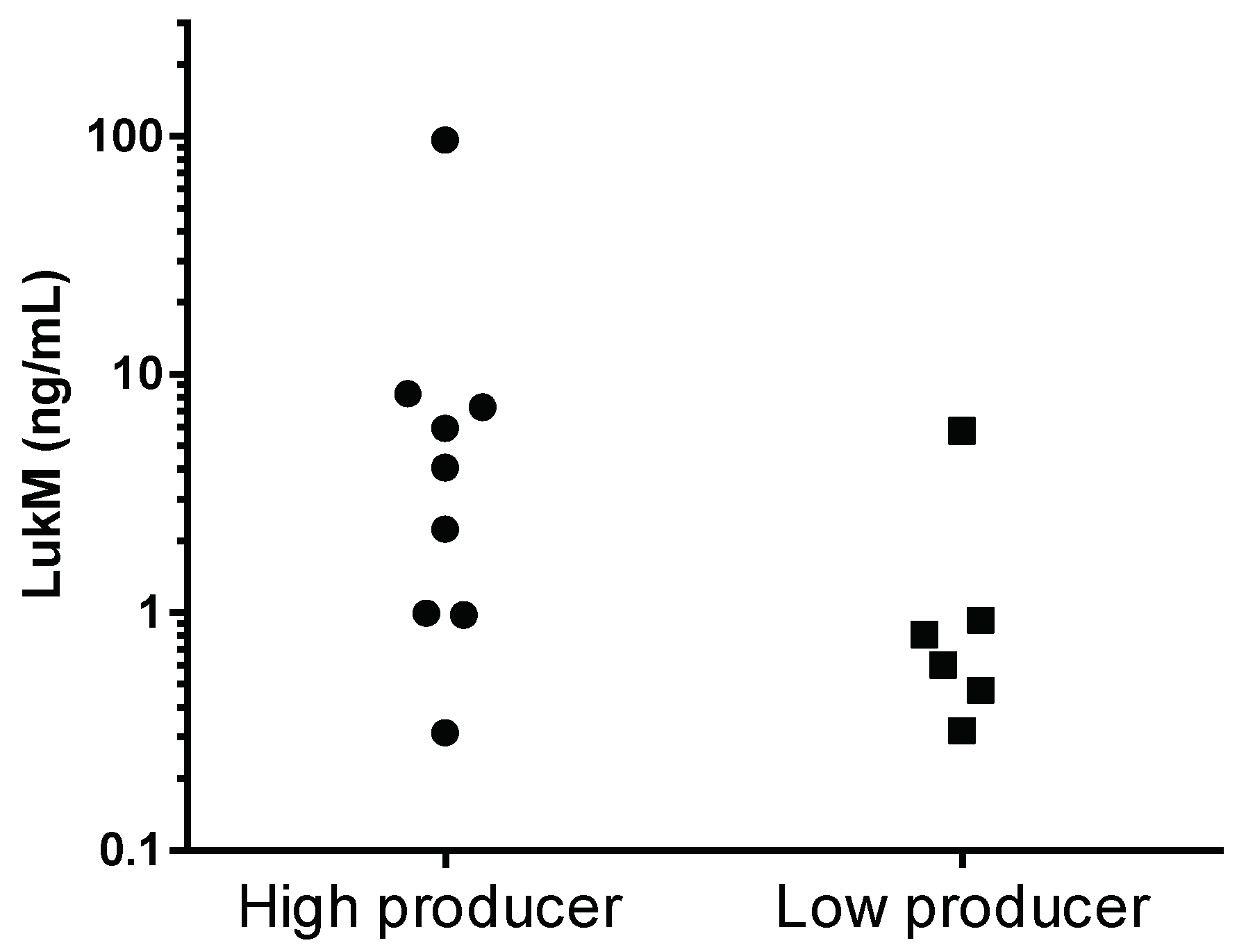
2.2. Production of LukM In Vitro and In Vivo
2.3. Genotyping of S. aureus Isolates
2.4. Analysis of the lukM-lukF’ Operon and saeS, saeR, rot Genes
3. Discussion
4. Materials and Methods
4.1. Collection of S. aureus Bovine Mastitis Isolates
4.2. DNA Extraction and Amplification of lukM, lukF’, femA and rot Genes
4.3. In Vitro and In Vivo LukM Production
4.4. Genotyping of Mastitis Strains and Genomic Analyses
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hogeveen, H.; Huijps, K.; Lam, T.J.G.M. Economic aspects of mastitis: New developments. N. Z. Vet. J. 2011, 59, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Halasa, T.; Huijps, K.; Østerås, O.; Hogeveen, H. Economic effects of bovine mastitis and mastitis management: A review. Vet. Q. 2007, 29, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Fogsgaard, K.K.; Bennedsgaard, T.W.; Herskin, M.S. Behavioral changes in freestall-housed dairy cows with naturally occurring clinical mastitis. J. Dairy Sci. 2015, 98, 1730–1738. [Google Scholar] [CrossRef] [PubMed]
- Peton, V.; Le Loir, Y. Staphylococcus aureus in veterinary medicine. Infect. Genet. Evol. 2014, 21, 602–615. [Google Scholar] [CrossRef] [PubMed]
- Wellnitz, O.; Bruckmaier, R.M. The innate immune response of the bovine mammary gland to bacterial infection. Vet. J. 2012, 192, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Koymans, K.J.; Vrieling, M.; Gorham, R.D., Jr.; van Strijp, J.A. Staphylococcal Immune Evasion Proteins: Structure, Function, and Host Adaptation. Curr. Top. Microbiol. Immunol. 2017, 409, 441–489. [Google Scholar] [PubMed]
- Thammavongsa, V.; Kim, H.K.; Missiakas, D.; Schneewind, O. Staphylococcal manipulation of host immune responses. Nat. Rev. Microbiol. 2015, 13, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, A.R.; Salgado-Pabón, W.; Kohler, P.L.; Horswill, A.R.; Leung, D.Y.M.; Schlievert, P.M. Staphylococcal and streptococcal superantigen exotoxins. Clin. Microbiol. Rev. 2013, 26, 422–447. [Google Scholar] [CrossRef] [PubMed]
- Bukowski, M.; Wladyka, B.; Dubin, G. Exfoliative toxins of Staphylococcus aureus. Toxins 2010, 2, 1148–1165. [Google Scholar] [CrossRef] [PubMed]
- O’Riordan, K.; Lee, J.C. Staphylococcus aureus Capsular Polysaccharides. Clin. Microbiol. Rev. 2004, 17, 218–234. [Google Scholar] [CrossRef] [PubMed]
- DuMont, A.L.; Torres, V.J. Cell targeting by the Staphylococcus aureus pore-forming toxins: It’s not just about lipids. Trends Microbiol. 2014, 22, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Deghorain, M.; Van Melderen, L. The staphylococci phages family: An overview. Viruses 2012, 4, 3316–3335. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, F., 3rd; Torres, V.J. The bicomponent pore-forming leucocidins of Staphylococcus aureus. Microbiol. Mol. Biol. Rev. 2014, 78, 199–230. [Google Scholar] [CrossRef] [PubMed]
- Koop, G.; Vrieling, M.; Storisteanu, D.M.L.; Lok, L.S.C.; Monie, T.; Van Wigcheren, G.; Raisen, C.; Ba, X.; Gleadall, N.; Hadjirin, N.; et al. Identification of LukPQ, a novel, equid-adapted leukocidin of Staphylococcus aureus. Sci. Rep. 2017, 7, 40660. [Google Scholar] [CrossRef] [PubMed]
- Bar-Gal, G.K.; Blum, S.E.; Hadas, L.; Ehricht, R.; Monecke, S.; Leitner, G. Host-specificity of Staphylococcus aureus causing intramammary infections in dairy animals assessed by genotyping and virulence genes. Vet. Microbiol. 2015, 176, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Schlotter, K.; Ehricht, R.; Hotzel, H.; Monecke, S.; Pfeffer, M.; Donat, K. Leukocidin genes lukF-P83 and lukM are associated with Staphylococcus aureus clonal complexes 151, 479 and 133 isolated from bovine udder infections in Thuringia, Germany. Vet. Res. 2012, 43. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Tochimaru, N.; Nakasuji, S.; Hata, E.; Kobayashi, H.; Eguchi, M.; Kaneko, J.; Kamio, Y.; Kaidoh, T.; Takeuchi, S. Leukotoxin family genes in Staphylococcus aureus isolated from domestic animals and prevalence of lukM-lukF-PV genes by bacteriophages in bovine isolates. Vet. Microbiol. 2005, 110, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Fromageau, A.; Gilbert, F.B.; Prévost, G.; Rainard, P. Binding of the Staphylococcus aureus leucotoxin LukM to its leucocyte targets. Microb. Pathog. 2010, 49, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Vrieling, M.; Koymans, K.J.; Heesterbeek, D.A.; Aerts, P.C.; Rutten, V.P.; de Haas, C.J.; van Kessel, K.P.M.; Koets, A.P.; Nijland, R.; van Strijp, J.A.G. Bovine Staphylococcus aureus Secretes the Leukocidin LukMF' To Kill Migrating Neutrophils through CCR1. MBio 2015, 6, e00335-15. [Google Scholar] [CrossRef] [PubMed]
- Rainard, P.; Riollet, C. Innate immunity of the bovine mammary gland. Vet. Res. 2006, 37, 369–400. [Google Scholar] [CrossRef] [PubMed]
- Vrieling, M.; Boerhout, E.M.; van Wigcheren, G.F.; Koymans, K.J.; Mols-Vorstermans, T.G.; de Haas, C.J.; Aerts, P.C.; Daemen, I.J.J.M.; van Kessel, K.P.M.; Koets, A.P.; et al. LukMF’ is the major secreted leukocidin of bovine Staphylococcus aureus and is produced in vivo during bovine mastitis. Sci. Rep. 2016, 25, 37759. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liang, Y.; Lynch, K.H.; Dennis, J.J.; Wishart, D.S. PHAST: A Fast Phage Search Tool. Nucleic Acids Res. 2011, 39 (Suppl. 2), W347–W352. [Google Scholar] [CrossRef] [PubMed]
- Mootz, J.M.; Benson, M.A.; Heim, C.E.; Crosby, H.A.; Kavanaugh, J.S.; Dunman, P.M.; Kielian, T.; Torres, V.J.; Horswill, A.R. Rot is a key regulator of Staphylococcus aureus biofilm formation. Mol. Microbiol. 2015, 96, 388–404. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, C.P.; Boyle-Vavra, S.; Daum, R.S. Importance of the global regulators agr and SaeRS in the pathogenesis of CA-MRSA USA300 infection. PLoS ONE 2010, 5, e15177. [Google Scholar] [CrossRef] [PubMed]
- Haran, K.P.; Godden, S.M.; Boxrud, D.; Jawahir, S.; Bender, J.B.; Sreevatsan, S. Prevalence and characterization of Staphylococcus aureus, including methicillin-resistant Staphylococcus aureus, isolated from bulk tank milk from Minnesota dairy farms. J. Clin. Microbiol. 2012, 50, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Veh, K.A.; Klein, R.C.; Ster, C.; Keefe, G.; Lacasse, P.; Scholl, D.; Roy, J.P.; Haine, D.; Dufour, S.; Talbot, B.G.; et al. Genotypic and phenotypic characterization of Staphylococcus aureus causing persistent and nonpersistent subclinical bovine intramammary infections during lactation or the dry period. J. Dairy Sci. 2015, 98, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Johler, S.; Layer, F.; Stephan, R. Comparison of virulence and antibiotic resistance genes of food poisoning outbreak isolates of Staphylococcus aureus with isolates obtained from bovine mastitis milk and pig carcasses. J. Food Prot. 2011, 74, 1852–1859. [Google Scholar] [CrossRef] [PubMed]
- Ikawaty, R.; Brouwer, E.C.; Jansen, M.D.; van Duijkeren, E.; Mevius, D.; Verhoef, J.; Fluit, A.C. Characterization of Dutch Staphylococcus aureus from bovine mastitis using a Multiple Locus Variable Number Tandem Repeat Analysis. Vet. Microbiol. 2009, 136, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Merz, A.; Stephan, R.; Johler, S. Staphylococcus aureus isolates from goat and sheep milk seem to be closely related and differ from isolates detected from bovine milk. Front. Microbiol. 2016, 7, 319. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Kock, M.M.; Ehlers, M.M. Molecular characterization of Staphylococcus aureus isolated from bovine mastitis and close human contacts in South African dairy herds: Genetic diversity and inter-species host transmission. Front. Microbiol. 2017, 8, 511. [Google Scholar] [CrossRef] [PubMed]
- Hata, E.; Katsuda, K.; Kobayashi, H.; Uchida, I.; Tanaka, K.; Eguchi, M. Genetic variation among Staphylococcus aureus strains from bovine milk and their relevance to methicillin-resistant isolates from humans. J. Clin. Microbiol. 2010, 48, 2130–2139. [Google Scholar] [CrossRef] [PubMed]
- Artursson, K.; Söderlund, R.; Liu, L.; Monecke, S.; Schelin, J. Genotyping of Staphylococcus aureus in bovine mastitis and correlation to phenotypic characteristics. Vet. Microbiol. 2016, 193, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Monecke, S.; Kuhnert, P.; Hotzel, H.; Slickers, P.; Ehricht, R. Microarray based study on virulence-associated genes and resistance determinants of Staphylococcus aureus isolates from cattle. Vet. Microbiol. 2007, 125, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Conceição, T.; De Lencastre, H.; Aires-De-Sousa, M. Healthy Bovines as Reservoirs of Major Pathogenic Lineages of Staphylococcus aureus in Portugal. Microb. Drug Resist. 2017, 23, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Killikelly, A.; Benson, M.A.; Ohneck, E.A.; Sampson, J.M.; Jakoncic, J.; Spurrier, B.; Torres, V.J.; Kong, X.-P. Structure-based functional characterization of repressor of toxin (Rot), a central regulator of Staphylococcus aureus virulence. J. Bacteriol. 2015, 197, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Benson, M.A.; Ohneck, E.A.; Ryan, C.; Alonzo, F.; Smith, H.; Narechania, A.; Kolokotronis, S.O.; Satola, S.W.; Uhlemann, A.C.; Sebra, R. Evolution of hypervirulence by a MRSA clone through acquisition of a transposable element. Mol. Microbiol. 2014, 93, 664–681. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, C.; Wei, X.; Zhao, X.; Zhong, X. Loop-mediated isothermal amplification for detection of Staphylococcus aureus in dairy cow suffering from mastitis. J. Biomed. Biotechnol. 2012, 2012, 435982. [Google Scholar]
- Britten, A.M. The role of diagnostic microbiology in mastitis control programs. Vet. Clin. N. Am. Food Anim. Pract. 2012, 28, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Hogan, J.S.; National Mastitis Council (U.S.). Laboratory Handbook on Bovine Mastitis; National Mastitis Council: Madison, WI, USA, 1999. [Google Scholar]
- Francois, P.; Pittet, D.; Bento, M.; Pepey, B.; Vaudaux, P.; Lew, D.; Schrenzel, J. Rapid detection of methicillin-resistant Staphylococcus aureus directly from sterile or nonsterile clinical samples by a new molecular assay. J. Clin. Microbiol. 2003, 41, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbakc, L.; Sicheritz-Ponténa, T.; Usserya, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Spa-Type | Isolates N | Clinical Mastitis N (%) | lukM-lukF’ Postive Isolates N (%) 1 | High LukMF’ Production Isolates N (%) 2 | Clonal Complex 3 |
|---|---|---|---|---|---|
| t529 | 40 | 23 (58) | 40 (100) | 0 (0) | CC151 |
| t543 | 12 | 12 (100) | 12 (100) | 12 (100) | CC479 |
| t524 | 1 | 1 (100) | 0 (0) | 0 (0) | ND 4 |
| t1403 | 1 | 1 (100) | 1 (100) | 0 (0) | CC133 |
| t015 | 1 | 0 (0) | 0 (0) | 0 (0) | ND 4 |
| Total | 55 | 37 (67) | 53 (96) | 12 (22) |
| Gene | Sequence | Product Size (bp) | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|
| femA | f: 5′-tgcctttacagatagcatgcca-3′ | 142 | 59.5 | [40] |
| r: 5′-agtaagtaagcaagctgcaatgacc-3′ | ||||
| lukM | f: 5′-aaacgcgcagttaataaaaag-3′ | 975 | 55 | This study |
| r: 5′-agcattaggtcctcttgtcg-3′ | ||||
| lukF’ | f: 5′-actcaggctatacccaaccca-3′ | 472 | 59.5 | This study |
| r: 5′-cgagctactctgtctgccac-3′ | ||||
| rot | f: 5′-accaatttagcctcattcggtttg-3′ | 705 | 55 | This study |
| r: 5′-catcgtcaacaggacgctct-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoekstra, J.; Rutten, V.; Sommeling, L.; Van Werven, T.; Spaninks, M.; Duim, B.; Benedictus, L.; Koop, G. High Production of LukMF’ in Staphylococcus aureus Field Strains Is Associated with Clinical Bovine Mastitis. Toxins 2018, 10, 200. https://doi.org/10.3390/toxins10050200
Hoekstra J, Rutten V, Sommeling L, Van Werven T, Spaninks M, Duim B, Benedictus L, Koop G. High Production of LukMF’ in Staphylococcus aureus Field Strains Is Associated with Clinical Bovine Mastitis. Toxins. 2018; 10(5):200. https://doi.org/10.3390/toxins10050200
Chicago/Turabian StyleHoekstra, Jurriaan, Victor Rutten, Laura Sommeling, Tine Van Werven, Mirlin Spaninks, Birgitta Duim, Lindert Benedictus, and Gerrit Koop. 2018. "High Production of LukMF’ in Staphylococcus aureus Field Strains Is Associated with Clinical Bovine Mastitis" Toxins 10, no. 5: 200. https://doi.org/10.3390/toxins10050200
APA StyleHoekstra, J., Rutten, V., Sommeling, L., Van Werven, T., Spaninks, M., Duim, B., Benedictus, L., & Koop, G. (2018). High Production of LukMF’ in Staphylococcus aureus Field Strains Is Associated with Clinical Bovine Mastitis. Toxins, 10(5), 200. https://doi.org/10.3390/toxins10050200