Warming Affects Growth Rates and Microcystin Production in Tropical Bloom-Forming Microcystis Strains



Abstract
1. Introduction
2. Results
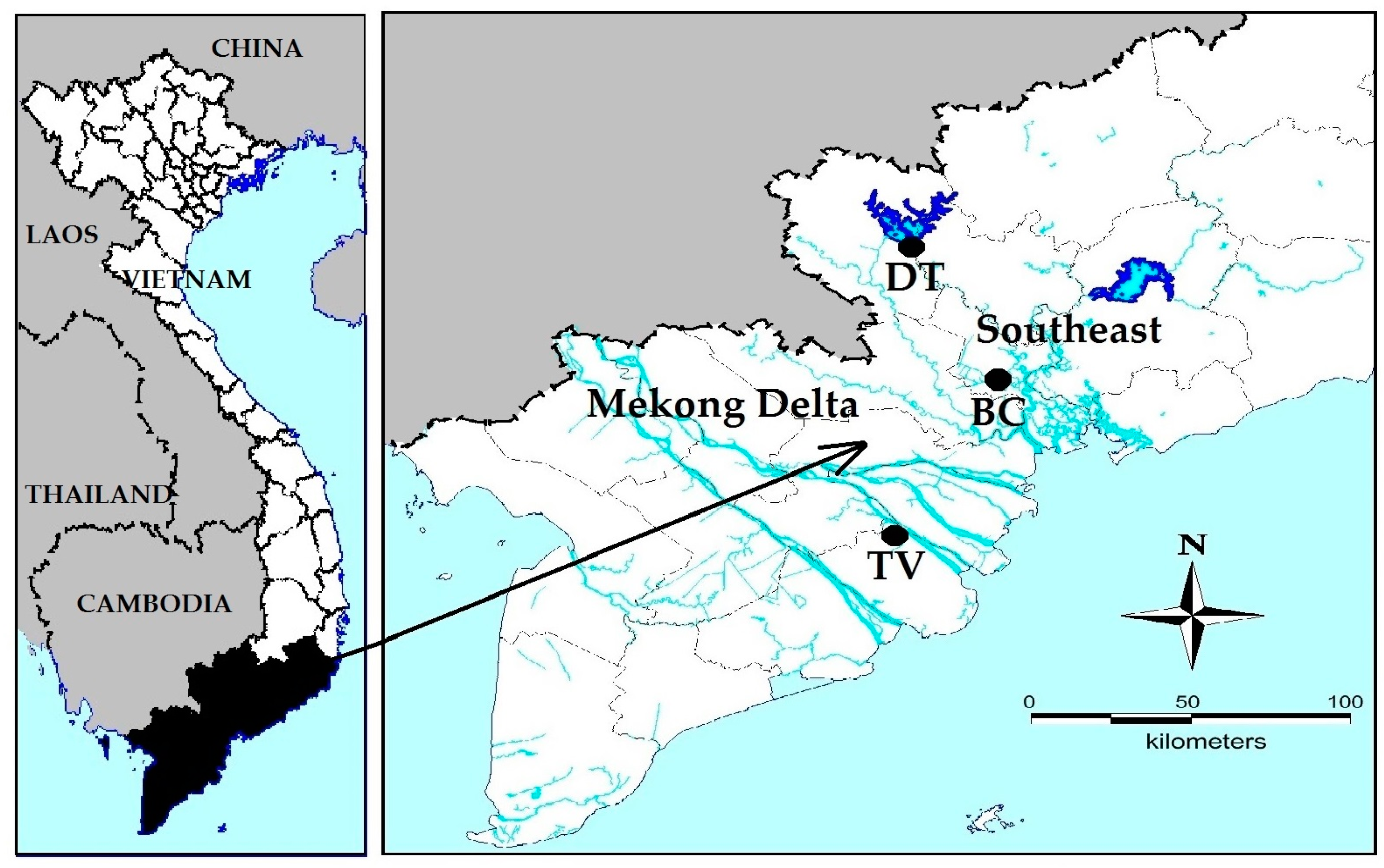
2.1. Water Bodies in Which Microcystis Strains Were Isolated
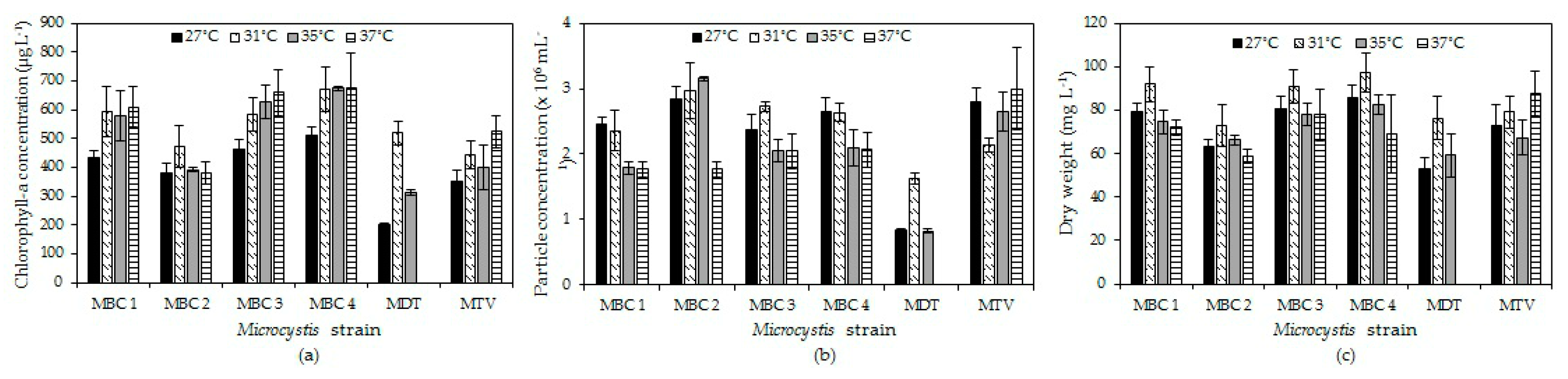
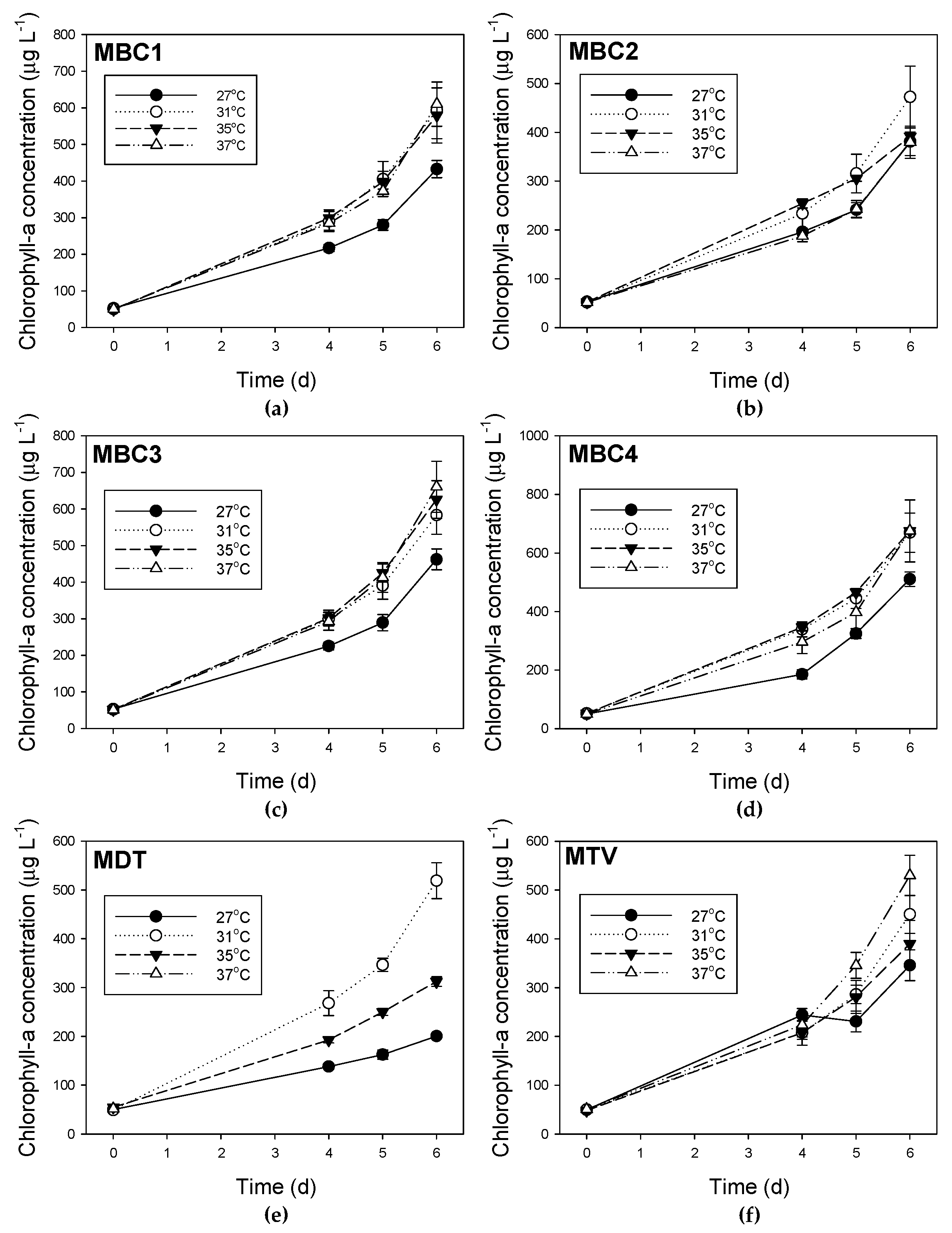
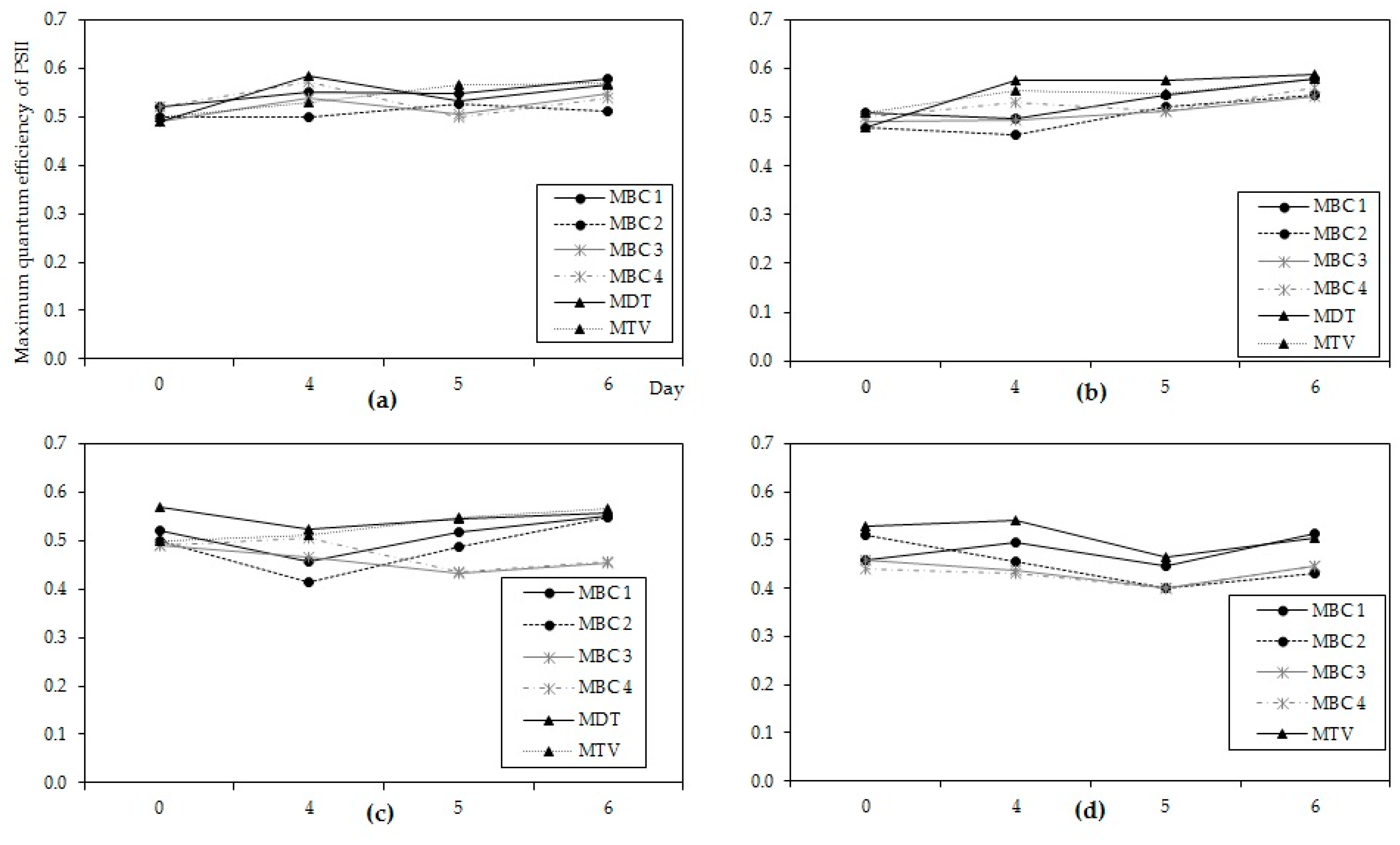
2.2. Effects of Temperature on Microcystis Biomass Indicators
2.3. Effects of Temperature on Microcystis Growth Rates
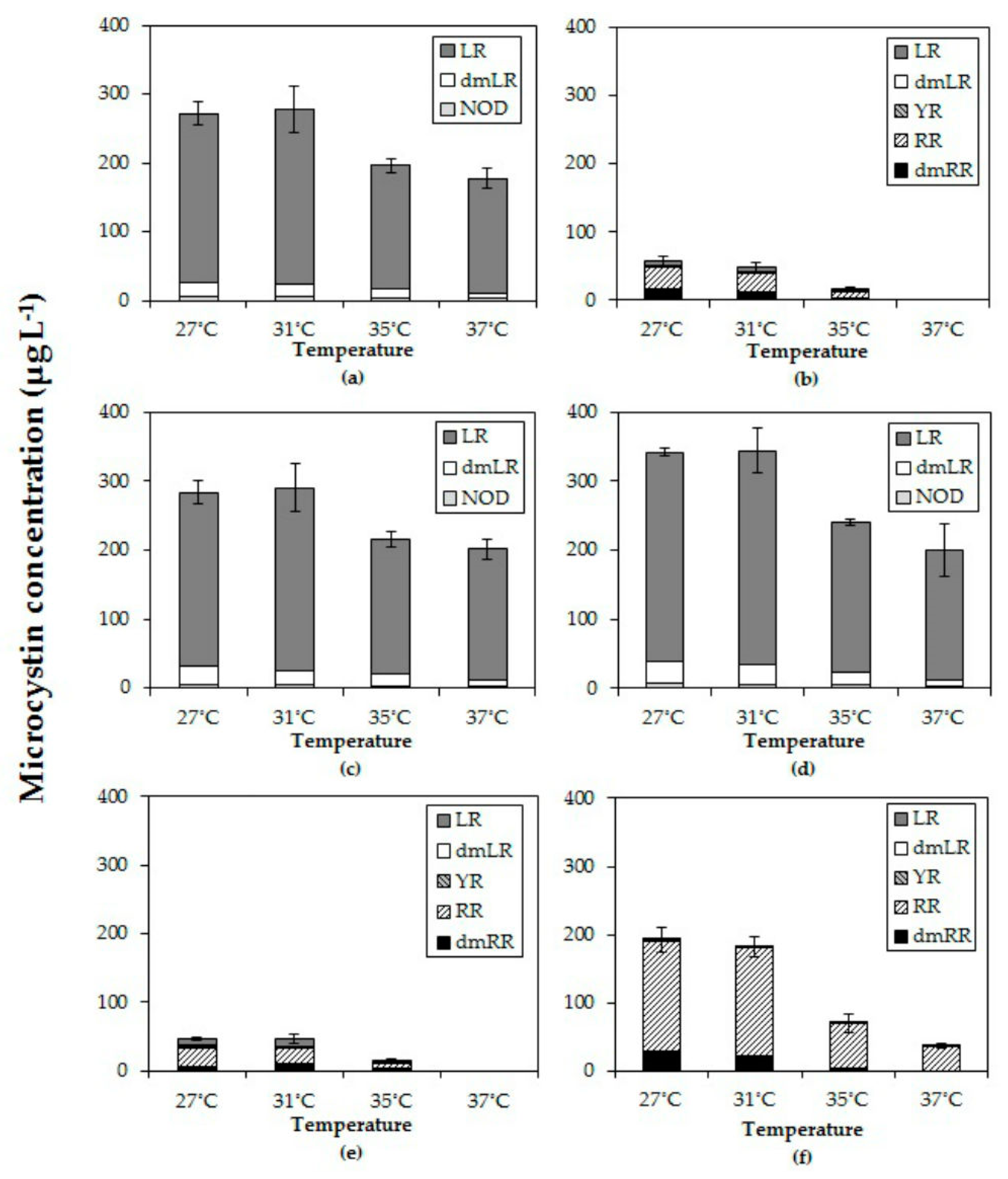
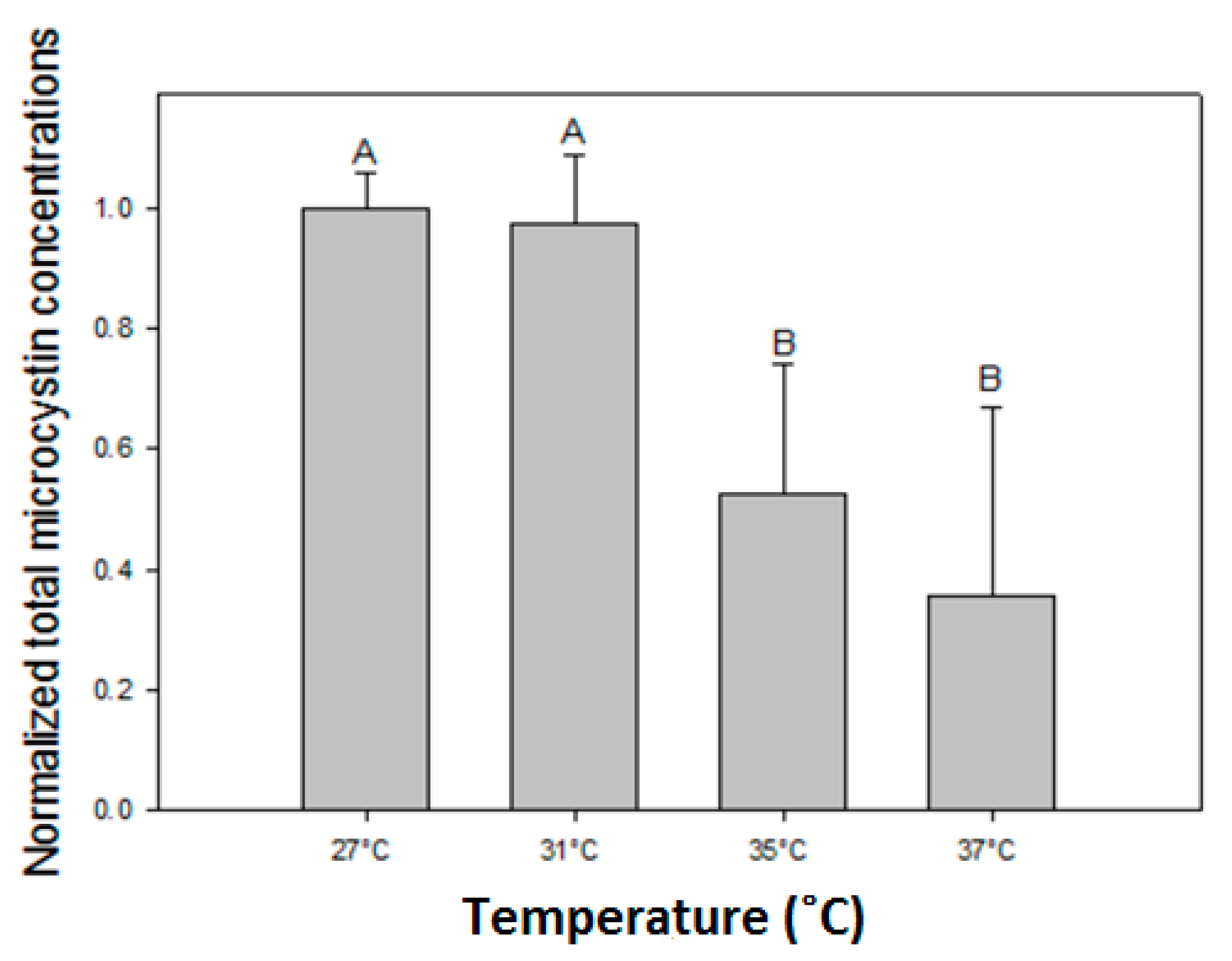
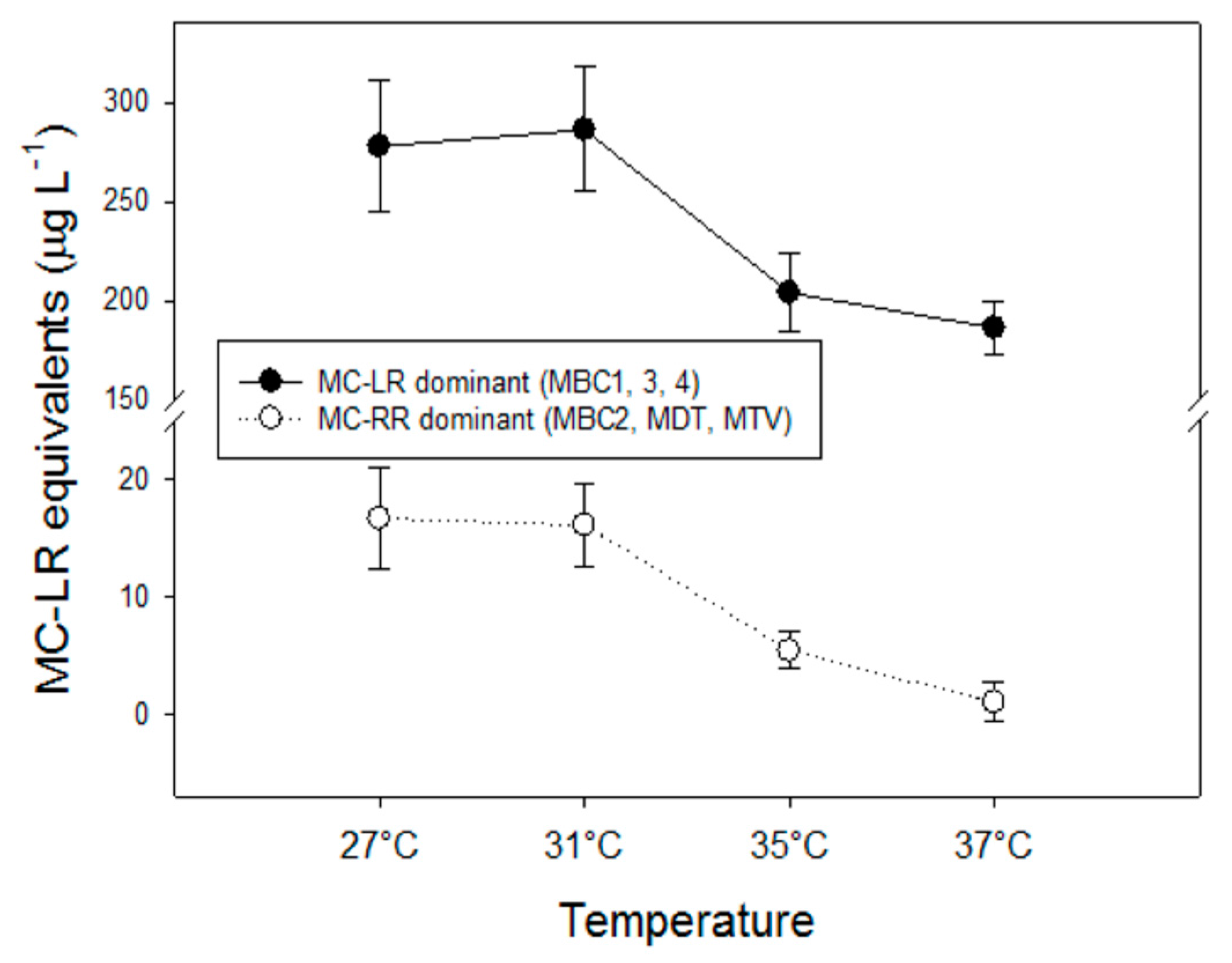
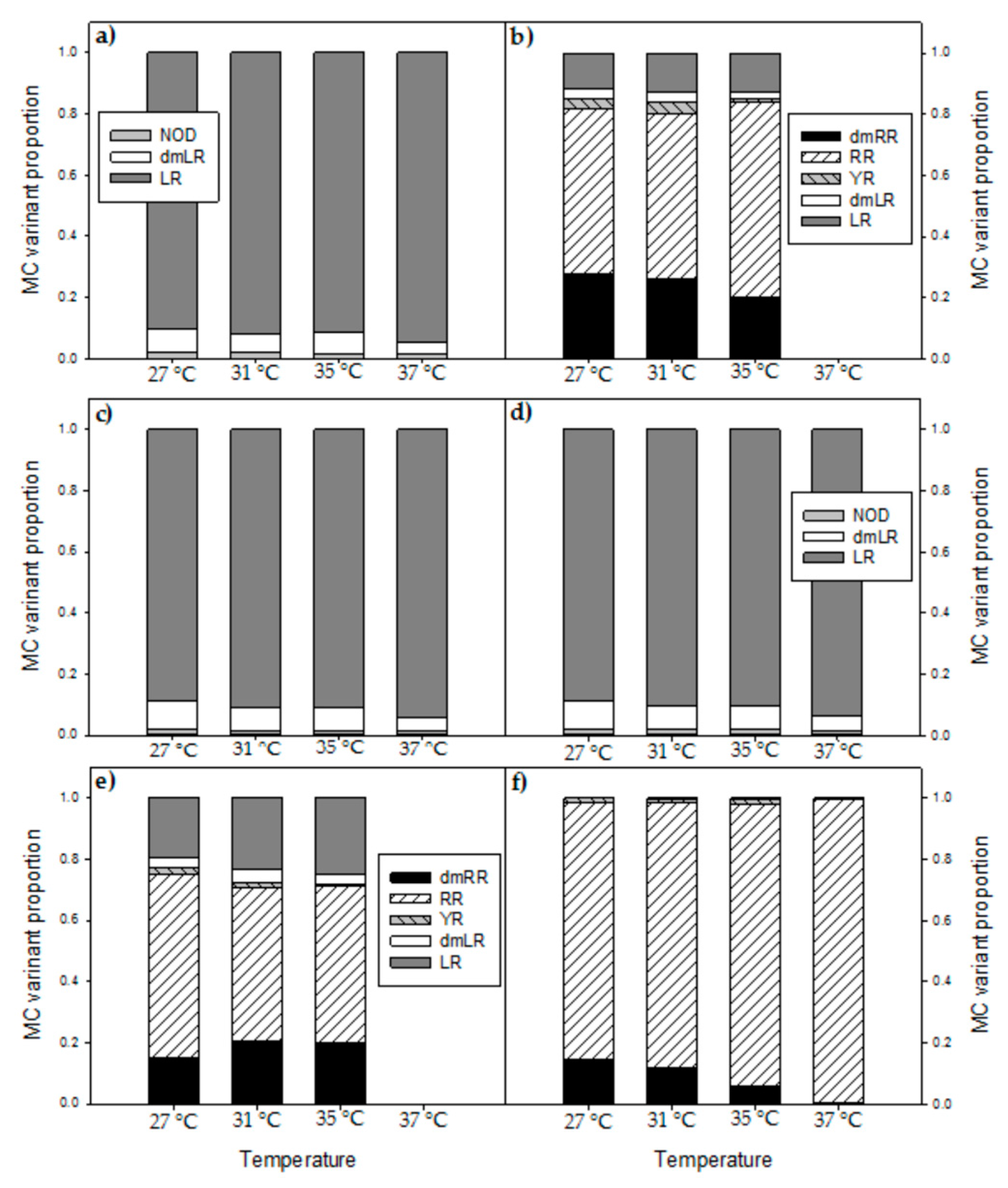
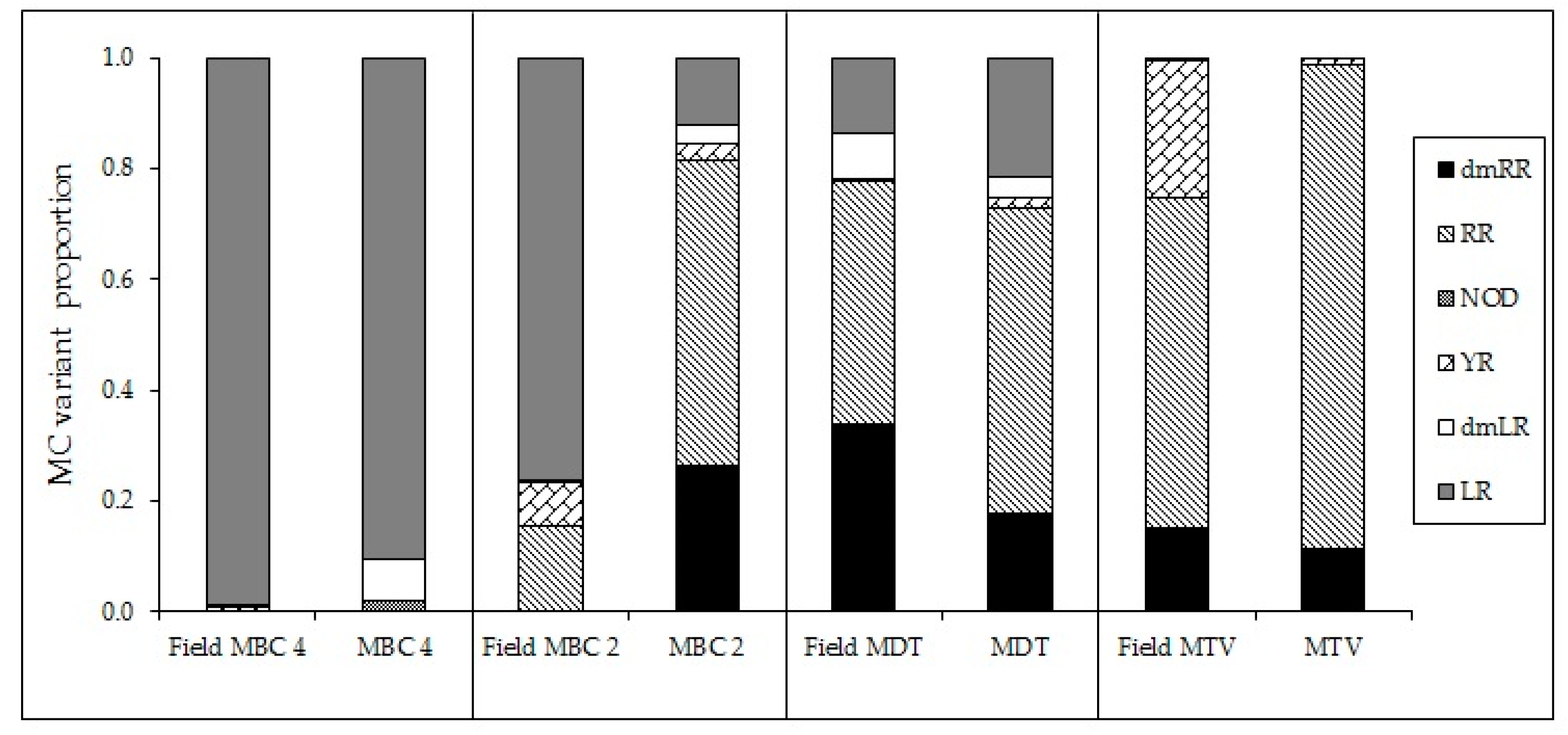
2.4. Effects of Temperature on Microcystins
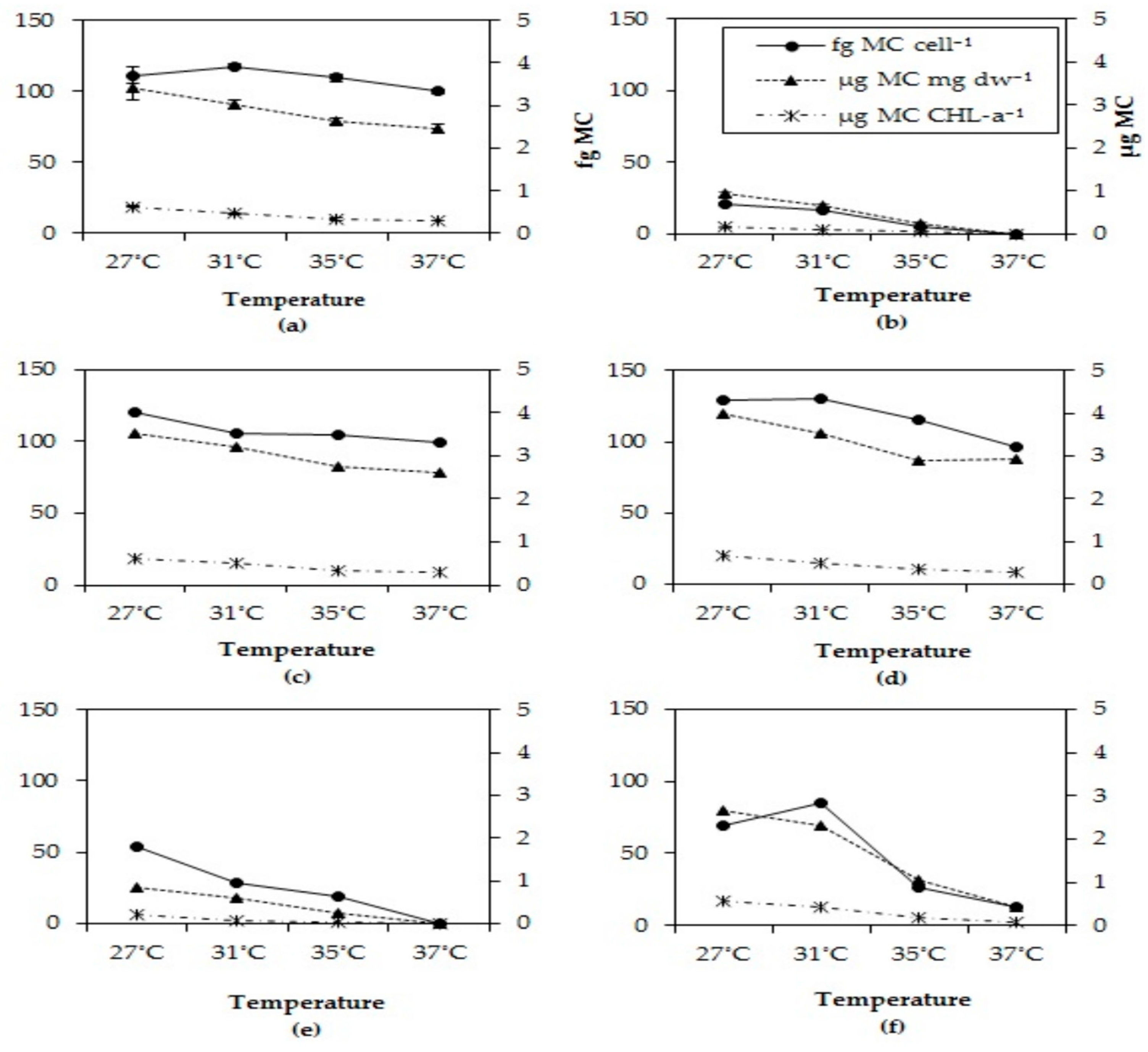
2.5. Microcystin Cell Quota
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Microcystis Strains
5.2. Growth Experiment
5.3. Microcystin (MC) Analysis
5.4. Data Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

Appendix B

Appendix C

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MC-LR | Microcystis Strains | |||||
| Temp. | MBC1 | MBC2 | MBC3 | MBC4 | MDT | MTV |
| 27 °C | A | A | A | A | A | A |
| 31 °C | B | A | B | B | AB | A |
| 35 °C | B | A | B | B | B | A |
| 37 °C | C | B | C | C | --- | A |
| F3,11 value | 333.2 | 187.8 | 290.1 | 153.6 | 6.84 | 0.75 |
| p-value | p < 0.001 | p < 0.001 | p = 0.001 | p < 0.001 | p = 0.028 | p = 0.551 |
| Normality | p = 0.759 | p = 0.085 | p = 0.605 | p = 0.219 | p = 0.967 | p = 0.217 |
| Equal Var. | p = 0.256 | p = 0.063 | p = 0.605 | p = 0.654 | p = 0.294 | p = 0.480 |
| MC-RR | Microcystis Strains | |||||
| Temp. | MBC1 | MBC2 @ | MBC3 | MBC4 | MDT | MTV # |
| 27 °C | --- | A | --- | --- | A | A |
| 31 °C | --- | A | --- | --- | B | AB |
| 35 °C | --- | B | --- | --- | B | AB |
| 37 °C | --- | --- | --- | --- | --- | B |
| F3,11 value | --- | 50.70 | --- | --- | 55.65 | H3 = 10.4 |
| p-value | --- | p < 0.001 | --- | --- | p = 0.028 | p = 0.015 |
| Normality | --- | p = 0.168 | --- | --- | p = 0.300 | p < 0.050 |
| Equal Var. | --- | p = 0.190 | --- | --- | p = 0.205 | --- |

Appendix D
| fg MC cell−1 | Microcystis Strains | |||||
| Temp. | MBC1 | MBC2 | MBC3 | MBC4 | MDT | MTV |
| 27 °C | A | A | A | A | A | A |
| 31 °C | A | B | AB | A | B | B |
| 35 °C | A | C | AB | AB | C | C |
| 37 °C | B | D | B | B | D | D |
| F3,11 value | 13.76 | 481.3 | 5.44 | 7.26 | 528.5 | 535.3 |
| p-value | p = 0.002 | p < 0.001 | p = 0.025 | p = 0.011 | p < 0.001 | p < 0.001 |
| Normality | p = 0.746 | p = 0.197 | p = 0.256 | p = 0.136 | p = 0.211 | p = 0.597 |
| Equal Var. | p = 0.289 | p = 0.129 | p = 0.928 | p = 0.754 | p = 0.659 | p = 0.211 |
| µg MC µg CHLa−1 | Microcystis Strains | |||||
| Temp. | MBC1 | MBC2 | MBC3 | MBC4 | MDT # | MTV |
| 27 °C | A | A | A | A | A | A |
| 31 °C | B | B | B | B | AB | B |
| 35 °C | C | C | C | C | AB | C |
| 37 °C | D | D | D | D | B | D |
| F3,11 value | 137.5 | 457.5 | 274.4 | 250.2 | H3 = 10.5 | 1167.7 |
| p-value | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p = 0.015 | p < 0.001 |
| Normality | p = 0.322 | p = 0.235 | p = 0.755 | p = 0.714 | p = 0.159 | p = 0.443 |
| Equal Var. | p = 0.225 | p = 0.135 | p = 0.768 | p = 0.079 | p < 0.050 | p = 0.241 |
| µg MC mg DW−1 | Microcystis Strains | |||||
| Temp. | MBC1 | MBC2 | MBC3 | MBC4 | MDT | MTV |
| 27 °C | A | A | A | A | A | A |
| 31 °C | B | B | AB | AB | AB | B |
| 35 °C | C | C | BC | B | AB | C |
| 37 °C | C | D | C | B | B | D |
| F3,11 value | 20.77 | 396.0 | 11.11 | 9.77 | 103.2 | 151.9 |
| p-value | p < 0.001 | p < 0.001 | p = 0.003 | p = 0.005 | p < 0.001 | p < 0.001 |
| Normality | p = 0.741 | p = 0.269 | p = 0.915 | p = 0.091 | p = 0730 | p = 0.799 |
| Equal Var. | p = 0.084 | p = 0.685 | p = 0.430 | p = 0.626 | p = 0.561 | p = 0.390 |
Appendix E
| Species | T Range | T Opt | µmax (d−1) | Reference |
|---|---|---|---|---|
| M. aeruginosa | 20–35 | 30 | 0.45 | [68] |
| M. aeruginosa | 20–35 | 32 | 1.6 | [56] |
| M. aeruginosa | 16–36 | 32 | 0.81 | [25] |
| M. aeruginosa | 15–30 | 25 | 0.19 | [69] |
| M. aeruginosa | 5–25 | 25 | 0.36 | [70] |
| M. aeruginosa | 10–30 | 27.5 | 0.8 | [71] |
| M. aeruginosa | 10–45 | 35 | 1.06 | [72] |
| M. aeruginosa | 20–35 | 32.5 | 1.12 | [73] |
| M. aeruginosa | 20–35 | 30 | 0.94 | [73] |
| M. flos-aquae | 27–36 | 30 | 0.81 | [28] |
| M. ichthyoblabe | 27–36 | 33 | 0.94 | [28] |
| M. ichthyoblabe | 27–36 | 33 | 0.77 | [28] |
| M. aeruginosa | 27–36 | 36 | 0.65 | [28] |
| M. viridis | 27–36 | 30 | 0.49 | [28] |
| M. aeruginosa | 18–30 | 30 | 0.4 | [74] |
| M. aeruginosa | 15–40 | 28.3 | 0.38 | [39] |
| M. aeruginosa | 15–40 | 28.8 | 0.41 | [39] |
| M. aeruginosa | 15–40 | 34.1 | 0.62 | [39] |
| M. aeruginosa | 15–40 | 28 | 0.29 | [39] |
| M. aeruginosa | 15–35 | 30 | 0.28 | [39] |
| M. aeruginosa | 15–40 | 35 | 1.02 | [39] |
| Average | 30.7 | 0.69 | ||
| SD | 3.1 | 0.35 |
Appendix F

Appendix G


Appendix H

References
- De Figueiredo, D.R.; Azeiteiro, U.M.; Esteves, S.M.; Gonçalves, F.J.M.; Pereira, M.J. Microcystin-producing blooms—A serious global public health issue. Ecotoxicol. Environ. Saf. 2004, 59, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci. Total Environ. 2011, 409, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Jöhnk, K.D.; Huisman, J.; Sharples, J.P.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef]
- Wagner, C.; Adrian, R. Cyanobacteria dominance: Quantifying the effect of climate change. Limnol. Oceanogr. 2009, 54, 2460–2468. [Google Scholar] [CrossRef]
- Rigosi, A.; Carey, C.C.; Ibelings, B.W.; Brookes, J.D. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnol. Oceanogr. 2014, 59, 99–114. [Google Scholar] [CrossRef]
- De Senerpont Domis, L.N.; Van de Waal, D.B.; Helmsing, N.R.; Van Donk, E.; Mooij, W.M. Community stoichiometry in a changing world: Combined effects of warming and eutrophication on phytoplankton dynamics. Ecology 2014, 95, 1485–1495. [Google Scholar] [CrossRef]
- Moss, B.; Kosten, S.; Meerhoff, M.; Battarbee, R.W.; Jeppesen, E.; Mazzeo, N.; Havens, K.; Lacerot, G.; Liu, Z.; De Meester, L.; et al. Allied attack: Climate change and eutrophication. Inland Waters 2011, 1, 101–105. [Google Scholar] [CrossRef]
- De Senerpont Domis, L.N.; Elser, J.J.; Gsell, A.; Huszar, V.L.M.; Ibelings, B.W.; Jeppesen, E.; Kosten, S.; Mooij, W.M.; Roland, F.; Sommer, U.; et al. Plankton dynamics under different climatic conditions in space and time. Freshw. Biol. 2013, 58, 463–482. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W. Chapter 10: Nutrient and other environmental controls of harmful cyanobacterial blooms along the freshwater-marine continuum. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Advances in Experimental Medicine and Biology; Hudnell, K.E., Ed.; Springer: New York, NY, USA, 2008; Volume 619, pp. 217–237. [Google Scholar]
- Dittmann, E.; Wiegand, C. Cyanobacterial toxins—Occurrence, biosynthesis and impact on human affairs. Mol. Nutr. Food Res. 2006, 50, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Funari, E.; Testai, E. Human health risk assessment related to cyanotoxins exposure. Crit. Rev. Toxicol. 2008, 38, 97–125. [Google Scholar] [CrossRef] [PubMed]
- Backer, L.C.; Manassaram-Baptiste, D.; LePrell, R.; Bolton, B. Cyanobacteria and Algae Blooms: Review of Health and Environmental Data from the Harmful Algal Bloom-Related Illness Surveillance System (HABISS) 2007–2011. Toxins 2015, 7, 1048–1064. [Google Scholar] [CrossRef] [PubMed]
- Harke, M.; Steffen, M.; Gobler, C.; Otten, T.; Wilhelm, S.; Wood, S.A.; Pearl, H. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Meriluoto, J.; Spoof, L.; Codd, G.A. Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons, Ltd.: Chichester, UK, 2017; p. 576. ISBN 9781119068747. [Google Scholar]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Dziallas, C.; Grossart, H.-P. Temperature and biotic factors influence bacterial communities associated with the cyanobacterium Microcystis sp. Environ. Microbiol. 2011, 13, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- Lehman, P.W.; Kurobe, T.; Lesmeister, S.; Baxa, D.; Tung, A.; Teh, S.J. Impacts of the 2014 severe drought on the Microcystis bloom in San Francisco Estuary. Harmful Algae 2017, 63, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Li, D.-M.; Zheng, H.-Y.; Pan, J.-L.; Zhang, T.-Q.; Tang, S.-K.; Lu, J.-M.; Zhong, L.-Q.; Liu, Y.-S.; Liu, X.-W. Seasonal dynamics of photosynthetic activity, Microcystis genotypes and microcystin production in Lake Taihu, China. J. Gt. Lakes Res. 2017, 43, 710–716. [Google Scholar] [CrossRef]
- Rinta-Kanto, J.M.; Konopko, E.A.; DeBruyn, J.M.; Bourbonniere, R.A.; Boyer, G.L.; Wilhelm, S.W. Lake Erie Microcystis: Relationship between microcystin production, dynamics of genotypes and environmental parameters in a large lake. Harmful Algae 2009, 8, 665–673. [Google Scholar] [CrossRef]
- Van der Westhuizen, A.J.; Eloff, J.N. Effect of temperature and light on the toxicity and growth of the blue-green alga Microcystis aeruginosa (UV-006). Planta 1985, 163, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.F.; Oishi, S. Effects of environmental factors on toxicity of a cyanobacterium (Microcystis aeruginosa) under culture conditions. Appl. Environ. Microbiol. 1985, 49, 1342–1344. [Google Scholar] [PubMed]
- Gianuzzi, L.; Krock, B.; Crettaz Minaglia, M.C.; Rosso, L.; Houghton, C.; Sedan, D.; Malanga, G.; Espinosa, M.; Andrinolo, D.; Hernando, M. Growth, toxin production, active oxygen species and catalase activity of Microcystis aeruginosa (Cyanophyceae) exposed to temperature stress. Comp. Biochem. Physiol. Part C 2016, 189, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Rapala, J.; Sivonen, K. Assessment of environmental conditions that favor hepatotoxic and neurotoxic Anabaena spp. strains cultured under light limitation at different temperatures. Microb. Ecol. 1998, 36, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Mowe, M.A.D.; Porojan, C.; Abbas, F.; Mitrovic, S.M.; Lim, R.P.; Furey, A.; Yeo, D.C.J. Rising temperatures may increase growth rates and microcystin production in tropical Microcystis species. Harmful Algae 2015, 50, 88–98. [Google Scholar] [CrossRef]
- Tonk, L.; Welker, M.; Huisman, J.; Visser, P.M. Production of cyanopeptolins, anabaenopeptins, and microcystins by the harmful cyanobacteria Anabaena 90 and Microcystis PCC 7806. Harmful Algae 2009, 8, 219–224. [Google Scholar] [CrossRef]
- Lürling, M.; van Oosterhout, F.; Faassen, E.J. Eutrophication and warming boost cyanobacterial biomass and microcystins. Toxins 2017, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Laplante, B.; Meisner, C.; Wheeler, D.; Yan, J. The Impact of Sea Level Rise on Developing Countries: A Comparative Analysis. Clim. Chang. 2007, 93, 379–388. [Google Scholar] [CrossRef]
- MRC. Adaptation to Climate Change in the Countries of the Lower Mekong Basin: Regional Synthesis Report; MRC Technical Paper No. 24; Mekong River Commission: Vientiane, Laos, 2009; p. 89. [Google Scholar]
- Toan, P.V.; Sebesvari, Z.; Bläsing, M.; Rosendahl, I.; Renaud, F.G. Pesticide management and their residues in sediments and surface and drinking water in the Mekong Delta, Vietnam. Sci. Total Environ. 2013, 452–453, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Wilbers, G.-J.; Becker, M.; Nga, L.T.; Sebesvari, Z.; Renaud, F.G. Spatial and temporal variability of surface water pollution in the Mekong Delta, Vietnam. Sci. Total Environ. 2014, 485–486, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Hummert, C.; Dahlmann, J.; Reinhardt, K.; Dang, H.P.H.; Dang, D.K.; Luckas, B. Liquid chromatography-mass spectrometry identification of microcystins in Microcystis aeruginosa strain from lake Thanh Cong, Hanoi, Vietnam. Chromatographia 2001, 54, 569–575. [Google Scholar] [CrossRef]
- Dao, T.S.; Cronberg, G.; Nimptsch, J.; Do-Hong, L.-C.; Wiegand, C. Toxic cyanobacteria from Tri An Reservoir, Vietnam. Nova Hedwig. 2010, 90, 433–448. [Google Scholar] [CrossRef]
- Duong, T.; Le, T.; Dao, T.-S.; Pflugmacher, S.; Rochelle-Newall, E.; Hoang, T.; Vu, T.; Ho, C.; Dang, D. Seasonal variation of cyanobacteria and microcystins in the Nui Coc Reservoir, Northern Vietnam. J. Appl. Phycol. 2013, 25, 1065–1075. [Google Scholar] [CrossRef]
- Pham, T.-L.; Dao, T.-S.; Tran, N.-D.; Nimptsch, J.; Wiegand, C.; Motoo, U. Influence of environmental factors on cyanobacterial biomass and microcystin concentration in the Dau Tieng Reservoir, a tropical eutrophic water body in Vietnam. Ann. Limnol. Int. J. Limnol. 2017, 53, 89–100. [Google Scholar] [CrossRef]
- NOAA National Centers for Environmental Information. State of the Climate: Global Climate Report for April 2016. Published Online May 2016, Retrieved on 21 November 2017. Available online: https://www.ncdc.noaa.gov/sotc/global/201604 (accessed on 14 December 2017).
- Wetzel, R.G. Limnology: Lake and River Ecosystems, 3rd ed.; Academic Press: San Diego, CA, USA, 2001; pp. 213–241. [Google Scholar]
- Thomas, M.K.; Litchman, E. Effects of temperature and nitrogen availability on the growth of invasive and native cyanobacteria. Hydrobiologia 2016, 763, 357–369. [Google Scholar] [CrossRef]
- Geada, P.; Pereira, R.N.; Vasconcelos, V.; Vicente, A.A.; Fernandes, B.D. Assessment of synergistic interactions between environmental factors on Microcystis aeruginosa growth and microcystin production. Algal Res. 2017, 27, 235–243. [Google Scholar] [CrossRef]
- Eastham, J.; Mpelasoka, F.; Mainuddin, M.; Ticehurst, C.; Dyce, P.; Hodgson, G.; Ali, R.; Kirby, M. Mekong River Basin Water Resources Assessment: Impacts of Climate Change. In Water for a Healthy Country National Research Flagship Report; Commonwealth Scientific and Industrial Research Organisation (CSIRO): Canberra, Australia, 2008. [Google Scholar]
- Crettaz Minaglia, M.C.; Rosso, L.; Aranda, J.O.; Goñi, S.; Sedan, D.; Andrinolo, D.; Giannuzzi, L. Mathematical modeling of Microcystis aeruginosa growth and [D-Leu1] microcystin-LR production in culture media at different temperatures. Harmful Algae 2017, 67, 13–25. [Google Scholar]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of Microcystis under oxidative stress conditions. PLoS ONE 2011, 6, e17615. [Google Scholar] [CrossRef] [PubMed]
- Meissner, S.; Fastner, J.; Dittmann, E. Microcystin production revisited: Conjugate formation makes a major contribution. Environ. Microbiol. 2013, 15, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Altaner, S.; Puddick, J.; Wood, S.A.; Dietrich, D.R. Adsorption of ten microcystin congeners to common laboratory-ware is solvent and surface dependent. Toxins 2017, 9, 129. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Faassen, E.J. Dog poisonings associated with a Microcystis aeruginosa bloom in the Netherlands. Toxins 2013, 5, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.L.; Loftin, K.A.; Meyer, M.T.; Ziegler, A.C. Cyanotoxin mixtures and taste-and-odor compounds in cyanobacterial blooms from the Midwestern United States. Environ. Sci. Technol. 2010, 44, 7361–7368. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Faassen, E.J. Controlling toxic cyanobacteria: Effects of dredging and phosphorus-binding clay on cyanobacteria and microcystins. Water Res. 2012, 46, 1447–1459. [Google Scholar] [CrossRef] [PubMed]
- Gehringer, M.M.; Adler, L.; Roberts, A.A.; Moffitt, M.C.; Mihali, T.K.; Mills, T.J.T.; Fieker, C.; Neilan, B.A. Nodularin, a cyanobacterial toxin, is synthesized in planta by symbiotic Nostoc sp. ISME J. 2012, 6, 1834–1847. [Google Scholar] [CrossRef] [PubMed]
- Jokela, J.; Heinilä, L.M.P.; Shishido, T.K.; Wahlsten, M.; Fewer, D.P.; Fiore, M.F.; Wang, H.; Haapaniemi, E.; Permi, P.; Sivonen, K. Production of high amounts of hepatotoxin nodularin and new protease inhibitors pseudospumigins by the Brazilian benthic Nostoc sp. CENA543. Front. Microbiol. 2017, 8, 1963. [Google Scholar] [CrossRef] [PubMed]
- McGregor, G.B.; Sendall, B.C. Iningainema pulvinus gen. nov., sp. nov. (Cyanobacteria, Scytonemataceae) a new nodularin producer from Edgbaston Reserve, north-eastern Australia. Harmful Algae 2017, 62, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Van de Waal, D.B.; Ferreruela, G.; Tonk, L.; Van Donk, E.; Huisman, J.; Visser, P.M.; Matthijs, H.C.P. Pulsed nitrogen supply induces dynamic changes in the amino acid composition and microcystin production of the harmful cyanobacterium Planktothrix agardhii. FEMS Microbiol. Ecol. 2010, 74, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.M.; Smith, A.J. Nitrogen chlorosis in blue-green algae. Arch. Mikrobiol. 1969, 69, 114–120. [Google Scholar] [CrossRef]
- Nalewajko, C.; Murphy, T.P. Effects of temperature, and availability of nitrogen and phosphorus on the abundance of Anabaena and Microcystis in Lake Biwa, Japan: An experimental approach. Limnology 2001, 2, 45–48. [Google Scholar] [CrossRef]
- Rapala, J.; Sivonen, K.; Lyra, C.; Niemela, S.I. Variation of microcystin, cyanobacterial hepatotoxins, in Anabaena spp. as a function of growth stimulation. Appl. Environ. Microbiol. 1997, 63, 2206–2212. [Google Scholar] [PubMed]
- Maliaka, V.; Faassen, E.J.; Smolders, A.J.P.; Lürling, M. The impact of warming and nutrients on algae production and microcystins in seston from the iconic Lake Lesser Prespa, Greece. Toxins 2018. submitted. [Google Scholar]
- Amé, M.V.; Wunderlin, D.A. Effects of iron, ammonium and temperature on microcystin content by a natural concentrated Microcystis aeruginosa population. Water Air Soil Pollut. 2005, 168, 235–248. [Google Scholar] [CrossRef]
- Rouco, M.; López-Rodas, V.; Flores-Moya, A.; Costas, E. Evolutionary changes in growth rate and toxin production in the cyanobacterium Microcystis aeruginosa under a scenario of eutrophication and temperature increase. Microb. Ecol. 2011, 62, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Trung, B.; Dao, T.-S.; Vo, T.-G.; Faassen, E.J.; Lürling, M. Microcystins in The Mekong Delta. Toxins 2018. in preparation. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA; American Water Works Foundation: Denver, CO, USA; Water Environment Federation: Alexandria, VA, USA, 2005; p. 2671. [Google Scholar]
- Hoshaw, R.; Rosowski, J.R. Methods for microscopic algae. In Handbook of Phycological Methods. Culture Methods and Growth Measurements; Stein, R., Ed.; Cambridge University Press: London, UK, 1973; pp. 53–68. [Google Scholar]
- Lürling, M.; Beekman, W. Palmelloids formation in Chlamydomonas reinhardtii: Defence against rotifer predators? Ann. Limnol. 2006, 42, 65–72. [Google Scholar] [CrossRef]
- Faassen, E.J.; Lürling, M. Occurrence of the microcystins MC-LW and MC-LF in Dutch surface waters and their contribution to total microcystin toxicity. Mar. Drugs 2013, 11, 2643–2654. [Google Scholar] [CrossRef] [PubMed]
- Zurawell, R.W.; Chen, H.; Burke, J.M.; Prepas, E.E. Hepatotoxic cyanobacteria: A review of the biological importance of microcystins in freshwater environments. J. Toxicol. Environ. Health Part B 2005, 8, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Hoeger, S.J.; Stemmer, K.; Feurstein, D.J.; Knobeloch, D.; Nussler, A.; Dietrich, D.R. The role of organic anion transporting polypeptides (OATPs/SLCOs) in the toxicity of different microcystin congeners in vitro: A comparison of primary human hepatocytes and OATP-transfected HEK293 cells. Toxicol. Appl. Pharmacol. 2010, 245, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Chang, K.-H.; Kusaba, M.; Nakano, S. Temperature-dependent dominance of Microcystis (Cyanophyceae) species: M. aeruginosa and M. wesenbergii. J. Plankton Res. 2009, 31, 171–178. [Google Scholar] [CrossRef]
- Sivonen, K. Effects of light, temperature, nitrate, orthophosphate, and bacteria on growth of and hepatotoxin production by Oscillatoria agardhii strains. Appl. Environ. Microbiol. 1990, 56, 2658–2666. [Google Scholar] [PubMed]
- Staehr, P.A.; Birkeland, M.J. Temperature acclimation of growth, photosynthesis and respiration in tow mesophilic phytoplankton species. Phycologia 2006, 45, 648–656. [Google Scholar] [CrossRef]
- Nicklisch, A.; Kohl, J.-G. Growth kinetics of Microcystis aeruginosa (Kütz.) Kütz as a basis for modelling its population dynamics. Int. Rev. Gesamten Hydrobiol. 1983, 68, 317–326. [Google Scholar] [CrossRef]
- Ohkubo, N.; Yagi, O.; Okada, M. Effects of temperature and illumination on the growth of blue-green alga Microcystis viridis. Jpn. J. Limnol. 1991, 52, 255–261. [Google Scholar] [CrossRef]
- Lürling, M.; Eshetu, F.; Faassen, E.J.; Kosten, S.; Huszar, V.L.M. Comparison of cyanobacterial and green algal growth rates at different temperatures. Freshwater Biol. 2013, 58, 552–559. [Google Scholar] [CrossRef]
- Gomes, A.; Azevedo, S.M.O.; Lürling, M. Temperature effect on exploitation and interference competition among Microcystis aeruginosa, Planktothrix agardhii and, Cyclotella meneghiniana. Sci. World J. 2015, 2015, 834197. [Google Scholar] [CrossRef]
- NNI. Water—Spectrophotometric Determination of Chlorophyll-a Content. NEN 6520+ C1; Netherlands Normalization Institute: Delft, The Netherlands, 2011. (In Dutch) [Google Scholar]
- Moed, J.A.; Hallegraeff, G.M. Some problems in the estimation of chlorophyll-a and phaeopigments from pre- and post-acidification spectrophotometric measurements. Int. Rev. Gesamten Hydrobiol. 1978, 63, 787–800. [Google Scholar] [CrossRef]





| Strain | Temp. (°C) | Salinity (‰) | pH | TN (mg/L) | TP (mg/L) | N-NH4 (mg/L) | N-NO3 (mg/L) | P-PO4 (mg/L) | CHLa (µg/L) |
|---|---|---|---|---|---|---|---|---|---|
| MBC1 | 37.5 | 7.4 | 9.89 | 9.13 | 0.25 | 0.57 | BDL | BDL | 1480 |
| MBC2 | 37.4 | 7.5 | 9.77 | 9.00 | 0.33 | 0.67 | BDL | BDL | 1520 |
| MBC3 | 37.4 | 7.4 | 9.79 | 9.36 | 0.30 | 0.46 | BDL | BDL | 1437 |
| MBC4 | 37.7 | 7.6 | 9.66 | 16.91 | 1.52 | 1.02 | 0.04 | 0.19 | 5100 |
| MDT | 33.5 | 0.0 | 9.82 | 3.98 | 0.59 | 0.39 | 0.14 | 0.10 | 169 |
| MTV | 33.1 | 0.5 | 9.58 | 19.5 | 1.75 | 1.14 | 0.28 | BDL | 4352 |
| BDL, below detection limit | |||||||||
| Strain | Chlorophyll-a Based Growth Rates | |||
| 27 °C | 31 °C | 35 °C | 37 °C | |
| MBC1 | 0.35 (0.01)A | 0.42 (0.03)B | 0.42 (0.03)B | 0.41 (0.02)B |
| MBC2 | 0.32 (0.02)A | 0.37 (0.03)B | 0.36 (0.03)B | 0.32 (0.02)A |
| MBC3 | 0.35 (0.02)A | 0.42 (0.02)B | 0.42 (0.02)B | 0.43 (0.02)B |
| MBC4 | 0.36 (0.03)A | 0.44 (0.03)B | 0.45 (0.02)B | 0.43 (0.03)B |
| MDT | 0.24 (0.01)A | 0.40 (0.02)B | 0.30 (0.01)C | --- |
| MTV | 0.34 (0.04)A | 0.35 (0.02)A | 0.36 (0.03)A | 0.38 (0.03)A |
| Strain | Particle Concentration Based Growth Rates | |||
| 27 °C | 31 °C | 35 °C | 37 °C | |
| MBC1 | 0.28 (0.03)A | 0.33 (0.04)B | 0.41 (0.04)C | 0.35 (0.02)B |
| MBC2 | 0.33 (0.02)A | 0.37 (0.04)A | 0.34 (0.03)A | 0.35 (0.02)A |
| MBC3 | 0.33 (0.03)A | 0.34 (0.04)A | 0.42 (0.03)B | 0.39 (0.02)B |
| MBC4 | 0.35 (0.06)AB | 0.33 (0.04)A | 0.40 (0.03)C | 0.39 (0.06)BC |
| MDT | 0.18 (0.03)A | 0.37 (0.01)B | 0.27 (0.01)C | --- |
| MTV | 0.31 (0.04)AB | 0.31 (0.03)A | 0.44 (0.04)C | 0.25 (0.06)B |
| Strain | Biovolume Concentration Based Growth Rates | |||
| 27 °C | 31 °C | 35 °C | 37 °C | |
| MBC1 | 0.31 (0.04)A | 0.34 (0.06)A | 0.44 (0.08)B | 0.38 (0.05)AB |
| MBC2 | 0.34 (0.06)A | 0.45 (0.11)AB | 0.54 (0.09)B | 0.19 (0.03)A |
| MBC3 | 0.41 (0.06)A | 0.42 (0.07)A | 0.46 (0.04)A | 0.23 (0.04)B |
| MBC4 | 0.25 (0.05)A | 0.29 (0.04)A | 0.38 (0.04)B | 0.45 (0.06)C |
| MDT | 0.39 (0.03)AB | 0.41 (0.05)A | 0.30 (0.02)B | --- |
| MTV | 0.27 (0.07)A | 0.38 (0.07)A | 0.36 (0.07)A | 0.40 (0.06)A |
| Temp. | Microcystis Strains | |||||
|---|---|---|---|---|---|---|
| MBC1 | MBC2 | MBC3 | MBC4 | MDT | MTV # | |
| 27 °C | A | A | A | A | A | A |
| 31 °C | A | B | A | A | A | AB |
| 35 °C | B | C | B | B | B | AB |
| 37 °C | B | D | B | B | C | B |
| F3,11 value | 17.6 | 94.4 | 14.7 | 24.4 | 149.1 | H3 = 9.5 |
| p-value | p < 0.001 | p < 0.001 | p = 0.001 | p < 0.001 | p < 0.001 | p = 0.024 |
| Normality | p = 0.555 | p = 0.627 | p = 0.420 | p = 0.374 | p = 0.114 | p = 0.439 |
| Equal Var. | p = 0.634 | p = 0.323 | p = 0.863 | p = 0.307 | p = 0.455 | p < 0.050 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bui, T.; Dao, T.-S.; Vo, T.-G.; Lürling, M. Warming Affects Growth Rates and Microcystin Production in Tropical Bloom-Forming Microcystis Strains. Toxins 2018, 10, 123. https://doi.org/10.3390/toxins10030123
Bui T, Dao T-S, Vo T-G, Lürling M. Warming Affects Growth Rates and Microcystin Production in Tropical Bloom-Forming Microcystis Strains. Toxins. 2018; 10(3):123. https://doi.org/10.3390/toxins10030123
Chicago/Turabian StyleBui, Trung, Thanh-Son Dao, Truong-Giang Vo, and Miquel Lürling. 2018. "Warming Affects Growth Rates and Microcystin Production in Tropical Bloom-Forming Microcystis Strains" Toxins 10, no. 3: 123. https://doi.org/10.3390/toxins10030123
APA StyleBui, T., Dao, T.-S., Vo, T.-G., & Lürling, M. (2018). Warming Affects Growth Rates and Microcystin Production in Tropical Bloom-Forming Microcystis Strains. Toxins, 10(3), 123. https://doi.org/10.3390/toxins10030123