A Single Tri-Epitopic Antibody Virtually Recapitulates the Potency of a Combination of Three Monoclonal Antibodies in Neutralization of Botulinum Neurotoxin Serotype A


 , ,
, ,  and
and 
Abstract
1. Introduction
2. Results and Discussion
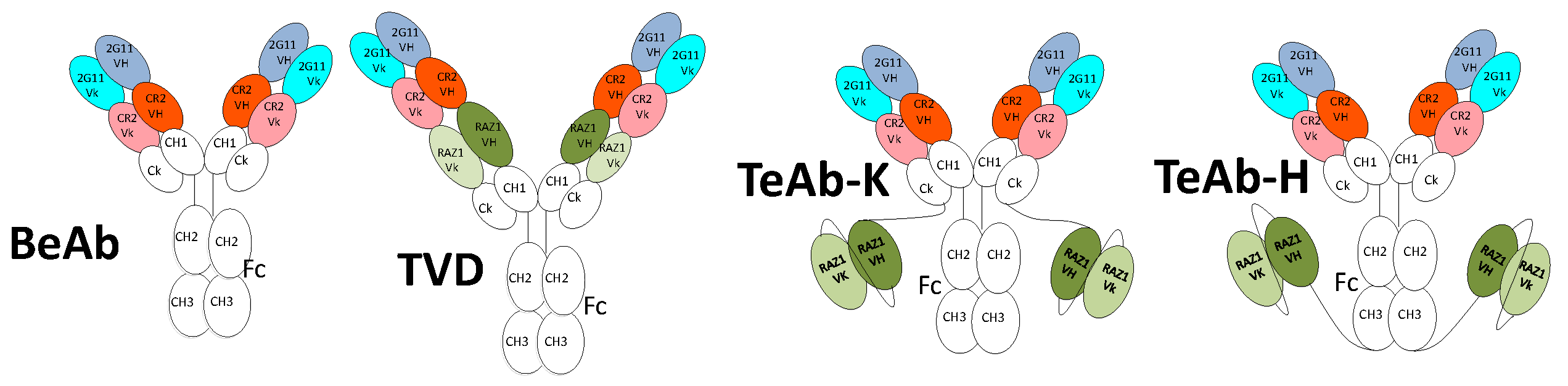
2.1. Design of Bi-Epitopic (BeAb) and Tri-Epitopic Antibodies (TeAb)
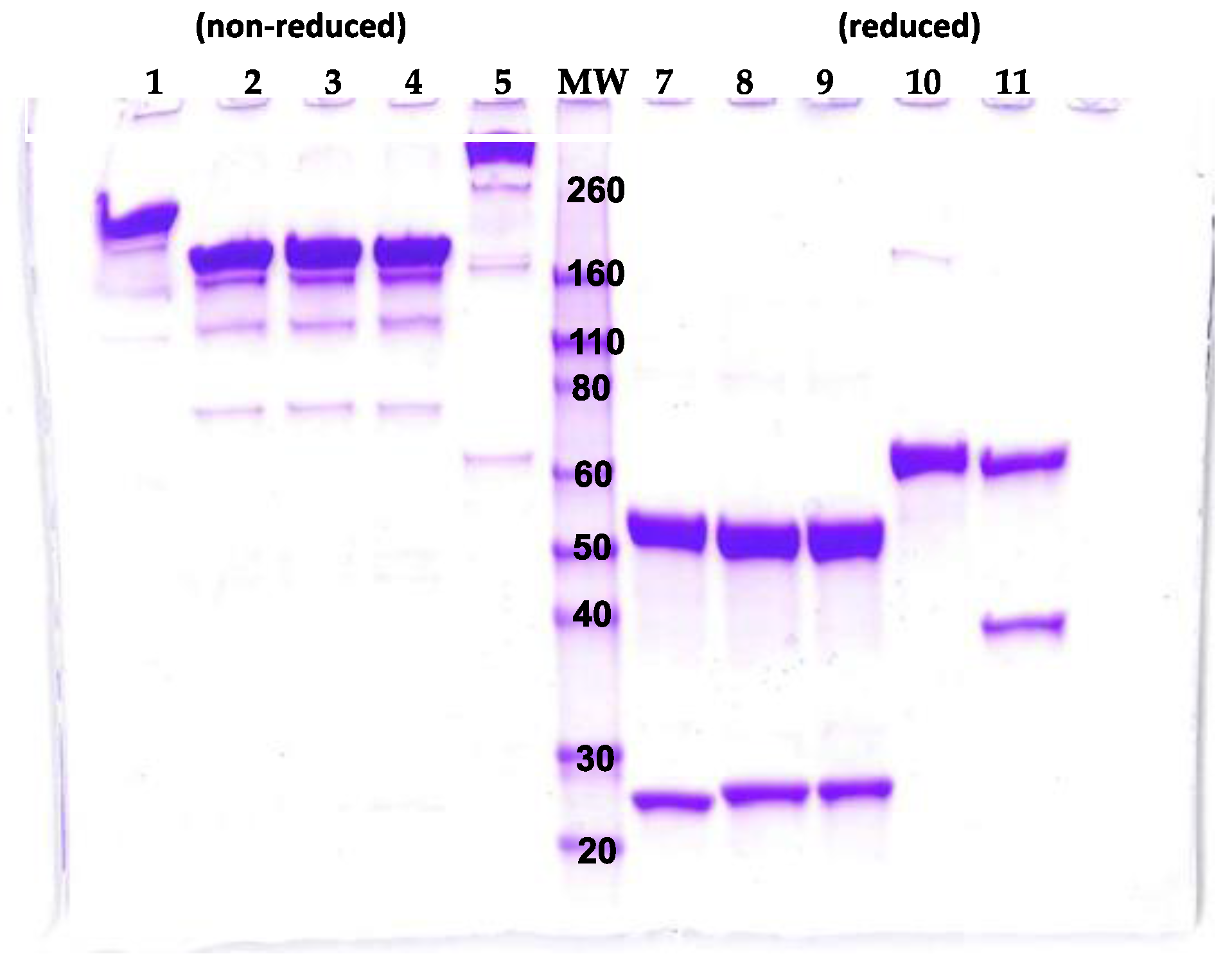
2.2. Initial Characterization of the BeAb
2.2.1. Affinity of the BeAb for Botulinum Neurotoxin (BoNT/A)
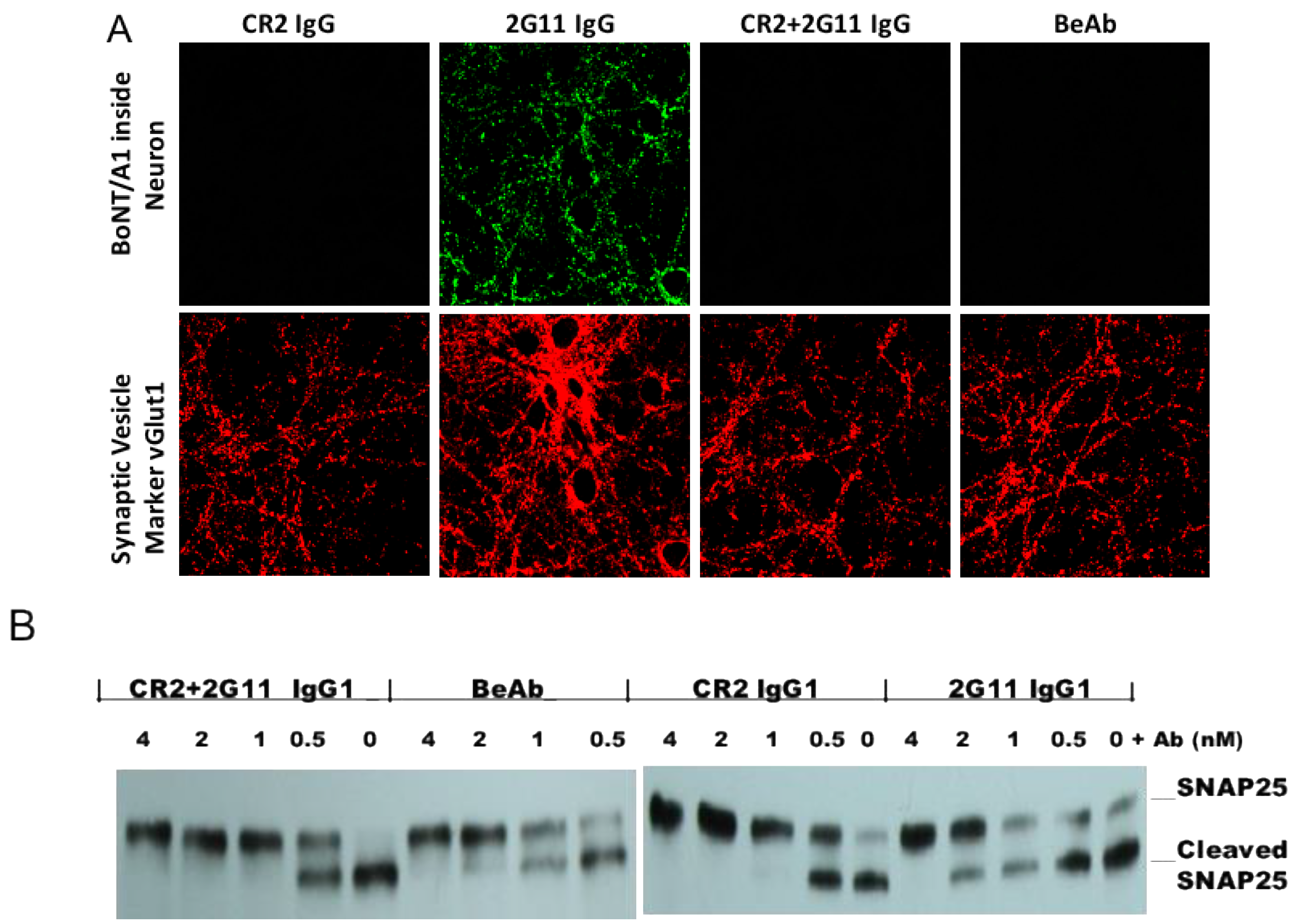
2.2.2. In Vitro Functional Effects of the BeAb Compared to 2G11 and CR2 IgG
2.3. Initial Characterization of the TeAb
Affinity of the TeAb for BoNT/A
2.4. BeAb and TeAb-K Neutralization of BoNT/A In Vivo
2.5. Mechanism of BeAb and TeAb BoNT/A Neutralization
3. Materials and Methods
3.1. Design, Cloning, Expression and Purification of BeAb and TeAb
- BglIIinDraIIIoutVk5′primer: (5′-ATCACAGATCTCTCACCATGAGGGTCCCCGCTCAGCTCCTGGGGCTCCTGCTGCTCTGGCTCCCAGGTGCCCGATG-3′)
- DraIIIinBsiWoutVklinker3′primer: (5′-ACTGCTCATCACATCGTGAGAAGATGAAGACAGATGGTGCAGCCACGGTACGTTTGATTTCC-3′)
- SalIinMluIoutVH5′primer: (5′-GACCCGTCGACATGGGTTGGAGCCTCATCTTGCTCTTCCTTGTCGCTGTTGCTACCCGTGTCTTGTCCC-3′)
- MluIinNheIout VH3′ tail primer: (5′-GGAGGAGGGTGCCAGACGCGTGACCGATGGGCCCTTGGTGCTTGCTGAGGAGACGG-3′)
3.2. Binding Affinity Determinations Using KinExA Analysis
3.3. Inhibition of BoNT/A1 Cellular Uptake and SNAP25 Substrate Cleavage in Primary Neurons by IgG1 or BeAb
3.4. Mouse Neutralization Assay of BoNT/A1 by IgG, BeAb or TeAb
3.5. Estimation of TeAb-K Complex Size and Composition Using Negative-Stain Electron Microscopy (NS-EM)
3.6. Pharmacokinetic Studies of ciBoNT/A in the Presence of Antibodies
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hatheway, C.H.; Snyder, J.D.; Seals, J.E.; Edell, T.A.; Lewis, G.E., Jr. Antitoxin levels in botulism patients treated with trivalent equine botulism antitoxin to toxin types A, B, and E. J. Infect. Dis. 1984, 150, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Hibbs, R.G.; Weber, J.T.; Corwin, A.; Allos, B.M.; El Rehim, M.S.A.; El Sharkawy, S.; Sarn, J.E.; McKee, K.T., Jr. Experience with the use of an investigational F (ab′) 2 heptavalent botulism immune globulin of equine origin during an outbreak of type E botulism in Egypt. Clin. Infect. Dis. 1996, 23, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Arnon, S.S.; Schechter, R.; Maslanka, S.E.; Jewell, N.P.; Hatheway, C.L. Human botulism immune globulin for the treatment of infant botulism. N. Engl. J. Med. 2006, 354, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Frankovich, T.L.; Arnon, S.S. Clinical trial of botulism immune globulin for infant botulism. West J. Med. 1991, 154, 103. [Google Scholar] [PubMed]
- Cangene Corp., BAT® [Botulism Antitoxin Heptavalent (A, B, C, D, E, F, G)—(Equine)]. Sterile Solution for Injection. Available online: https://www.fda.gov/downloads/.../UCM345147.pdf (accessed on 1 October 2017).
- Fagan, R.P.; Neil, K.P.; Sasich, R.; Luquez, C.; Asaad, H.; Maslanka, S.; Khalil, W. Initial recovery and rebound of type f intestinal colonization botulism after administration of investigational heptavalent botulinum antitoxin. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2011, 53, e125–e128. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, A.; Wang, C.; Powers, D.B.; Amersdorfer, P.; Smith, T.J.; Montgomery, V.A.; Sheridan, R.; Blake, R.; Smith, L.A.; Marks, J.D. Potent neutralization of botulinum neurotoxin by recombinant oligoclonal antibody. Proc. Natl. Acad. Sci. USA 2002, 99, 11346–11350. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Barash, J.R.; Lou, J.; Conrad, F.; Marks, J.D.; Arnon, S.S. Immunological Characterization and Neutralizing Ability of Monoclonal Antibodies Directed Against Botulinum Neurotoxin Type H. J. Infect. Dis. 2016, 213, 1606–1614. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Garcia-Rodriguez, C.; Lou, J.; Wen, W.; Conrad, F.; Zhai, W.; Smith, T.J.; Smith, L.A.; Marks, J.D. A three monoclonal antibody combination potently neutralizes multiple botulinum neurotoxin serotype F subtypes. PLoS ONE 2017, 12, e0174187. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.; Griffiss, J.; McKenzie, R.; Fuchs, E.; Jurao, R.; An, A.; Ahene, A.; Tomic, M.; Hendrix, C.; Zenilman, J. Safety and Pharmacokinetics of XOMA 3AB, a Novel Mixture of Three Monoclonal Antibodies against Botulinum Toxin A. Antimicrob. Agents Chemother. 2014, 58, 5047–5053. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Ying, H.; Grinnell, C.; Bryant, S.; Miller, R.; Clabbers, A.; Bose, S.; McCarthy, D.; Zhu, R.R.; Santora, L.; et al. Simultaneous targeting of multiple disease mediators by a dual-variable-domain immunoglobulin. Nat. Biotechnol. 2007, 25, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- Lou, J.; Geren, I.; Garcia-Rodriguez, C.; Forsyth, C.M.; Wen, W.; Knopp, K.; Brown, J.; Smith, T.; Smith, L.A.; Marks, J.D. Affinity maturation of human botulinum neurotoxin antibodies by light chain shuffling via yeast mating. Protein Eng. Des. Sel. 2010, 23, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Razai, A.; Garcia-Rodriguez, C.; Lou, J.; Geren, I.; Forsyth, C.M.; Robles, Y.; Tsai, R.; Smith, T.J.; Amith, L.A.; Siegel, R.W.; et al. Molecular evolution of antibody affinity for sensitive detection of botulinum neurotoxin type A. J. Mol. Biol. 2005, 351, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Li, M.; Silberg, M.A.; Conrad, F.; Bettencourt, J.; To, R.; Huang, C.; Ma, J.; Meyer, K.; Shimizu, R.; et al. Domain-based assays of individual antibody concentrations in an oligoclonal combination targeting a single protein. Anal. Biochem. 2012, 421, 351–361. [Google Scholar] [CrossRef] [PubMed]
- DiGiammarino, E.L.; Harlan, J.E.; Walter, K.A.; Ladror, U.S.; Edalji, R.P.; Hutchins, C.W.; Lake, M.R.; Greischar, A.J.; Liu, J.; Ghayur, T. Ligand Association Rates to the Inner-Variable-Domain of a Dual-Variable-Domain Immunoglobulin are Significantly Impacted by Linker Design. MAbs 2011, 3, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Strotmeier, J.; Mahrhold, S.; Krez, N.; Janzen, C.; Lou, J.; Marks, J.D.; Binz, T.; Rummel, A. Identification of the synaptic vesicle glycoprotein 2 receptor binding site in botulinum neurotoxin A. FEBS Lett. 2014, 588, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.; Terraube, V.; Tam, A.S.P.; Stochaj, W.; Fennell, B.J.; Lin, L.; Stahl, M.; LaVallie, E.R.; Somers, W.; Finlay, W.J. A Combination of Structural and Empirical Analyses Delineates the Key Contacts Mediating Stability and Affinity Increases in an Optimized Biotherapeutic Single-chain Fv (scFv). J. Biol. Chem. 2016, 291, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Solomon, H.M.; Lilly, T., Jr. Bacteriological Analytical Manual (BAM): Clostridium botulinum. Available online: https://www.fda.gov/Food/FoodScienceResearch/LaboratoryMethods/ucm070879.htm (accessed on 25 November 2017).
- Al-Saleem, F.H.; Ancharski, D.M.; Ravichandran, E.; Joshi, S.G.; Singh, A.K.; Gong, Y.; Simpson, L.L. The role of systemic handling in the pathophysiologic actions of botulinum toxin. J. Pharmacol. Exp. Ther. 2008, 326, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Webb, R.P.; Smith, T.J.; Wright, P.; Brown, J.; Smith, L.A. Production of catalytically inactive BoNT/A1 holoprotein and comparison with BoNT/A1 subunit vaccines against toxin subtypes A1, A2, and A3. Vaccine 2009, 27, 4490–4497. [Google Scholar] [CrossRef] [PubMed]
- Lacy, D.B.; Tepp, W.; Cohen, A.C.; DasGupta, B.R.; Stevens, R.C. Crystal structure of botulinum neurotoxin type A and implications for toxicity. Nat. Struct. Biol. 1998, 5, 898–902. [Google Scholar] [CrossRef] [PubMed]
- O’Hear, C.E.; Foote, J. Antibody buffering of a ligand in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Mannik, M.; Haakenstad, A.O.; Arend, W.P. The fate and detection of circulating immune complexes. In Progress in Immunology, II; Brent, L., Holborow, J., Eds.; North-Holland: Amsterdam, The Netherlands, 1974; p. 91. [Google Scholar]
- Montero-Julian, F.A.; Klein, B.; Gautherot, E.; Brailly, H. Pharmacokinetic study of anti-interleukin-6 (IL-6) therapy with monoclonal antibodies: Enhancement of IL-6 clearance by cocktails of anti-IL-6 antibodies. Blood 1995, 85, 917–924. [Google Scholar] [PubMed]
- Garcia-Rodriguez, C.; Levy, R.; Arndt, J.W.; Forsyth, C.M.; Razai, A.; Lou, J.; Geren, I.; Stevens, R.C.; Marks, J.D. Molecular evolution of antibody cross-reactivity for two subtypes of type A botulinum neurotoxin. Nat. Biotechnol. 2007, 25, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rodriguez, C.; Geren, I.; Lou, J.; Conrad, F.; Forsyth, C.; Wen, W.; Chakraborti, S.; Zao, H.; Manzanarez, G.; Smith, T. Neutralizing human monoclonal antibodies binding multiple serotypes of botulinum neurotoxin. Protein Eng. Des. Sel. 2011, 24, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Blake, R.C., II; Pavlov, A.R.; Blake, D.A. Automated kinetic exclusion assays to quantify protein binding interactions in homogeneous solution. Anal. Biochem. 1999, 272, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Kaech, S.; Banker, G. Culturing hippocampal neurons. Nat. Protoc. 2006, 1, 2406–2415. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Geren, I.N.; Dong, J.; Lou, J.; Wen, W.; Conrad, F.; Smith, T.J.; Smith, L.A.; Ho, M.; Pires-Alves, M.; et al. Monoclonal Antibodies Targeting the Alpha-Exosite of Botulinum Neurotoxin Serotype/A Inhibit Catalytic Activity. PLoS ONE 2015, 10, e0135306. [Google Scholar] [CrossRef] [PubMed]
- Ohi, M.; Li, Y.; Cheng, Y.; Walz, T. Negative Staining and Image Classification—Powerful Tools in Modern Electron Microscopy. Biol. Proced. Online 2004, 6, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.W.; Onisko, B.; Johnson, E.A.; Reader, J.R.; Griffey, S.M.; Larson, A.E.; Tepp, W.H.; Stanker, L.H.; Brandon, D.L.; Carter, J.M. Effects of purification on the bioavailability of botulinum neurotoxin type A. Toxicology 2008, 249, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Correia, I.; Sung, J.; Burton, R.; Jakob, C.G.; Carragher, B.; Ghayur, T.; Radziejewski, C. The Structure of Dual-Variable-Domain Immunoglobulin Molecules Alone and Bound to Antigen. MAbs 2013, 5, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Adekar, S.P.; Takahashi, T.; Jones, R.M.; Al-Saleem, F.H.; Ancharski, D.M.; Root, M.J.; Kapadnis, B.P.; Simpson, L.L.; Dessain, S.K. Neutralization of botulinum neurotoxin by a human monoclonal antibody specific for the catalytic light chain. PLoS ONE 2008, 3, e3023. [Google Scholar] [CrossRef] [PubMed]
- Dimasi, N.; Gao, C.; Fleming, R.; Woods, R.M.; Yao, X.T.; Shirinian, L.; Kiener, P.A.; Wu, H. The design and characterization of oligospecific antibodies for simultaneous targeting of multiple disease mediators. J. Mol. Biol. 2009, 393, 672–692. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J.E.; Dufour, P. Adecatumumab: An anti-EpCAM monoclonal antibody, from the bench to the bedside. Expert Opin. Biol. Ther. 2010, 10, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Torisu-Itakura, H.; Schoellhammer, H.F.; Sim, M.S.; Irie, R.F.; Hausmann, S.; Raum, T.; Baeuerle, P.A.; Morton, D.L. Redirected Lysis of Human Melanoma Cells by a MCSP/CD3-bispecific BiTE Antibody That Engages Patient-derived T Cells. J. Immunother. 2011, 34, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Vugmeyster, Y.; Zhang, Y.E.; Zhong, X.; Wright, J.; Leung, S.S. Pharmacokinetics of anti-IL17A and anti-IL22 peptide–antibody bispecific genetic fusions in mice. Int. Immunopharmacol. 2014, 18, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Alt, M.; Müller, R.; Kontermann, R.E. Novel tetravalent and bispecific IgG-like antibody molecules combining single-chain diabodies with the immunoglobulin γ1 Fc or CH3 region. FEBS Lett. 1999, 454, 90–94. [Google Scholar] [CrossRef]
- Castoldi, R.; Jucknischke, U.; Pradel, L.P.; Arnold, E.; Klein, C.; Scheiblich, S.; Niederfellner, G.; Sustmann, C. Molecular characterization of novel trispecific ErbB-cMet-IGF1R antibodies and their antigen-binding properties. Protein Eng. Des. Sel. 2012, 25, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Schoonjans, R.; Willems, A.; Schoonooghe, S.; Fiers, W.; Grooten, J.; Mertens, N. Fab chains as an efficient heterodimerization scaffold for the production of recombinant bispecific and trispecific antibody derivatives. J. Immunol. 2000, 165, 7050–7057. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-B.; Zhao, B.-F.; Zhao, Q.; Piao, J.-H.; Liu, J.; Lin, Q.; Huang, H.-L. A new recombinant single chain trispecific antibody recruits T lymphocytes to kill CEA (carcinoma embryonic antigen) positive tumor cells in vitro efficiently. J. Biochem. 2004, 135, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.M.; Vakis, S.A.; Ayre, K.R.; Ellwood, C.N.; Howell, W.M.; Tutt, A.L.; Cawley, M.I.; Smith, J.L. Rheumatoid arthritis T cells produce Th1 cytokines in response to stimulation with a novel trispecific antibody directed against CD2, CD3, and CD28. Scand. J. Rheumatol. 2000, 29, 282–287. [Google Scholar] [PubMed]
- Somasundaram, C.; Sundarapandiyan, K.; Keler, T.; Deo, Y.M.; Graziano, R.F. Development of a trispecific antibody conjugate that directs two distinct tumor-associated antigens to CD64 on myeloid effector cells. Hum. Antib. 1999, 9, 47–54. [Google Scholar]
- Mertens, N.; Schoonjans, R.; Willems, A.; Schoonooghe, S.; Leoen, J.; Grooten, J. New Recombinant Bi-and Trispecific Antibody Derivatives. Novel Front. Prod. Compd. Biomed. Use 2001, 1, 195–208. [Google Scholar]
- Xu, L.; Pegu, A.; Rao, E.; Doria-Rose, N.; Beninga, J.; McKee, K.; Lord, D.M.; Wei, R.R.; Deng, G.; Louder, M.; et al. Trispecific broadly neutralizing HIV antibodies mediate potent SHIV protection in macaques. Science 2017, 358, 85–90. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BoNT/A1 Domain | BoNT/A1 Holotoxin | |||||
|---|---|---|---|---|---|---|
| mAb | KD (pM) | kon (106 M−1s−1) | koff (10−6 s−1) | KD (pM) | kon (106 M−1 s−1) | koff (10−6 s−1) c |
| HCN (CR2 epitope) | ||||||
| CR2 | 263.49 (325.94–171.09) a | 4.681 (6.081–3.514) a | 1234 | 8.84 (13.39–5.31) | 3.650 (2.988–4.417) | 32.27 |
| BeAb b | 10,040 (12,760–6790) | 0.3986 (0.4957–0.3463) | 4002 | 4.73 (6.31–3.44) | 0.7907 (0.8688–0.7181) | 3.740 |
| TeAb-K | 22,620 (27,550–15,540) | 0.1563 (0.1716–0.1429) | 3536 | 2.57 (4.60–1.02) | 1.138 (1.290–0.992) | 2.924 |
| LC-HN (2G11 epitope) | ||||||
| 2G11 | 11.11 (13.75–8.46) | 0.6317 (0.7363–0.5383) | 7.018 | 11.41 (15.28–8.23) | 0.5027 (5.514–4.559) | 5.736 |
| BeAb | 21.13 (25.37–16.23) | 0.3937 (.4378–.3525) | 8.320 | 12.78 (21.52–6.62) | 2.346 (2.927–1.842) | 29.98 |
| TeAb-K | 25.79 (29.75–22.21) | 0.2616 (0.2739–0.2508) | 6.747 | 2.59 (5.78–0.57) | 2.128 (2.779–1.615) | 5.512 |
| HCC (RAZ1 epitope) | ||||||
| RAZ1 | 1.33 (1.92–0.813) | 10.02 (14.61–6.222) | 13.33 | 1.73 (1.96–1.53) | 15.89 (16.84–15.03) | 27.49 |
| TeAb-K | 25.53 (45.03–12.23) | 2.115 (2.331–1.919) | 53.99 | 15.18 (25.62–8.10) | 1.186 (1.284–1.092) | 18.00 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lou, J.; Wen, W.; Conrad, F.; Meng, Q.; Dong, J.; Sun, Z.; Garcia-Rodriguez, C.; Farr-Jones, S.; Cheng, L.W.; Henderson, T.D.; et al. A Single Tri-Epitopic Antibody Virtually Recapitulates the Potency of a Combination of Three Monoclonal Antibodies in Neutralization of Botulinum Neurotoxin Serotype A. Toxins 2018, 10, 84. https://doi.org/10.3390/toxins10020084
Lou J, Wen W, Conrad F, Meng Q, Dong J, Sun Z, Garcia-Rodriguez C, Farr-Jones S, Cheng LW, Henderson TD, et al. A Single Tri-Epitopic Antibody Virtually Recapitulates the Potency of a Combination of Three Monoclonal Antibodies in Neutralization of Botulinum Neurotoxin Serotype A. Toxins. 2018; 10(2):84. https://doi.org/10.3390/toxins10020084
Chicago/Turabian StyleLou, Jianlong, Weihua Wen, Fraser Conrad, Qi Meng, Jianbo Dong, Zhengda Sun, Consuelo Garcia-Rodriguez, Shauna Farr-Jones, Luisa W. Cheng, Thomas D. Henderson, and et al. 2018. "A Single Tri-Epitopic Antibody Virtually Recapitulates the Potency of a Combination of Three Monoclonal Antibodies in Neutralization of Botulinum Neurotoxin Serotype A" Toxins 10, no. 2: 84. https://doi.org/10.3390/toxins10020084
APA StyleLou, J., Wen, W., Conrad, F., Meng, Q., Dong, J., Sun, Z., Garcia-Rodriguez, C., Farr-Jones, S., Cheng, L. W., Henderson, T. D., Brown, J. L., Smith, T. J., Smith, L. A., Cormier, A., & Marks, J. D. (2018). A Single Tri-Epitopic Antibody Virtually Recapitulates the Potency of a Combination of Three Monoclonal Antibodies in Neutralization of Botulinum Neurotoxin Serotype A. Toxins, 10(2), 84. https://doi.org/10.3390/toxins10020084