Mitigating Toxic Planktonic Cyanobacterial Blooms in Aquatic Ecosystems Facing Increasing Anthropogenic and Climatic Pressures


Abstract
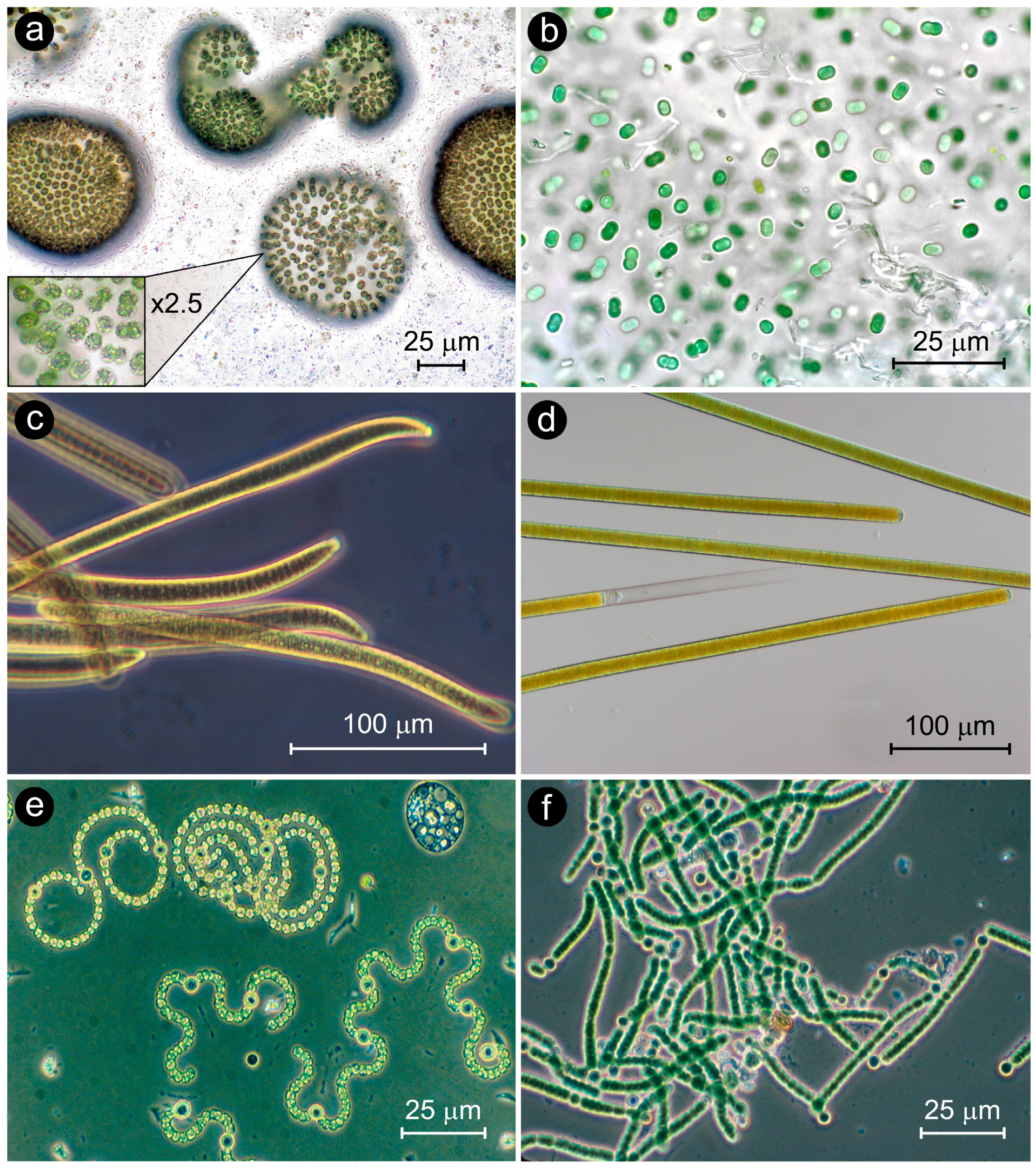
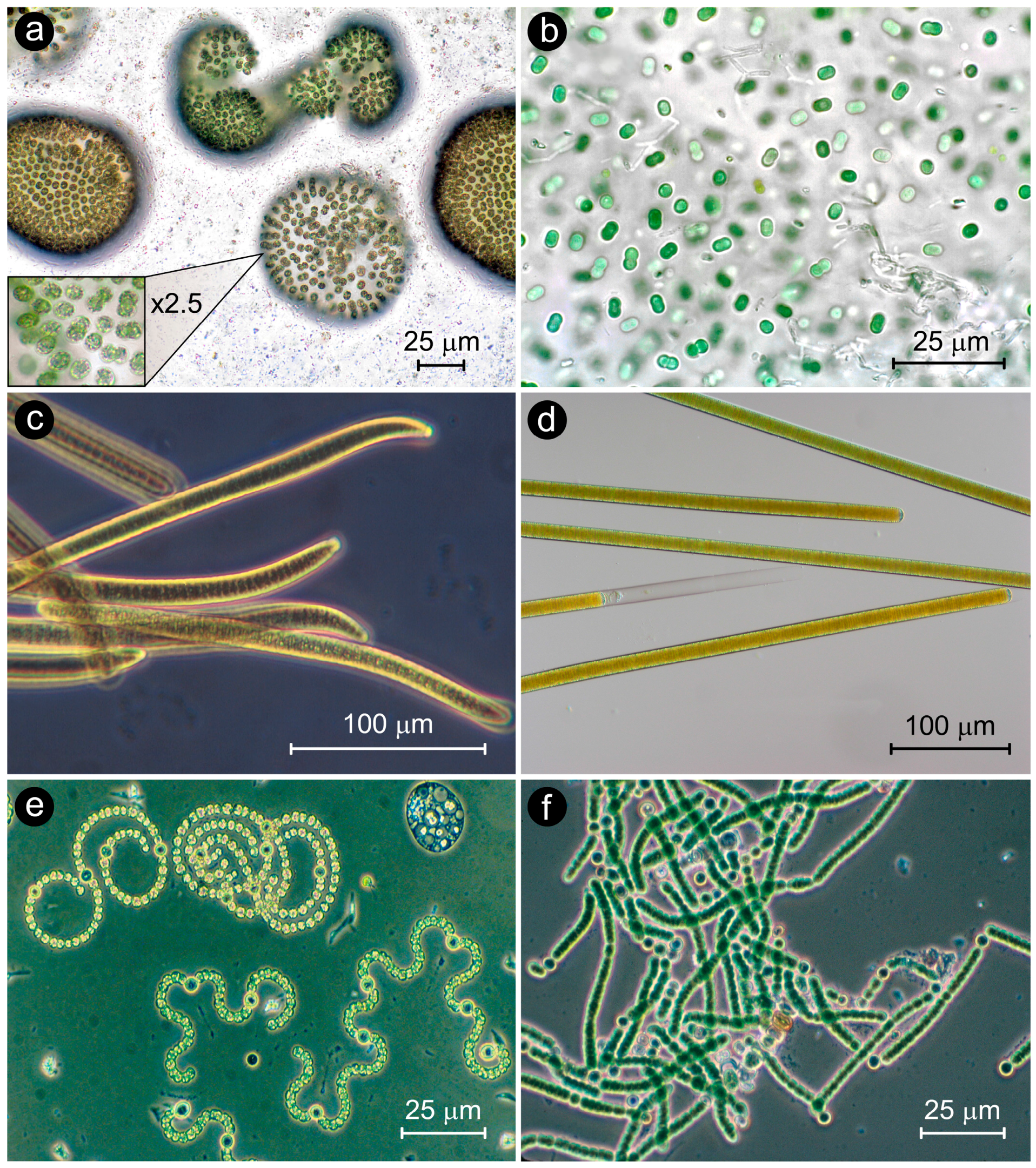
:1. Introduction
2. CyanoHAB Mitigation Strategies
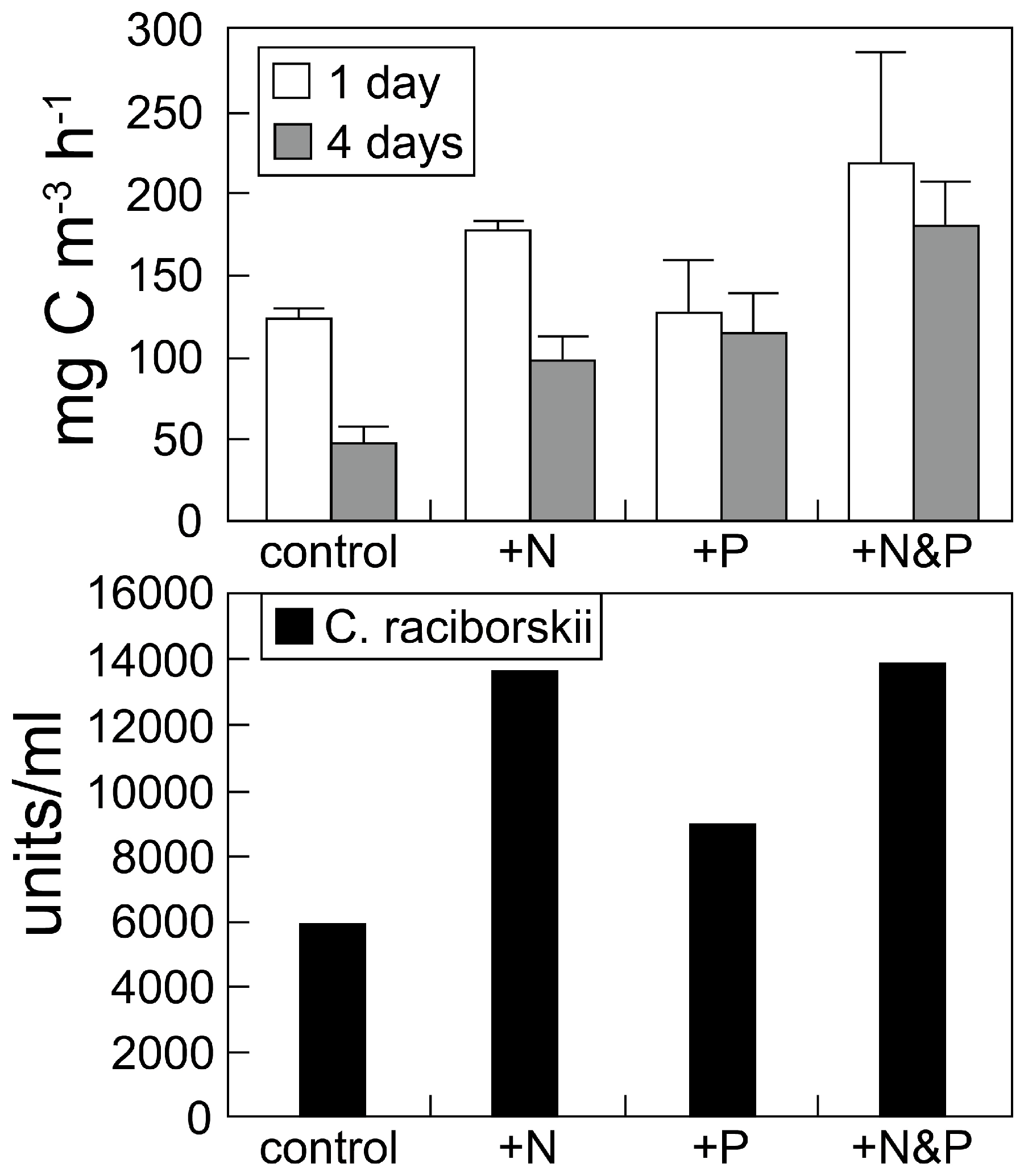
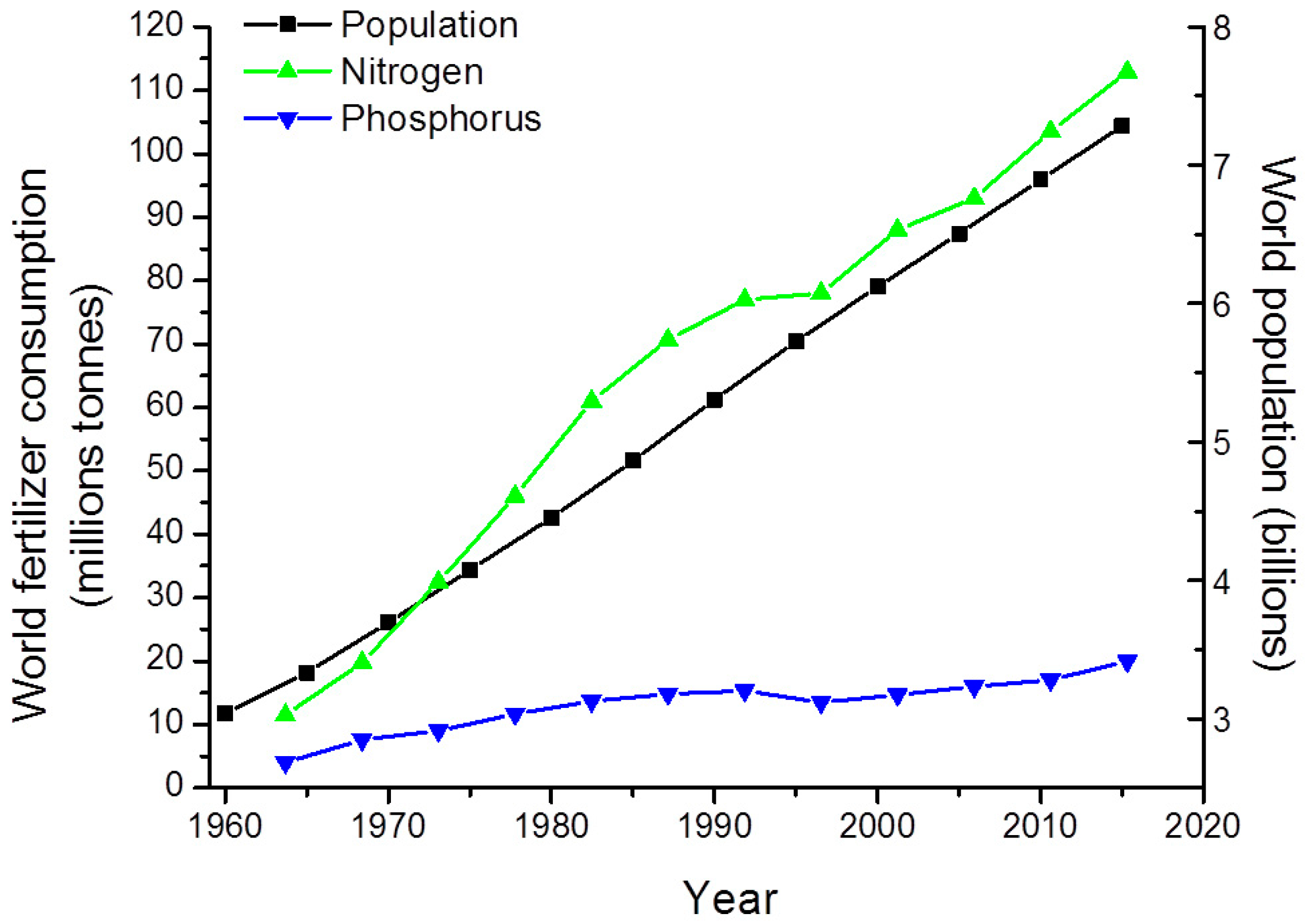
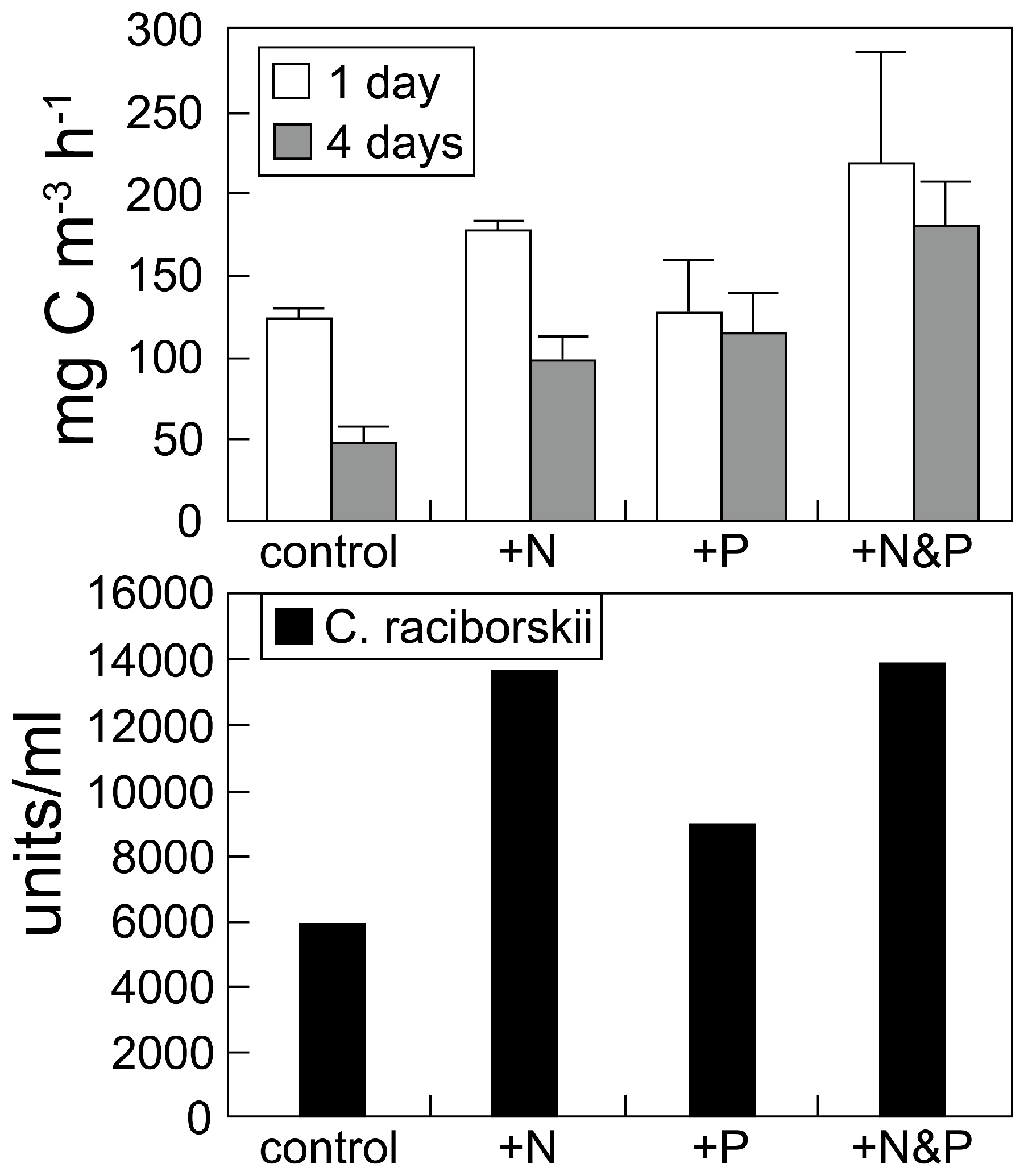
2.1. Nutrient-Based Mitigation Strategies
2.2. Altering Sediment Nutrient Dynamics
2.3. Hydrologic Manipulations
2.4. Application of Algaecides and Biomanipulation
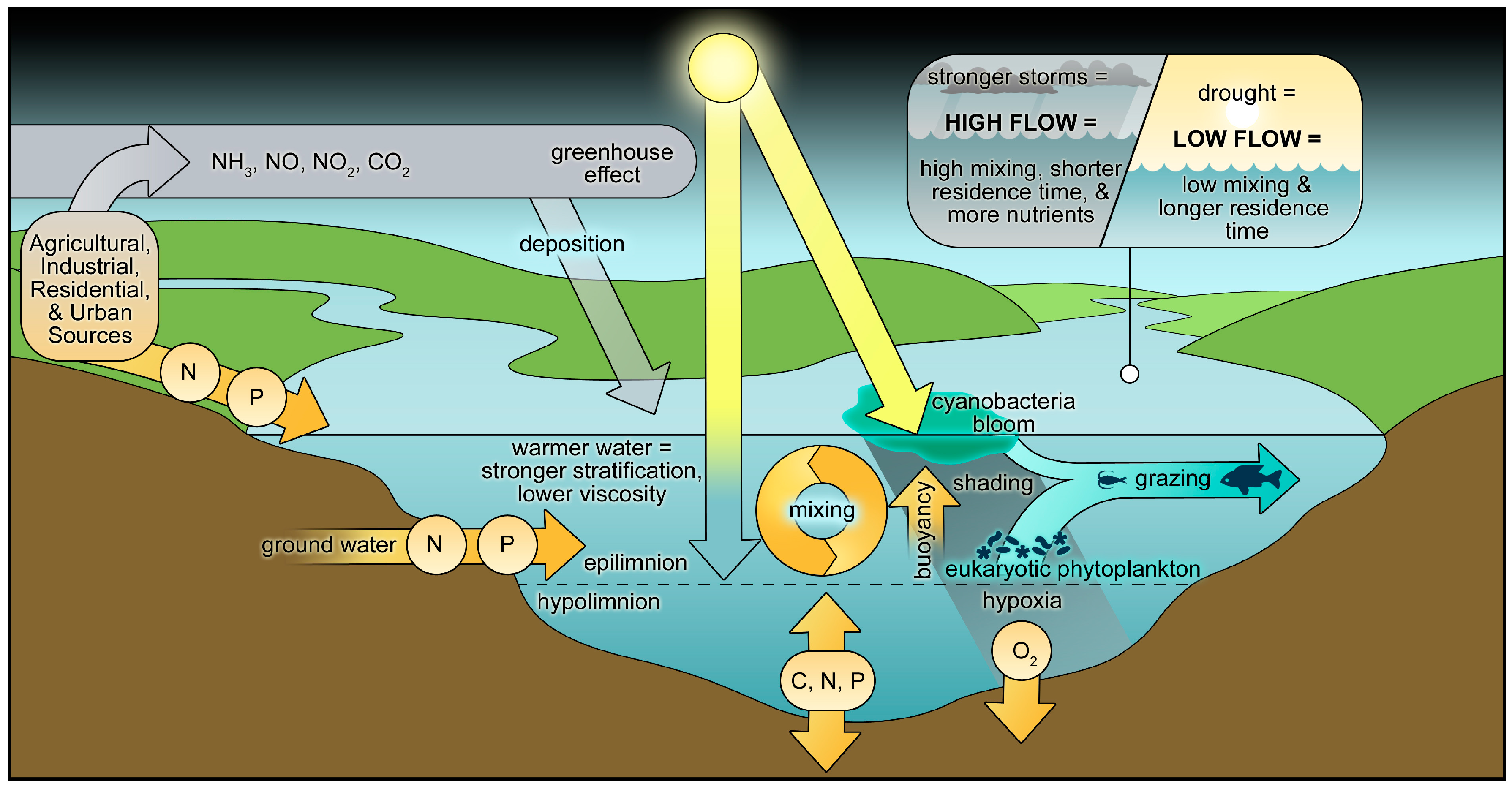
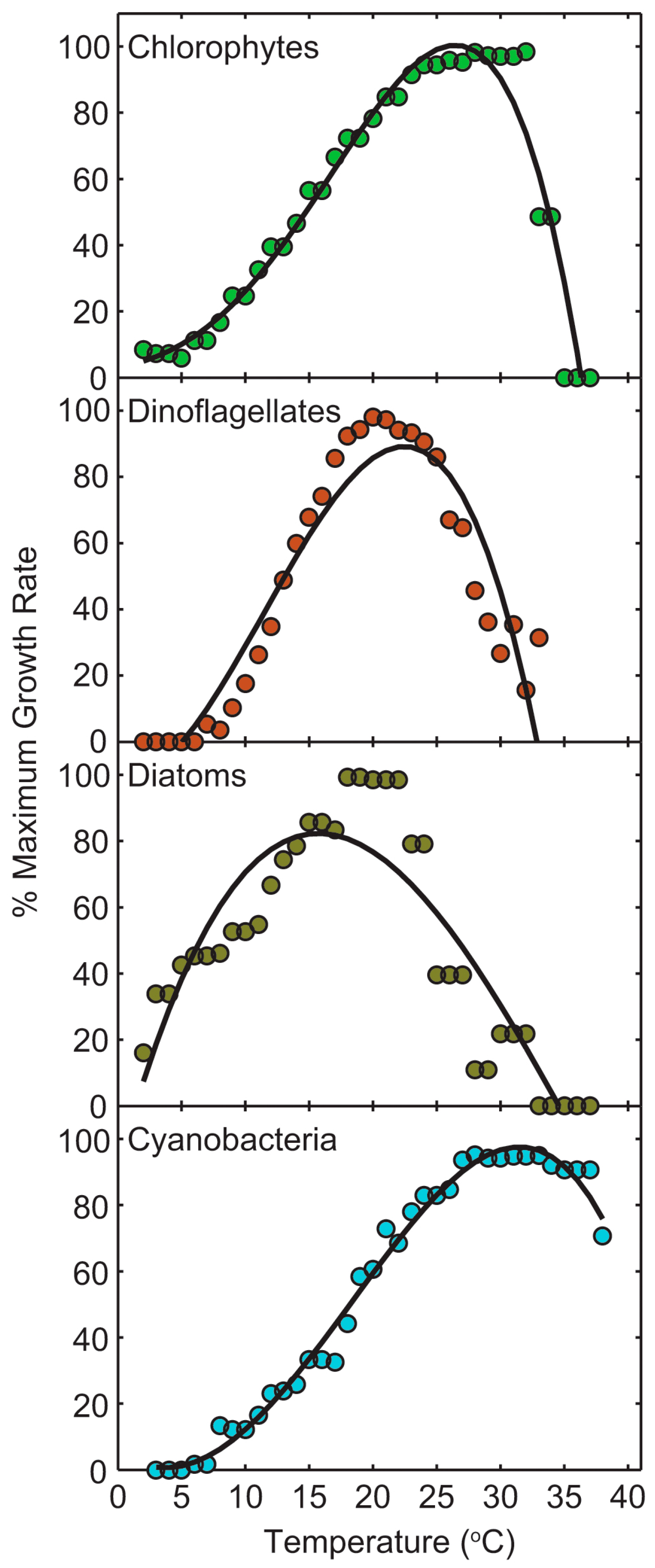
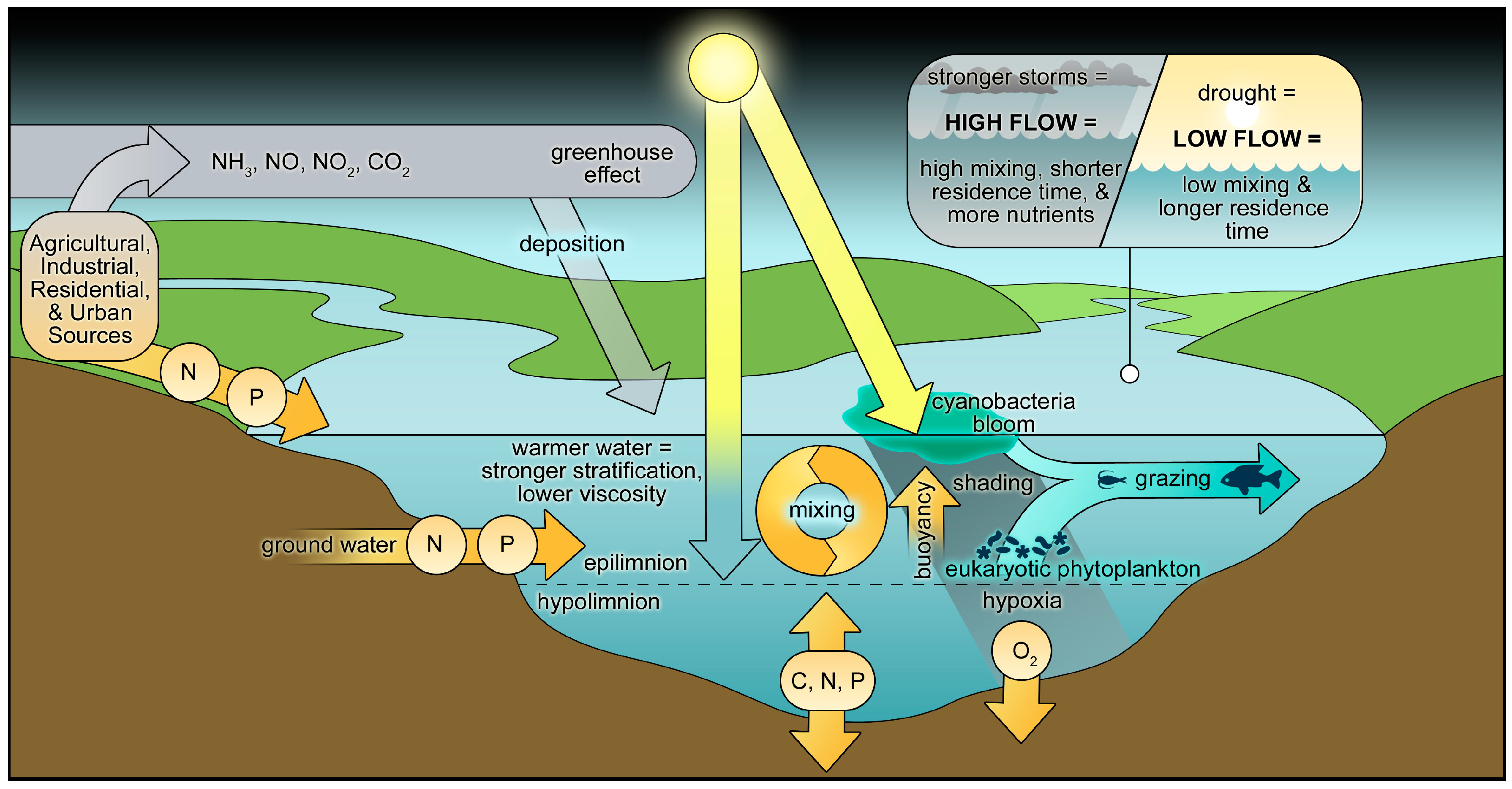
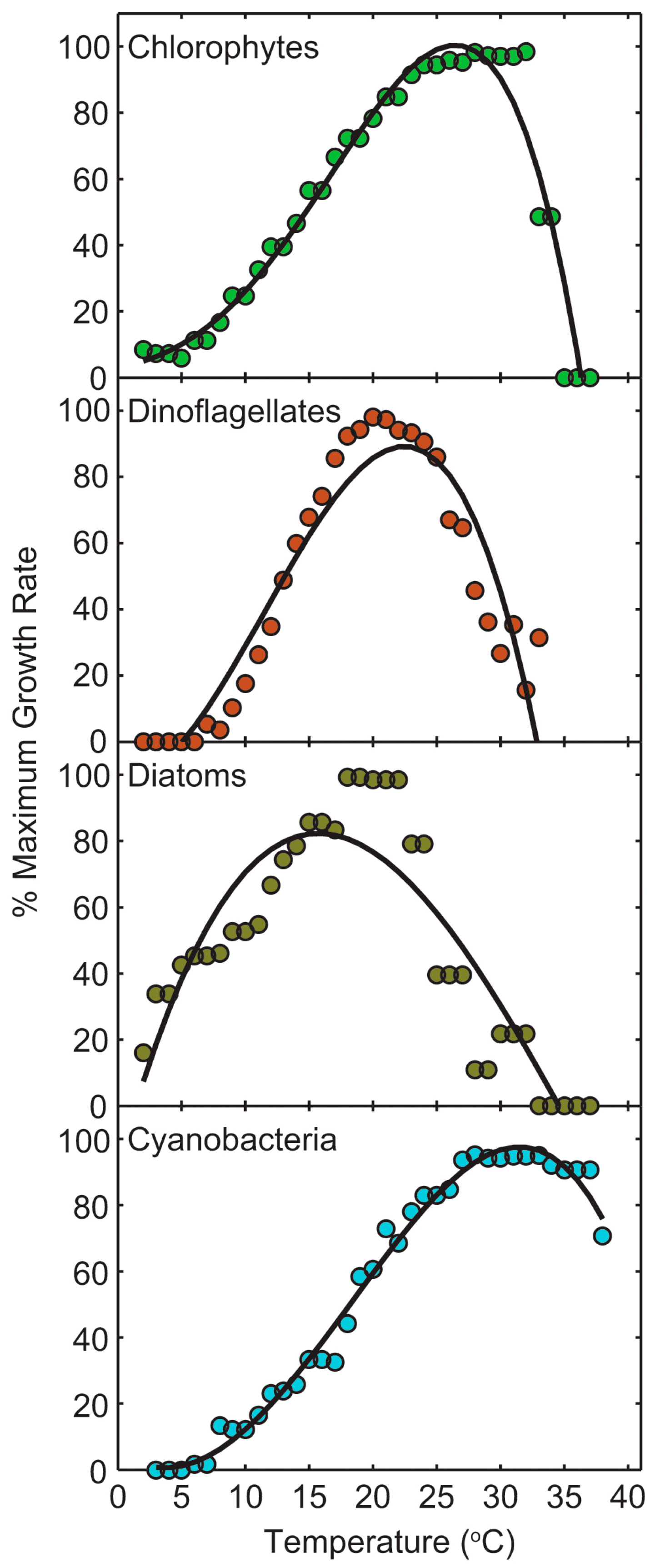
2.5. Climate Change and CyanoHAB Potentials
3. Conclusions
Acknowledgments
Conflicts of Interest
References
- Schopf, J.W. The Fossil Record: Tracing the Roots of the Cyanobacterial Lineage. In The Ecology of Cyanobacteria; Whitton, B.A., Potts, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 13–35. [Google Scholar]
- Knoll, A.H. Cyanobacteria and Earth History. In The Cyanobacteria, Molecular Biology, Genetics and Evolution; Herrero, A., Flores, E., Eds.; Caister Academic Press: Norfolk, UK, 2008; pp. 1–20. [Google Scholar]
- Potts, M.; Whitton, B.A. The Biology and Ecology of Cyanobacteria, 2nd ed.; Blackwell Scientific Publications: Oxford, UK, 2000; p. 669. [Google Scholar]
- Reynolds, C.S. Ecology of Phytoplankton (Ecology, Biodiversity and Conservation); Cambridge University Press: Cambridge, UK, 2006; p. 524. [Google Scholar]
- Gallon, J.R. Tansley Review No. 44/Reconciling the incompatible: N2 fixation and O2. New Phytol. 1992, 122, 571–609. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Trick, C.G. Iron-limited growth of cyanobacteria: Multiple siderophore production is a common response. Limnol. Oceanogr. 1994, 39, 1979–1984. [Google Scholar] [CrossRef]
- Paerl, H.W.; Millie, D.F. Physiological ecology of toxic cyanobacteria. Phycologia 1996, 35, 160–167. [Google Scholar] [CrossRef]
- Huisman, J.M.; Matthijs, H.C.P.; Visser, P.M. Harmful Cyanobacteria. Springer Aquatic Ecology Series, 3rd ed.; Springer: Dordrecht, The Netherlands, 2005; p. 243. [Google Scholar]
- Paerl, H.W.; Otten, T.G. Harmful Cyanobacterial Blooms: Causes, Consequences and Controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Blooms bite the hand that feeds them. Science 2013, 342, 433–434. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W. Interactions with Bacteria. In The Biology of Cyanobacteria; Carr, N.G., Whitton, B.A., Eds.; Blackwell Scientific Publications Ltd.: Oxford, UK, 1982; pp. 441–461. [Google Scholar]
- Paerl, H.W. Growth and Reproductive Strategies of Freshwater Blue-green Algae (Cyanobacteria). In Growth and Reproductive Strategies of Freshwater Phytoplankton; Sandgren, C.D., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 363–400. [Google Scholar]
- Paerl, H.W. Nuisance phytoplankton blooms in coastal, estuarine, and inland waters. Limnol. Oceanogr. 1988, 33, 823–847. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci. Total Environ. 2011, 409, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Gardner, W.S.; Havens, K.E.; Joyner, A.R.; McCarthy, M.J.; Newell, S.E.; Qin, B.; Scott, J.T. Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 2016, 54, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Trenberth, K.E. The Impact of Climate Change and Variability on Heavy Precipitation, Floods, and Droughts. In Encyclopedia of Hydrological Sciences; Anderson, M.G., Ed.; John Wiley and Sons, Ltd.: Hoboken, NJ, USA, 2005; pp. 1–11. [Google Scholar]
- Webster, P.J.; Holland, G.J.; Curry, J.A.; Chang, H.R. Changes in tropical cyclone number, duration, and intensity in a warming environment. Science 2005, 309, 1844–1846. [Google Scholar] [CrossRef] [PubMed]
- Holland, G.J.; Webster, P.J. Heightened tropical cyclone activity in the North Atlantic: Natural variability of climate trend? Philos. Trans. R. Soc. A 2007. [Google Scholar] [CrossRef] [PubMed]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L.A., Eds.; Intergovernment Panel on Climate Change (IPCC): Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Bgai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- Sterner, R.W. On the phosphorus limitation paradigm for lakes. Int. Rev. Hydrobiol. 2008, 93, 433–445. [Google Scholar] [CrossRef]
- Lewis, W.M.; Wurtsbaugh, W.A.; Paerl, H.W. Rationale for control of anthropogenic nitrogen and phosphorus in inland waters. Environ. Sci. Technol. 2011, 45, 10030–10035. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Fulton, R.S., III; Moisander, P.H.; Dyble, J. Harmful freshwater algal blooms, with an emphasis on cyanobacteria. The Sci. World 2001, 1, 76–113. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.; Qin, B.; Li, Y.; Gardner, W.S. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P) management strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar]
- Rabalais, N.N. Nitrogen in aquatic ecosystems. Ambio 2002, 31, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Simpson, T.W.; Sharpley, A.N.; Howarth, R.W.; Paerl, H.W.; Mankin, K.R. The new gold rush: Fueling ethanol production while protecting water quality. J. Environ. Qual. 2008, 37, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Howarth, R.W.; Swaney, D.P.; Billen, G.; Garnier, J.; Hong, B.; Humborg, C.; Johnes, P.; Mörth, C.M.; Marino, R.M. Nitrogen fluxes from large watersheds to coastal ecosystems controlled by net anthropogenic nitrogen inputs and climate. Front. Ecol. Environ. 2012, 10, 37–43. [Google Scholar] [CrossRef]
- Likens, G.E. Nutrients and Eutrophication: The Limiting Nutrient Controversy. In Nutrients and Eutrophication: The Limiting Nutrient Controversy; Likens, G.E., Ed.; American Society of Limnology and Oceanography: Lawrence, KS, USA, 1972; p. 328. [Google Scholar]
- Paerl, H.W. Coastal eutrophication and harmful algal blooms: Importance of atmospheric deposition and groundwater as “new” nitrogen and other nutrient sources. Limnol. Oceanogr. 1997, 42, 1154–1165. [Google Scholar] [CrossRef]
- Galloway, J.N.; Cowling, E.B.; Seitzinger, S.P.; Sokolow, R.H. Reactive nitrogen: Too much of a good thing. aMbio 2002, 31, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Andersen, T.; Baron, J.S.; Bergstrom, A.K.; Jansson, M.; Kyle, M.; Nydick, K.R.; Steger, L.; Hessen, D.O. Shifts in lake N:P stoichiometry and nutrient limitation driven by atmospheric nitrogen deposition. Science 2009, 326, 835–837. [Google Scholar] [CrossRef] [PubMed]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling eutrophication: Nitrogen and phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Scott, J.T.; McCarthy, M.J.; Newell, S.E.; Gardner, W.S.; Havens, K.E.; Hoffman, D.K.; Wilhelm, S.W.; Wurtsbaugh, W.A. It takes two to tango: When and where dual nutrient (N & P) reductions are needed to protect lakes and downstream ecosystems. Environ. Sci. Technol. 2016, 50, 10805–10813. [Google Scholar]
- Davis, T.W.; Berry, D.L.; Boyer, G.L.; Gobler, C.J. The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae 2009, 8, 715–725. [Google Scholar] [CrossRef]
- Seitzinger, S.P.; Harrison, J.A.; Boehlke, J.K.; Bouwman, A.F.; Lowrance, R.; Peterson, B.; Tobias, C.; van Drecht, G. Denitrification across landscapes and waterscapes: A synthesis. Ecol. Appl. 2006, 16, 2064–2090. [Google Scholar] [CrossRef]
- Grantz, E.M.; Kogo, A.; Scott, J.T. Partitioning whole-lake denitrification using in situ dinitrogen gas accumulation and intact sediment core experiments. Limnol. Oceanogr. 2012, 57, 925–935. [Google Scholar] [CrossRef]
- Scott, J.T.; McCarthy, M.J. Nitrogen fixation may not balance the nitrogen pool in lakes over timescales relevant to eutrophication management. Limnol. Oceangr. 2010, 55, 1265–1270. [Google Scholar] [CrossRef]
- Paerl, H.W.; Scott, J.T. Throwing fuel on the fire: Synergistic effects of excessive nitrogen inputs and global warming on harmful algal blooms. Environ. Sci. Technol. 2010, 44, 7756–7758. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Paul, V. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2011, 46, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.W. The dilemma of controlling cultural eutrophication of lakes. Proc. R. Soc. B 2012. [Google Scholar] [CrossRef] [PubMed]
- Downing, J.A.; Watson, S.B.; McCauley, E. Predicting cyanobacteria dominance in lakes. Can. J. Fish. Aquat. Sci. 2001, 58, 1905–1908. [Google Scholar] [CrossRef]
- Paerl, H.W. Combating the global proliferation of harmful cyanobacterial blooms by integrating conceptual and technological advances in an accessible water management toolbox. Environ. Microbiol. Rep. 2013, 5, 12–14. [Google Scholar]
- Scott, J.T.; McCarthy, M.J.; Otten, T.G.; Steffen, M.M.; Baker, B.C.; Grantz, E.M.; Wilhelm, S.W.; Paerl, H.W. Comment: An alternative interpretation of the relationship between TN:TP and microcystins in Canadian lakes. Can. J. Fish. Aquat. Sci. 2013, 70, 1–4. [Google Scholar] [CrossRef]
- Smith, V.H. Low nitrogen to phosphorus ratios favor dominance by blue-green algae in Lake Phytoplankton. Science 1983, 221, 669–671. [Google Scholar] [CrossRef] [PubMed]
- U.S. Environmental Protection Agency. Reactive Nitrogen in the United States: An Analysis of Inputs, Flows, Consequences, and Management Options; EPA Scientific Advisory Board: Washington, DC, USA, August 2011.
- Hamilton, D.P.; Salmaso, N.; Paerl, H.W. Mitigating harmful cyanobacterial blooms: Strategies for control of nitrogen and phosphorus loads. Aquat. Ecol. 2016, 50, 351–366. [Google Scholar] [CrossRef]
- Sharpley, A.N.; Daniel, T.; Sims, T.; Lemunyon, T.J.; Stevens, R.; Parry, R. Agricultural Phosphorus and Eutrophication, 2nd ed.; USDA-ARS Pasture Systems & Watershed Management Research Unit: University Park, PA, USA, 2010; p. 44.
- King, S.E.; Osmond, D.L.; Smith, L.; Dukes, M.; Evans, R.O.; Knies, S.; Kunickis, S.; Burchell, M.; Gilliam, J.W. Effects of riparian buffer vegetation and width: A 12-year retrospective study. J. Environ. Qual. 2015, 45, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Healy, F.P. Phosphate. In The Biology of Cyanobacteria, 1st ed.; Carr, N.G., Whitton, B.A., Eds.; Blackwell Scientific Publications: Oxford, UK, 1982; pp. 105–124. [Google Scholar]
- Lean, D.R.S. Movement of phosphorus between its biologically-important forms in lakewater. J. Fish. Res. Board Can. 1973, 30, 1525–1536. [Google Scholar] [CrossRef]
- Galloway, J.N.; Dentener, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitzinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen cycles: Past, present, and future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Cooke, G.D.; Welch, E.B.; Spencer, S.P.; Newroth, P.R. Lake and Reservoir Restoration, 2nd ed.; Lewis Publishers: Boca Raton, FL, USA, 1993; p. 390. [Google Scholar]
- Cronberg, G. Changes in the phytoplankton of Lake Trummen induced by restoration. Hydrobiologia 1982, 86, 185–193. [Google Scholar] [CrossRef]
- Petersen, S.A. Lake restoration by sediment removal. J. Am. Water Res. Assoc. 1982, 18, 423–435. [Google Scholar] [CrossRef]
- Robb, M.; Greenop, B.; Goss, Z.; Douglas, G.; Adeney, J. Application of Phoslock, an innovative phosphorous binding clay, to two Western Australian waterways: Preliminary findings. Hydrobiologia 2003, 494, 237–243. [Google Scholar] [CrossRef]
- Spears, B.M.; Lurling, M.; Yasseri, S.; Castro-Castellon, A.; Gibbs, M.; Meis, S.; McDonald, C.; McIntosh, J.; Sleep, D.; Van Oosterhout, F. Lake responses following lanthanum-modified bentonite clay (Phoslock®) application: An analysis of water column lanthanum data from 16 case study lakes. Water Res. 2013, 15, 5930–5942. [Google Scholar] [CrossRef] [PubMed]
- Visser, P.M.; Ibelings, B.W.; van der Veer, B.; Koedood, J.; Mur, L.R. Artificial mixing prevents nuisance blooms of the cyanobacterium Microcystis in Lake Nieuwe Meer, The Netherlands. Freshw. Biol. 1996, 36, 435–450. [Google Scholar] [CrossRef]
- Mitrovic, S.M.; Oliver, R.L.; Rees, C.; Bowling, L.C.; Buckney, R.T. Critical flow velocities for the growth and dominance of Anabaena circinalis in some turbid freshwater rivers. Freshw. Biol. 2003, 48, 164–174. [Google Scholar] [CrossRef]
- Maier, H.R.; Kingston, G.B.; Clark, T.; Frazer, A.; Sanderson, A. Risk-based approach for assessing the effectiveness of flow management in controlling cyanobacterial blooms in rivers. River Res. Appl. 2004, 20, 459–471. [Google Scholar] [CrossRef]
- Hanson, M.J.; Stefan, H.G. Side effects of 58 years of copper sulfate treatment of the Fairmont Lakes, Minnesota. J. Am. Water Res. Assoc. 1984, 20, 889–900. [Google Scholar] [CrossRef]
- Matthijs, H.C.P.; Visser, P.M.; Reeze, B.; Meeuse, J.; Slot, P.C.; Wjin, G.; Talens, R.; Huisman, J. Selective suppression of harmful cyanobacteria in an entire lake with hydrogen peroxide. Water Res. 2012, 46, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- McQueen, D.J. Manipulating lake community structure: Where do we go from here? Freshw. Biol. 1990, 23, 613–620. [Google Scholar] [CrossRef]
- Ghadouani, A.; Pinel-Alloul, B.; Prepas, E.E. Effects of experimentally induced cyanobacterial blooms on crustacean zooplankton communities. Freshw. Biol. 2003, 48, 363–381. [Google Scholar] [CrossRef]
- Moss, B.; Madgwick, J.; Phillips, J.G. A Guide to the Restoration of Nutrient-Enriched Shallow Lakes, 1st ed.; W.W. Hawes: Bury St. Edmonds, UK, 1996; p. 194. [Google Scholar]
- Scheffer, M. Ecology of Shallow Lakes, 1st ed.; Chapman and Hall: London, UK, 1998; p. 313. [Google Scholar]
- Jeppesen, E.; Meerhoff, M.; Jacobsen, B.A.; Hansen, R.S.; Søndergaard, M.; Jensen, J.P.; Lauridsen, T.L.; Mazzeo, N.; Branco, C.W.C. Restoration of shallow lakes by nutrient control and biomanipulation: The successful strategy varies with lake size and climate. Hydrobiologia 2007, 581, 269–285. [Google Scholar] [CrossRef]
- Jöhnk, K.D.; Huisman, J.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef]
- Paul, V.J. Global Warming and Cyanobacterial Harmful Algal Blooms. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Hudnell, H.K., Ed.; Springer: New York, NY, USA, 2008; pp. 239–257. [Google Scholar]
- Kosten, S.; Huszar, V.L.M.; Bécares, E.; Costa, L.S.; van Donk, E.; Hansson, L.A.; Jeppesen, E.; Kruk, C.; Lacerot, G.; Mazzeo, N.; De Meester, L.; et al. Warmer climates boost cyanobacterial dominance in shallow lakes. Glob. Chang. Biol. 2012, 18, 118–126. [Google Scholar] [CrossRef]
- Foy, R.H.; Gibson, C.E.; Smith, R.V. The influence of daylength, light intensity and temperature on the growth rates of planktonic blue-green algae. Eur. J. Phycol. 1976, 11, 151–163. [Google Scholar] [CrossRef]
- Robarts, R.D.; Zohary, T. Temperature effects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria. N. Z. J. Mar. Freshw. Res. 1987, 21, 391–399. [Google Scholar] [CrossRef]
- Butterwick, C.; Heaney, S.I.; Talling, J.F. Diversity in the influence of temperature on the growth rates of freshwater algae, and its ecological relevance. J. Freshw. Biol. 2005, 50, 291–300. [Google Scholar] [CrossRef]
- Weyhenmeyer, G.A. Warmer winters: Are planktonic algal populations in Sweden’s largest lakes affected? aMbio 2001, 30, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Elliott, J.A. The seasonal sensitivity of cyanobacteria and other phytoplankton to changes in flushing rate and water temperature. Glob. Chang. Biol. 2010, 16, 864–876. [Google Scholar] [CrossRef]
- Stüken, A.; Rücker, J.; Endrulat, T.; Preussel, K.; Hemm, M.; Nixdorf, B.; Karsten, U.; Wiedner, C. Distribution of three alien cyanobacterial species (Nostocales) in northeast Germany: Cylindrospermopsis raciborskii, Anabaena bergii and Aphanizomenon aphanizomenoides. Phycologia 2006, 45, 696–703. [Google Scholar] [CrossRef]
- Peeters, F.; Straile, D.; Lorke, A.; Livingstone, D.M. Earlier onset of the spring phytoplankton bloom in lakes of the temperate zone in a warmer climate. Glob. Chang. Biol. 2007, 13, 1898–1909. [Google Scholar] [CrossRef]
- Suikkanen, S.; Laamanen, M.; Huttunen, M. Long-term changes in summer phytoplankton communities of the open northern Baltic Sea. Estuar. Coast. Shelf Sci. 2007, 71, 580–592. [Google Scholar] [CrossRef]
- Wiedner, C.; Rücker, J.; Brüggemann, R.; Nixdorf, B. Climate change affects timing and size of populations of an invasive cyanobacterium in temperate regions. Oecologia 2007, 152, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.; Adrian, R. Cyanobacteria dominance: Quantifying the effects of climate change. Limnol. Oceanogr. 2009, 54, 2460–2468. [Google Scholar] [CrossRef]
- Allan, R.P.; Soden, B.J. Atmospheric warming and the amplification of precipitation extremes. Science 2008, 321, 1481–1484. [Google Scholar] [CrossRef] [PubMed]
- Walsby, A.E.; Hayes, P.K.; Boje, R.; Stal, L.J. The selective advantage of buoyancy provided by gas vesicles for planktonic cyanobacteria in the Baltic Sea. New Phytol. 1997, 136, 407–417. [Google Scholar] [CrossRef]
- Paerl, H.W.; Tucker, J.; Bland, P.T. Carotenoid enhancement and its role in maintaining blue-green algal (Microcystis aeruginosa) surface blooms. Limnol. Oceanogr. 1983, 28, 847–857. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Potential Toxin(s) | Characteristic | Salinity Range | ||
|---|---|---|---|---|---|
| Low (0–4) | Mod. (4–16) | High (16+) | |||
| Anabaenopsis | MC | P,D,F | X | X | X |
| Aphanizomenon | ATX, CYN, STX | P,D,F | X | X | |
| Cylindrospermopsis | ATX, CYN, STX | P,D,F | X | ||
| Cylindrospermum | ATX, MC | B,D,F | X | ||
| Dolichospermum | ATX, CYN, MC, STX | P,D,F | X | X | |
| Fischerella | MC | B,D,F | X | X | X |
| Hapalosiphon | MC | B,D,F | X | ||
| Lyngbya | CYN, LYN, STX | B,F | X | X | X |
| Microcystis | MC | P,C | X | ||
| Nodularia | NOD | B/P,D,F | X | X | X |
| Nostoc | ATX, MC | B,D,F | X | X | |
| Oscillatoria | ATX, CYN, MC, STX | B/P,D,F | X | X | X |
| Phormidium | ATX, MC | B,F | X | X | X |
| Planktothrix | ATX, MC | P,F | X | X | |
| Raphidiopsis | ATX, CYN, MC | P,F | X | X | |
| Scytonema | MC, STX | B,D,F | X | X | X |
| Umezakia | CYN, MC | P,D,F | X | ||
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paerl, H.W. Mitigating Toxic Planktonic Cyanobacterial Blooms in Aquatic Ecosystems Facing Increasing Anthropogenic and Climatic Pressures. Toxins 2018, 10, 76. https://doi.org/10.3390/toxins10020076
Paerl HW. Mitigating Toxic Planktonic Cyanobacterial Blooms in Aquatic Ecosystems Facing Increasing Anthropogenic and Climatic Pressures. Toxins. 2018; 10(2):76. https://doi.org/10.3390/toxins10020076
Chicago/Turabian StylePaerl, Hans W. 2018. "Mitigating Toxic Planktonic Cyanobacterial Blooms in Aquatic Ecosystems Facing Increasing Anthropogenic and Climatic Pressures" Toxins 10, no. 2: 76. https://doi.org/10.3390/toxins10020076
APA StylePaerl, H. W. (2018). Mitigating Toxic Planktonic Cyanobacterial Blooms in Aquatic Ecosystems Facing Increasing Anthropogenic and Climatic Pressures. Toxins, 10(2), 76. https://doi.org/10.3390/toxins10020076