Generation of Highly Efficient Equine-Derived Antibodies for Post-Exposure Treatment of Ricin Intoxications by Vaccination with Monomerized Ricin

 ,
,
Abstract
1. Introduction
2. Results
2.1. Elicitation of Anti-Ricin Antibodies Following Rabbit Immunization with Monomerized Ricin
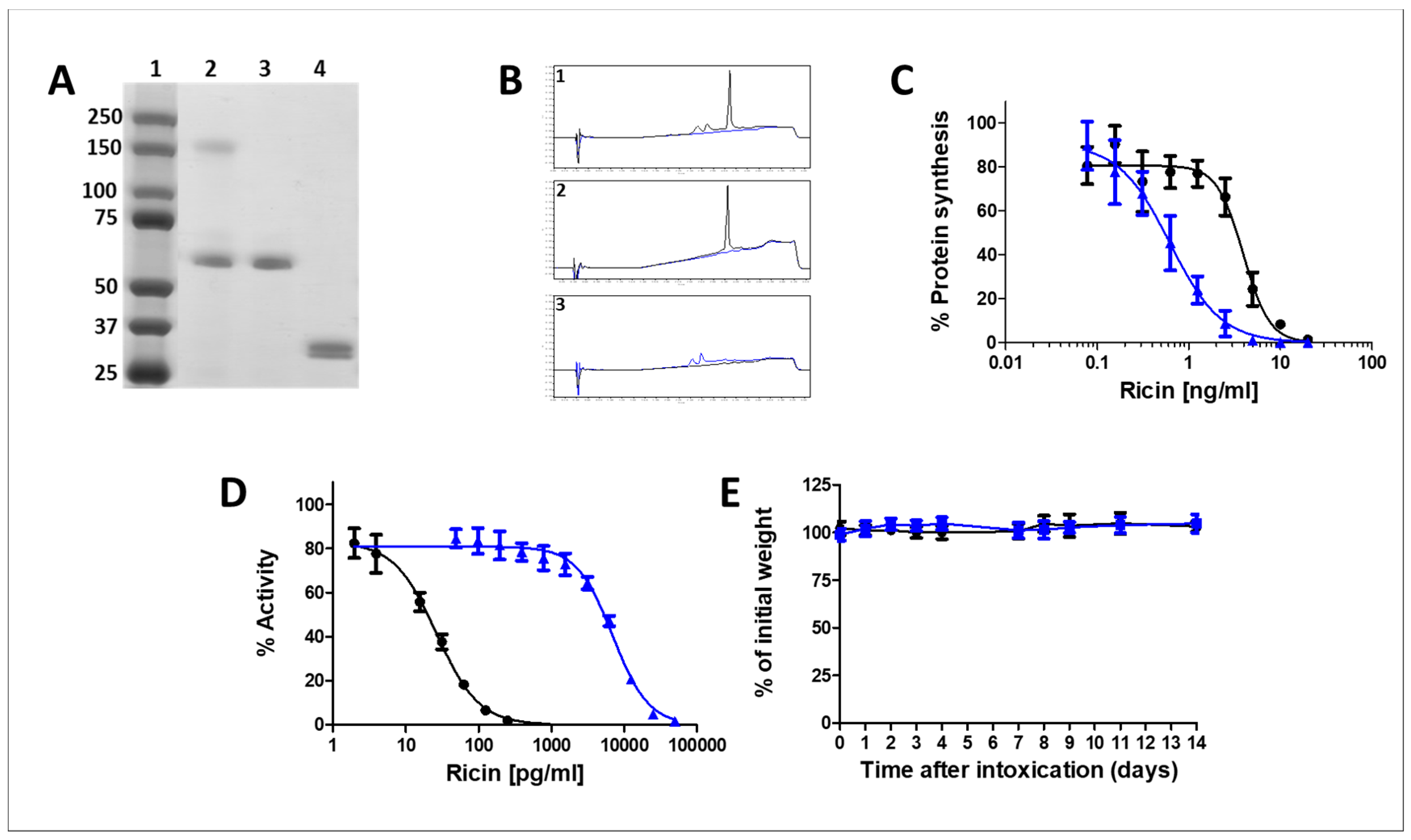
2.2. Characterization of the Monomerized Ricin Vaccine
2.2.1. Analytical Assessment
2.2.2. In-Vitro and In-Vivo Toxicity of the Monomerized Ricin
2.3. Anti-Ricin Titer Buildup Following Horse Immunization
2.4. In Vitro and In Vivo Efficacy of the Anti-Ricin Antitoxin
2.5. Horse Antitoxin Attenuates Ricin-Induced Pulmonary Damage Markers
3. Discussion
4. Materials and Methods
4.1. Ricin Preparation
4.2. Reduction and Alkylation of Pure Ricin
4.3. Gel Electrophoresis
4.4. UPLC
4.5. Isolation and Purification of Alkylated-Ricin Subunits
4.6. Assessment of Ricin Activity in a Cell-Free Translational Assay
4.7. Assessment of Ricin Activity in a Cell Culture
4.8. In Vitro Ricin Neutralization Assay
4.9. Animal Studies
4.10. Rabbit Anti-Ricin Hyperimmune Serum Production
4.11. Safety Studies in Mice
4.12. In Vivo Ricin Neutralization Determination
4.13. Survival Experiments in Mice
4.14. Bronchoalveolar Lavage Fluid (BALF) Analysis
4.15. Horse Vaccination and Plasmapheresis
4.16. F(ab’)2-Based Antitoxin Production
4.17. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Olsnes, S.; Kozlov, J.V. Ricin. Toxicon 2001, 39, 1723–1728. [Google Scholar] [CrossRef]
- Greenfield, R.A.; Brown, B.R.; Hutchins, J.B.; Iandolo, J.J.; Jackson, R.; Slater, L.N.; Bronze, M.S. Microbiological, biological, and chemical weapons of warfare and terrorism. Am. J. Med. Sci. 2002, 323, 326–340. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, J.M.; Brey, R.N., 3rd; Mantis, N.J. Comparative efficacy of two leading candidate ricin toxin a subunit vaccines in mice. Clin. Vaccine Immunol. 2013, 20, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Cohen, O.; Mechaly, A.; Sabo, T.; Alcalay, R.; Aloni-Grinstein, R.; Seliger, N.; Kronman, C.; Mazor, O. Characterization and epitope mapping of the polyclonal antibody repertoire elicited by ricin holotoxin-based vaccination. Clin. Vaccine. Immunol. 2014, 21, 1534–1540. [Google Scholar] [CrossRef] [PubMed]
- Pincus, S.H.; Das, A.; Song, K.; Maresh, G.A.; Corti, M.; Berry, J. Role of fc in antibody-mediated protection from ricin toxin. Toxins 2014, 6, 1512–1525. [Google Scholar] [CrossRef] [PubMed]
- Pratt, T.S.; Pincus, S.H.; Hale, M.L.; Moreira, A.L.; Roy, C.J.; Tchou-Wong, K.M. Oropharyngeal aspiration of ricin as a lung challenge model for evaluation of the therapeutic index of antibodies against ricin a-chain for post-exposure treatment. Exp. Lung. Res. 2007, 33, 459–481. [Google Scholar] [CrossRef] [PubMed]
- Vance, D.J.; Tremblay, J.M.; Mantis, N.J.; Shoemaker, C.B. Stepwise engineering of heterodimeric single domain camelid vhh antibodies that passively protect mice from ricin toxin. J. Biol. Chem. 2013, 288, 36538–36547. [Google Scholar] [CrossRef] [PubMed]
- Maddaloni, M.; Cooke, C.; Wilkinson, R.; Stout, A.V.; Eng, L.; Pincus, S.H. Immunological characteristics associated with the protective efficacy of antibodies to ricin. J. Immunol. 2004, 172, 6221–6228. [Google Scholar] [CrossRef] [PubMed]
- Foxwell, B.M.; Detre, S.I.; Donovan, T.A.; Thorpe, P.E. The use of anti-ricin antibodies to protect mice intoxicated with ricin. Toxicology 1985, 34, 79–88. [Google Scholar] [CrossRef]
- Noy-Porat, T.; Rosenfeld, R.; Ariel, N.; Epstein, E.; Alcalay, R.; Zvi, A.; Kronman, C.; Ordentlich, A.; Mazor, O. Isolation of anti-ricin protective antibodies exhibiting high affinity from immunized non-human primates. Toxins 2016, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Pincus, S.H.; Smallshaw, J.E.; Song, K.; Berry, J.; Vitetta, E.S. Passive and active vaccination strategies to prevent ricin poisoning. Toxins 2011, 3, 1163–1184. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, S.J.C.; Griffiths, G.D.; Jenner, D.C.; Gwyther, R.J.; Stahl, F.M.; Cork, L.J.; Holley, J.L.; Green, A.C.; Clark, G.C. Production, characterisation and testing of an ovine antitoxin against ricin; efficacy, potency and mechanisms of action. Toxins 2017, 9, 329. [Google Scholar] [CrossRef] [PubMed]
- Sabo, T.; Kronman, C.; Mazor, O. Ricin-holotoxin-based vaccines: Induction of potent ricin-neutralizing antibodies. Methods Mol. Biol. 2016, 1403, 683–694. [Google Scholar] [PubMed]
- Hale, M.L. Microtiter-based assay for evaluating the biological activity of ribosome-inactivating proteins. Pharmacol. Toxicol. 2001, 88, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Langer, M.; Rothe, M.; Eck, J.; Mockel, B.; Zinke, H. A nonradioactive assay for ribosome-inactivating proteins. Anal. Biochem. 1996, 243, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Kronman, C.; Velan, B.; Gozes, Y.; Leitner, M.; Flashner, Y.; Lazar, A.; Marcus, D.; Sery, T.; Papier, Y.; Grosfeld, H.; et al. Production and secretion of high levels of recombinant human acetylcholinesterase in cultured cell lines: Microheterogeneity of the catalytic subunit. Gene 1992, 121, 295–304. [Google Scholar] [CrossRef]
- Bielory, L.; Kemeny, D.M.; Richards, D.; Lessof, M.H. Igg subclass antibody production in human serum sickness. J. Allergy Clin. Immunol. 1990, 85, 573–577. [Google Scholar] [CrossRef]
- Lu, L.L.; Suscovich, T.J.; Fortune, S.M.; Alter, G. Beyond binding: Antibody effector functions in infectious diseases. Nat. Rev. Immunol. 2018, 18, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, F.; Ravetch, J.V. Fcgamma receptors as regulators of immune responses. Nat. Rev. Immunol. 2008, 8, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Gal, Y.; Mazor, O.; Alcalay, R.; Seliger, N.; Aftalion, M.; Sapoznikov, A.; Falach, R.; Kronman, C.; Sabo, T. Antibody/doxycycline combined therapy for pulmonary ricinosis: Attenuation of inflammation improved survival of ricin-intoxicated mice. Toxicol. Rep. 2014, 1, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Gal, Y.; Sapoznikov, A.; Falach, R.; Ehrlich, S.; Aftalion, M.; Kronman, C.; Sabo, T. Total body irradiation mitigates inflammation and extends the therapeutic time window for anti-ricin antibody treatment against pulmonary ricinosis in mice. Toxins 2017, 9, 278. [Google Scholar] [CrossRef] [PubMed]
- Gal, Y.; Sapoznikov, A.; Falach, R.; Ehrlich, S.; Aftalion, M.; Sabo, T.; Kronman, C. Potent antiedematous and protective effects of ciprofloxacin in pulmonary ricinosis. Antimicrob Agents Chemother. 2016, 60, 7153–7158. [Google Scholar] [PubMed]
- Lindauer, M.L.; Wong, J.; Iwakura, Y.; Magun, B.E. Pulmonary inflammation triggered by ricin toxin requires macrophages and il-1 signaling. J. Immunol. 2009, 183, 1419–1426. [Google Scholar] [CrossRef] [PubMed]
- Locher, C.P.; Tam, L.Q. Reduction of disulfide bonds in plasmodium falciparum gp195 abolishes the production of growth-inhibitory antibodies. Vaccine 1993, 11, 1119–1123. [Google Scholar] [CrossRef]
- Sabo, T.; Gal, Y.; Elhanany, E.; Sapoznikov, A.; Falach, R.; Mazor, O.; Kronman, C. Antibody treatment against pulmonary exposure to abrin confers significantly higher levels of protection than treatment against ricin intoxication. Toxicol. Lett. 2015, 237, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Komaki, Y.; Sugiura, H.; Koarai, A.; Tomaki, M.; Ogawa, H.; Akita, T.; Hattori, T.; Ichinose, M. Cytokine-mediated xanthine oxidase upregulation in chronic obstructive pulmonary disease’s airways. Pulm. Pharmacol. Ther. 2005, 18, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.M.; Ginger, L.A.; Kosila, N.; Elkins, N.D.; Essary, B.; McManaman, J.L.; Repine, J.E. Mononuclear phagocyte xanthine oxidoreductase contributes to cytokine-induced acute lung injury. Am. J. Respir. Cell Mol. Biol. 2004, 30, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Faggioni, R.; Gatti, S.; Demitri, M.T.; Delgado, R.; Echtenacher, B.; Gnocchi, P.; Heremans, H.; Ghezzi, P. Role of xanthine oxidase and reactive oxygen intermediates in lps- and tnf-induced pulmonary edema. J. Lab. Clin. Med. 1994, 123, 394–399. [Google Scholar] [PubMed]
- Grosso, M.A.; Brown, J.M.; Viders, D.E.; Mulvin, D.W.; Banerjee, A.; Velasco, S.E.; Repine, J.E.; Harken, A.H. Xanthine oxidase-derived oxygen radicals induce pulmonary edema via direct endothelial cell injury. J. Surg. Res. 1989, 46, 355–360. [Google Scholar] [CrossRef]
- Lin, J.Y.; Liu, S.Y. Studies on the antitumor lectins isolated from the seeds of ricinus communis (castor bean). Toxicon 1986, 24, 757–765. [Google Scholar] [CrossRef]
- Griffiths, G.D.; Lindsay, C.D.; Upshall, D.G. Examination of the toxicity of several protein toxins of plant origin using bovine pulmonary endothelial cells. Toxicology 1994, 90, 11–27. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Antigen | ELISA-Antibody Titer a | Neutralizing-Antibody Titer a | PD50 (μL) b |
|---|---|---|---|
| Native Toxin | 256,000 ± 41,300 | 76,800 ± 14,300 | 1.1 ± 0.31 |
| Monomerized Toxin | 480,000 ± 71,500 | 96,000 ± 14,300 | 0.8 ± 0.14 |
| Treatment Group | |||
|---|---|---|---|
| Naïve a | Ricin b | Ricin + Antitoxin c | |
| IL-6 (pg/mL) | 0 ± 0 | 3547 ± 1372 ** | 451 ± 532 && |
| Protein (mg/mL) | 0.5 ± 0.1 | 5.7 ± 1.8 ** | 2.1 ± 0.8 **&& |
| ChE (mU/mL) | 0 ± 0 | 306 ± 93 ** | 68 ± 31 **&& |
| XO (mU/mL) | 0.6 ± 0.1 | 4.1 ± 1.5 ** | 1.7 ± 0.7 *&& |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falach, R.; Sapoznikov, A.; Alcalay, R.; Aftalion, M.; Ehrlich, S.; Makovitzki, A.; Agami, A.; Mimran, A.; Rosner, A.; Sabo, T.; et al. Generation of Highly Efficient Equine-Derived Antibodies for Post-Exposure Treatment of Ricin Intoxications by Vaccination with Monomerized Ricin. Toxins 2018, 10, 466. https://doi.org/10.3390/toxins10110466
Falach R, Sapoznikov A, Alcalay R, Aftalion M, Ehrlich S, Makovitzki A, Agami A, Mimran A, Rosner A, Sabo T, et al. Generation of Highly Efficient Equine-Derived Antibodies for Post-Exposure Treatment of Ricin Intoxications by Vaccination with Monomerized Ricin. Toxins. 2018; 10(11):466. https://doi.org/10.3390/toxins10110466
Chicago/Turabian StyleFalach, Reut, Anita Sapoznikov, Ron Alcalay, Moshe Aftalion, Sharon Ehrlich, Arik Makovitzki, Avi Agami, Avishai Mimran, Amir Rosner, Tamar Sabo, and et al. 2018. "Generation of Highly Efficient Equine-Derived Antibodies for Post-Exposure Treatment of Ricin Intoxications by Vaccination with Monomerized Ricin" Toxins 10, no. 11: 466. https://doi.org/10.3390/toxins10110466
APA StyleFalach, R., Sapoznikov, A., Alcalay, R., Aftalion, M., Ehrlich, S., Makovitzki, A., Agami, A., Mimran, A., Rosner, A., Sabo, T., Kronman, C., & Gal, Y. (2018). Generation of Highly Efficient Equine-Derived Antibodies for Post-Exposure Treatment of Ricin Intoxications by Vaccination with Monomerized Ricin. Toxins, 10(11), 466. https://doi.org/10.3390/toxins10110466