Anti-Breast Cancer Activity of Latroeggtoxin-V Mined from the Transcriptome of Spider Latrodectus tredecimguttatus Eggs

Abstract
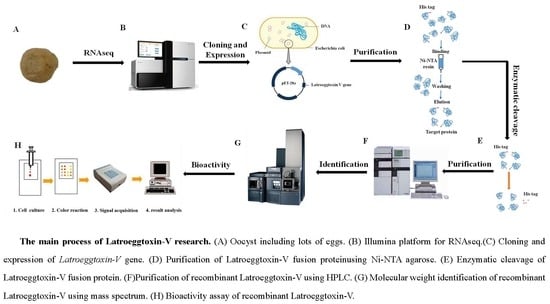
1. Introduction
2. Results
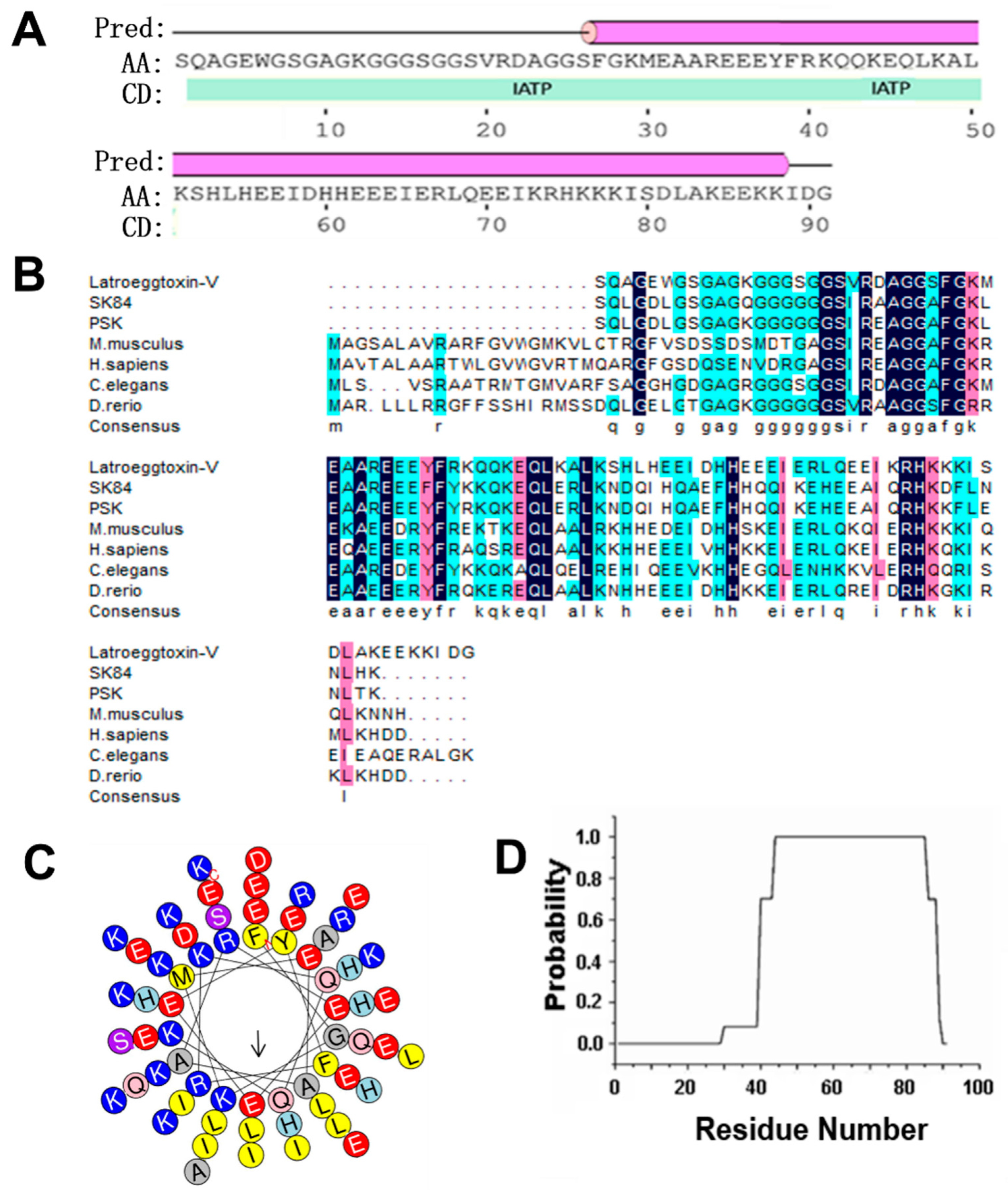
2.1. Bioinformatic Analysis on Latroeggtoxin-V
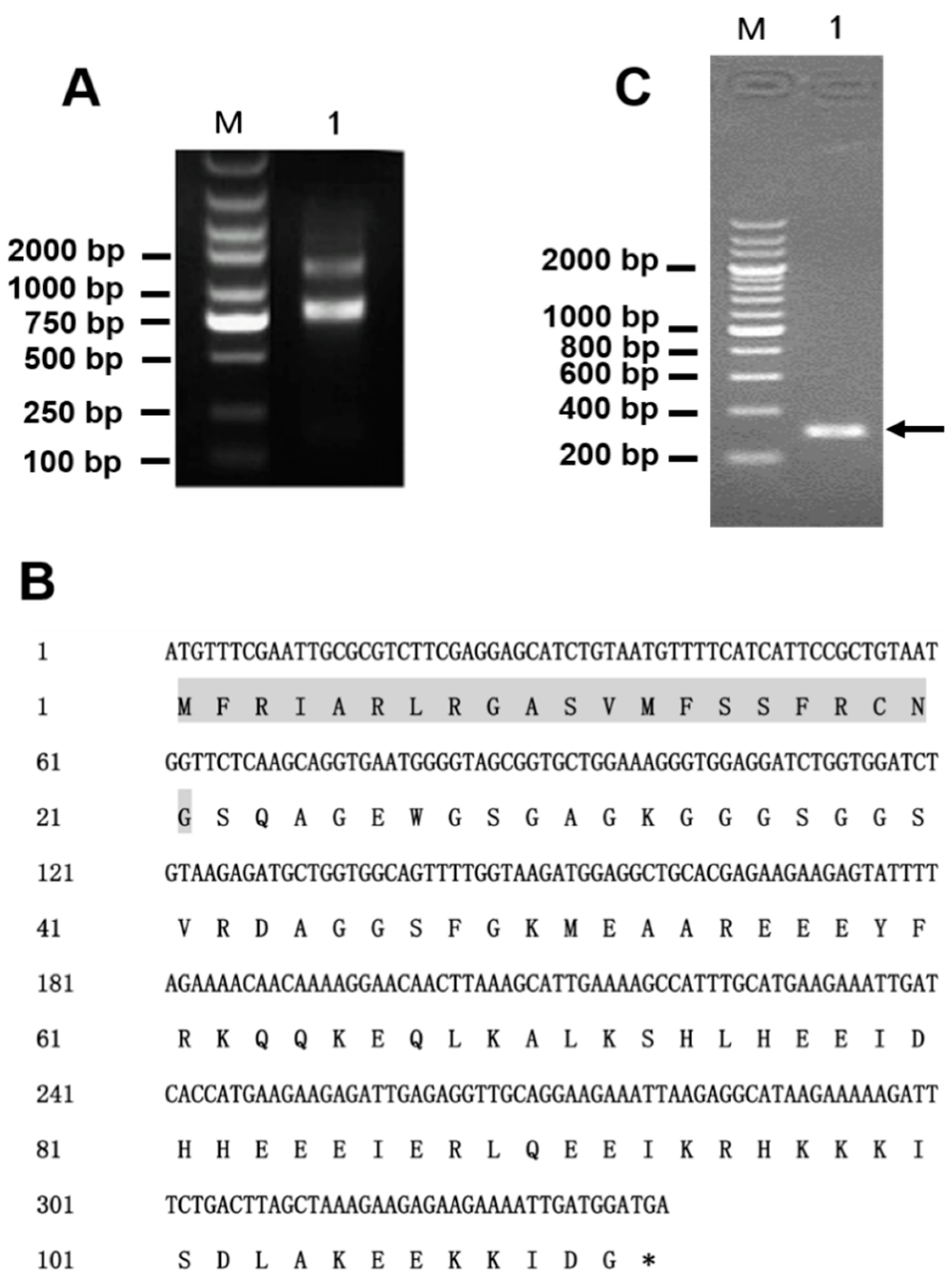
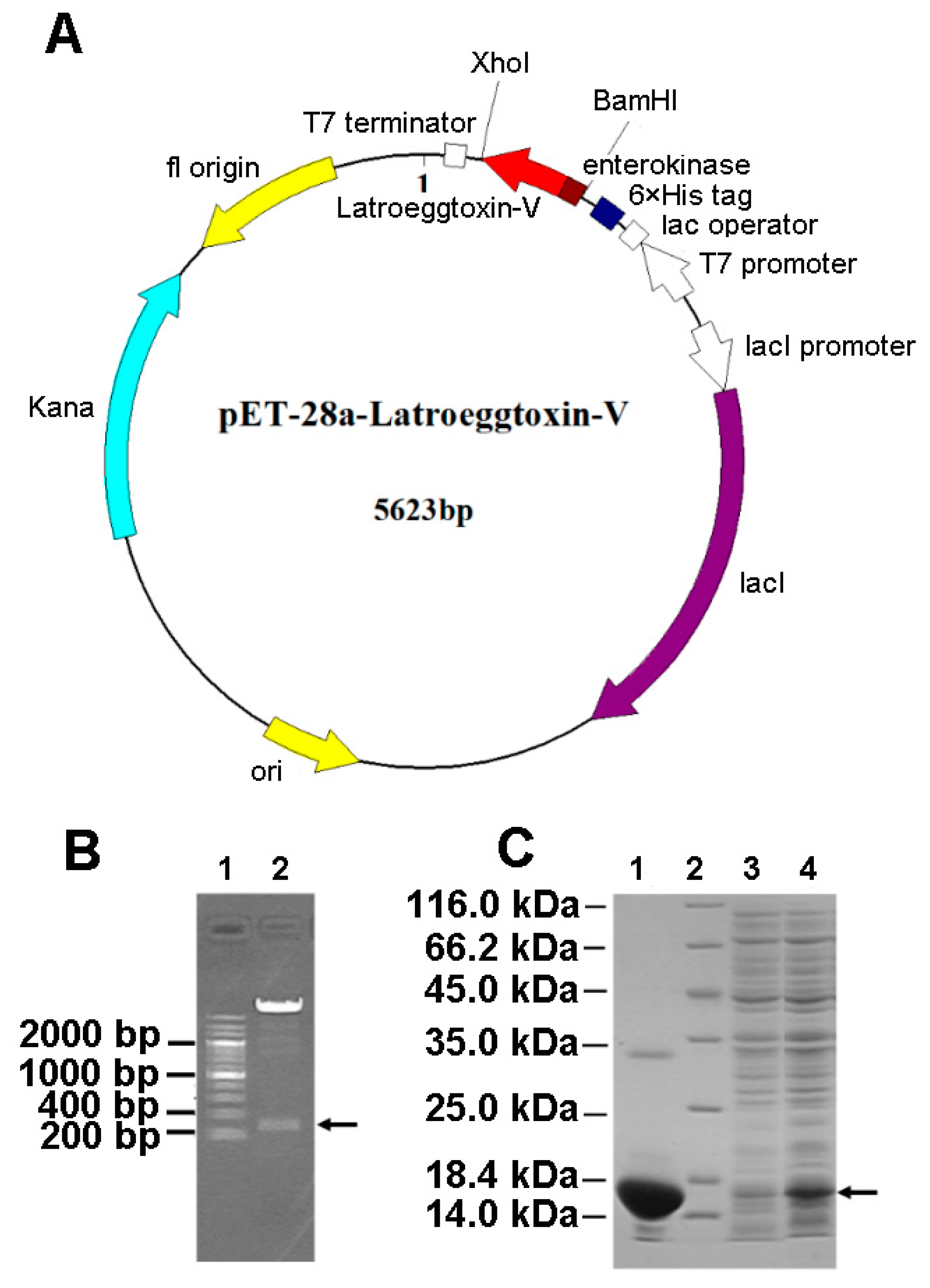
2.2. Gene Cloning of Latroeggtoxin-V
2.3. Prokaryotic Expression and Purification of Latroeggtoxin-V Fusion Protein
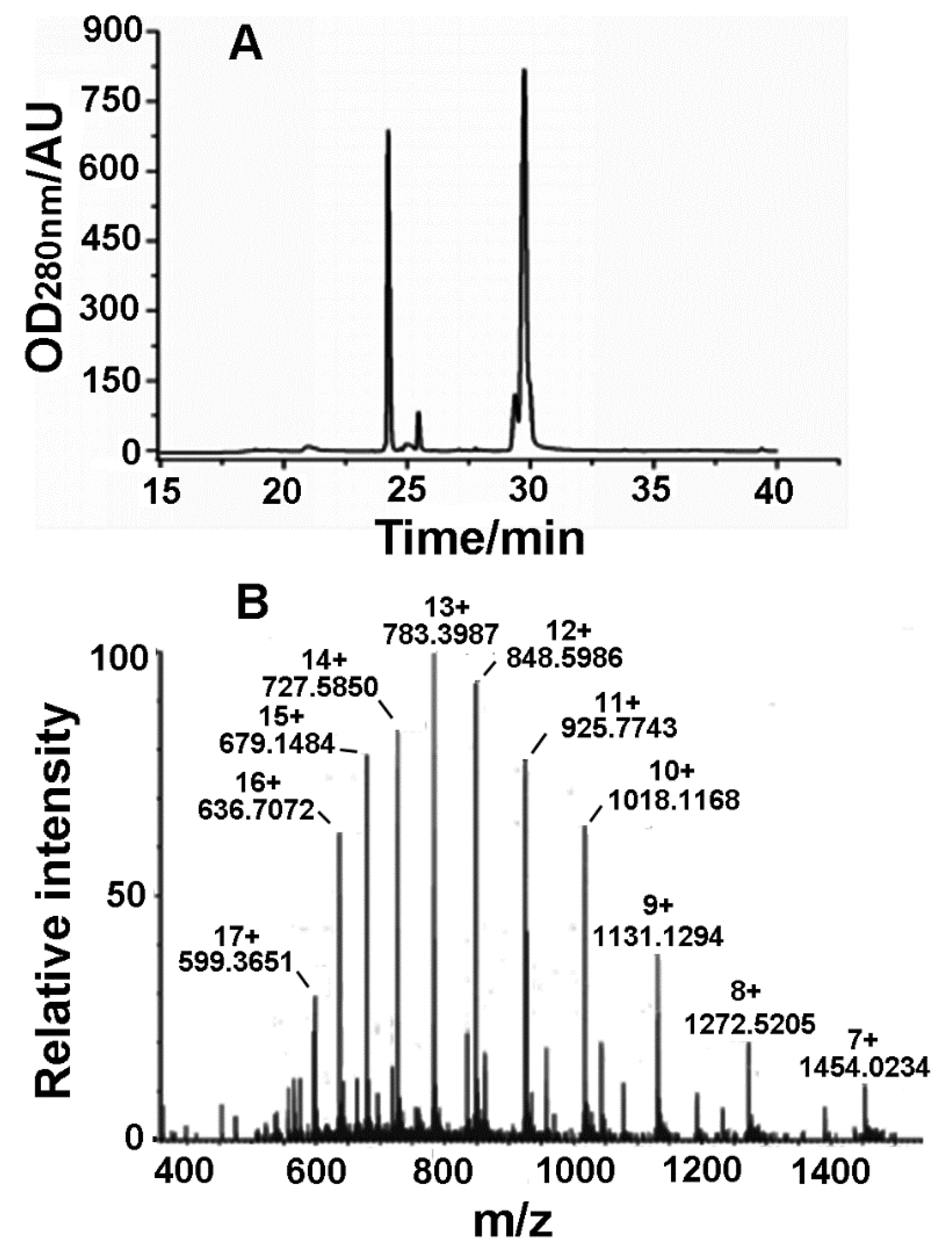
2.4. Acquisition and Identification of rLatroeggtoxin-V
2.5. Effects of rLatroeggtoxin-V on Cell Viability
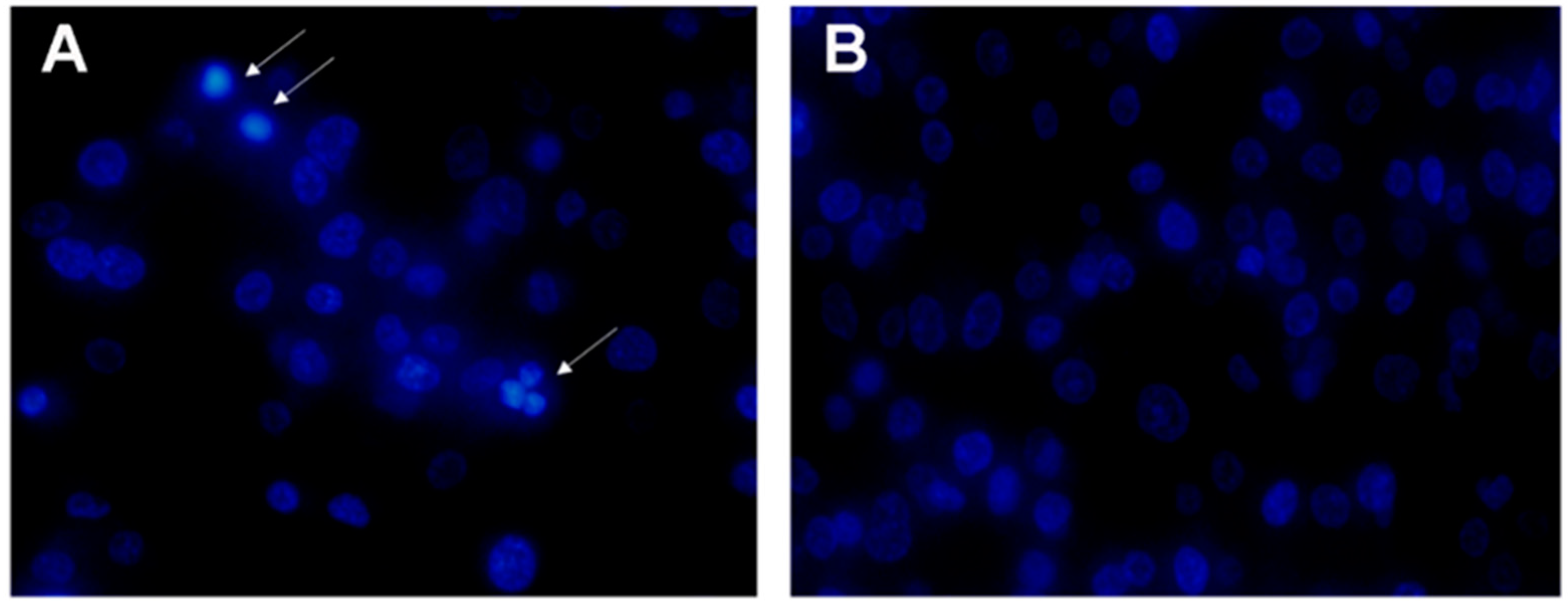
2.6. Apoptosis of MDA-MB-231 Cells Induced by rLatroeggtoxin-V
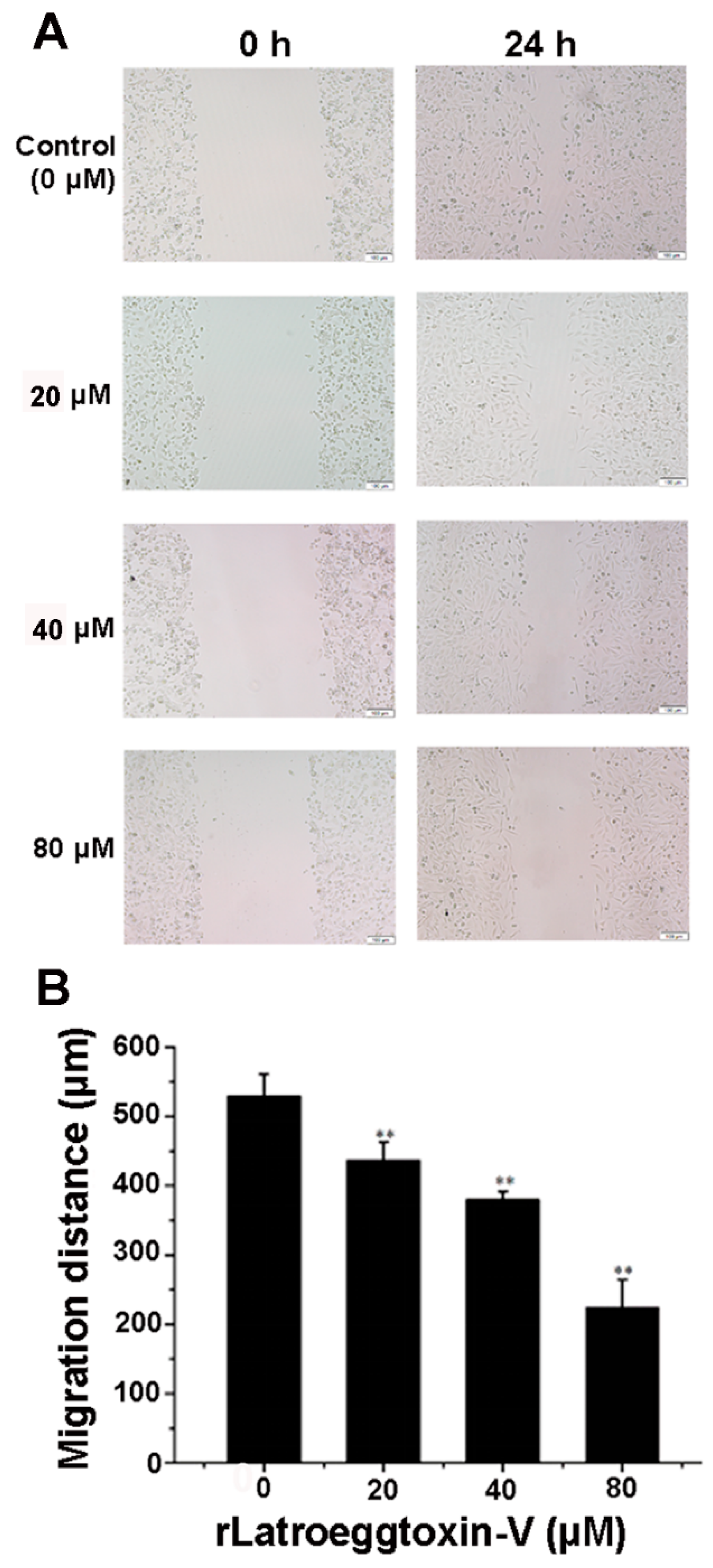
2.7. Inhibition of MDA-MB-231 Cell Migration by rLatroeggtoxin-V
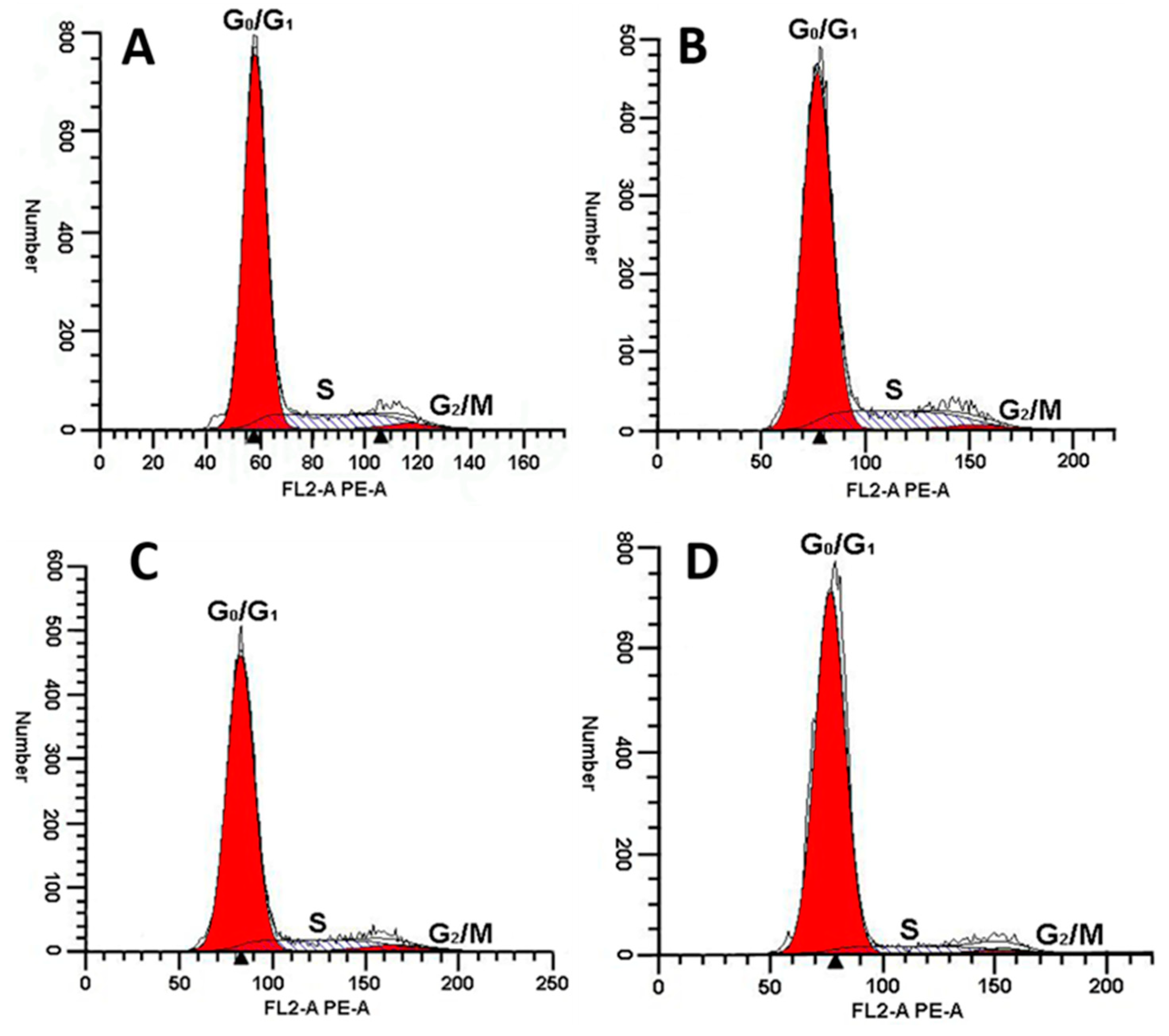
2.8. MDA-MB-231 Cell Cycle Arrest Caused by rLatroeggtoxin-V
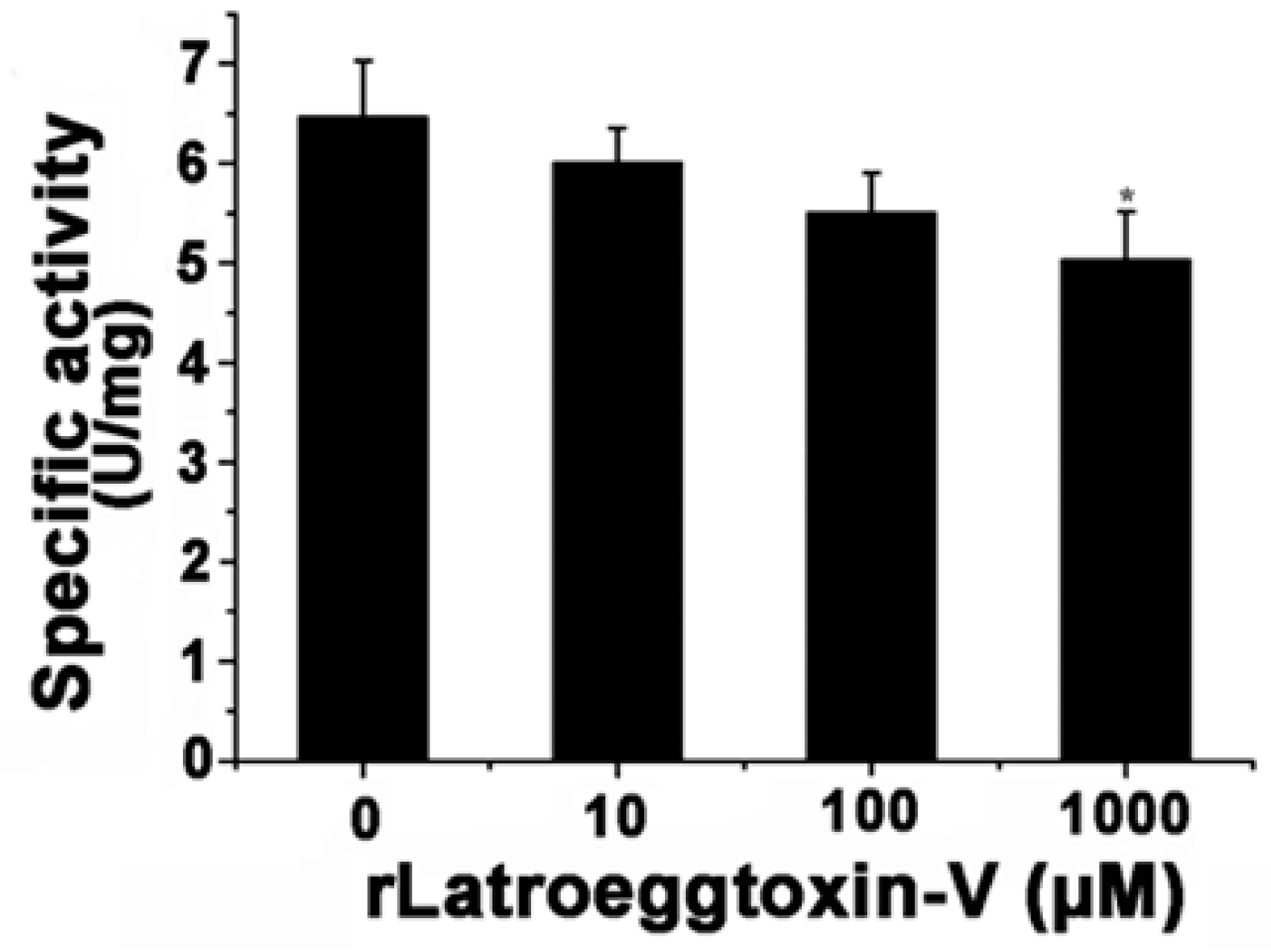
2.9. rLatroeggtoxin-V Inhibition of Na+/K+-ATPase of MDA-MB-231 Cells
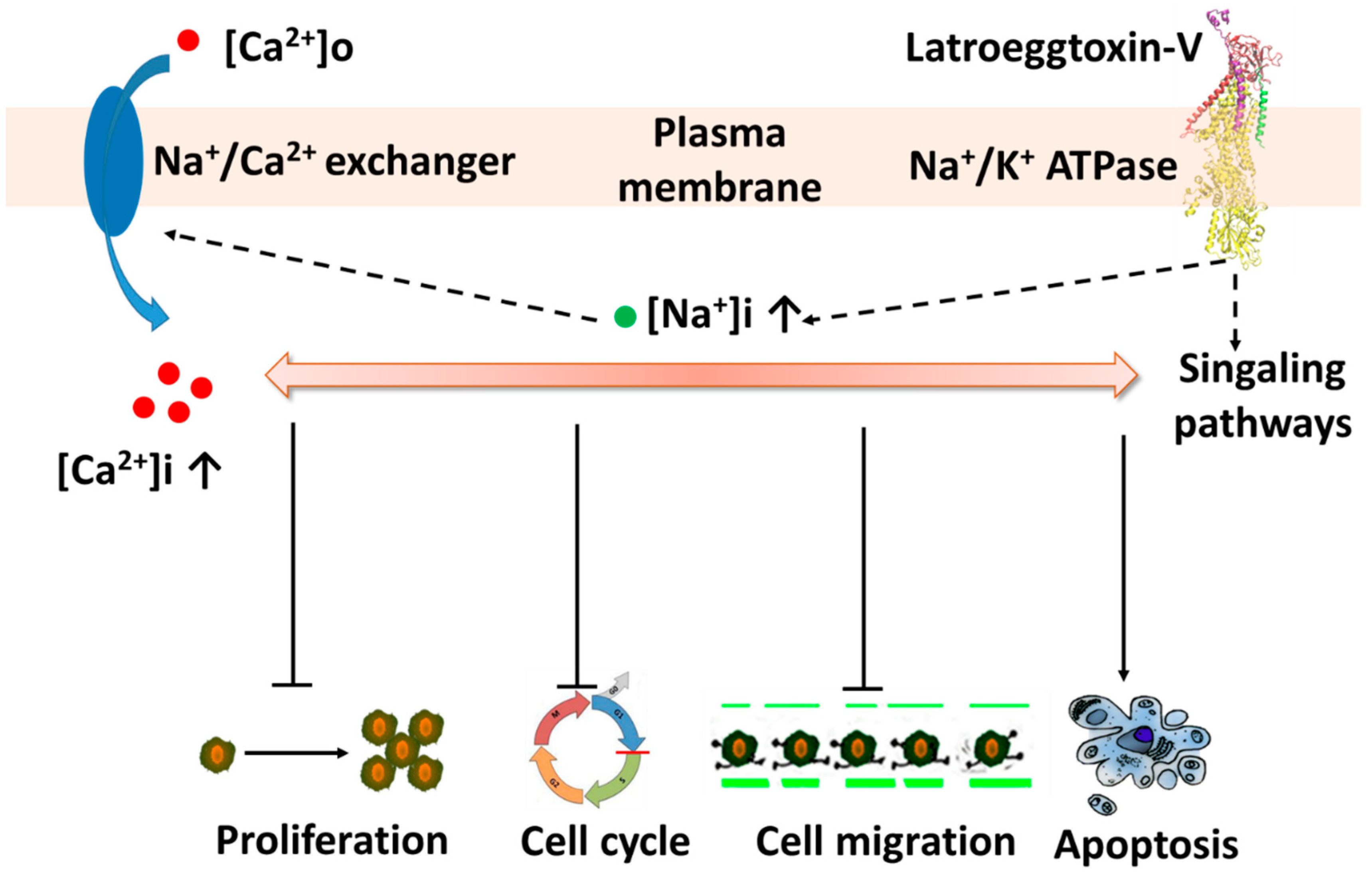
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Bioinformatic Analysis of Latroeggtoxin-V
5.2. Gene Cloning of Latroeggtoxin-V
5.3. Prokaryotic Expression of Latroeggtoxin-V
5.4. Purification of Latroeggtoxin-V Fusion Protein
5.5. Cleavage of Latroeggtoxin-V Fusion Protein and Isolation of rLatroeggtoxin-V
5.6. Bioactivity Assessment of rLatroeggtoxin-V against MDA-MB-231 Cells
5.6.1. Cell Viability-Inhibiting Activity
5.6.2. Apoptosis-Inducing Activity
5.6.3. Cell Migration-Inhibiting Activity
5.6.4. Cell Cycle Arrest Activity
5.6.5. Na+/K+-ATPase-Inhibiting Activity
5.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- World Spider Catalog. World Spider Catalog Version 19.5. Natural History Museum Bern, 2018. Available online: http://wsc.nmbe.ch/ (accessed on 14 August 2018).
- Zhu, M.S. The Spiders of China Arachnida Araneae Theridiidae; Science Press: Beijing, China, 1998; pp. 293–294. ISBN 9787030062437. (In Chinese) [Google Scholar]
- Camp, N.E. Black widow spider envenomation. J. Emerg. Nurs. 2014, 40, 193–194. [Google Scholar] [CrossRef] [PubMed]
- Hoxha, R. Two Albanians die from black widow spider bites. BMJ 2006, 333, 278. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wang, X.C. Recent Advances in Research on Widow Spider Venoms and Toxins. Toxins 2015, 7, 5055–5067. [Google Scholar] [CrossRef] [PubMed]
- He, Q.Z.; Duan, Z.G.; Yu, Y.; Liu, Z.; Liu, Z.H.; Liang, S.P. The venom gland transcriptome of Latrodectus tredecimguttatus revealed by deep sequencing and cDNA library analysis. PLoS ONE 2013, 8, e81357. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Duan, Z.G.; Yang, J.; Yan, X.J.; Zhou, H.; He, X.Z.; Liang, S.P. Physiological and biochemical analysis of L. tredecimguttatus venom collected by electrical stimulation. J. Physiol. Biochem. 2007, 63, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.G.; Yan, X.J.; He, X.Z.; Zhou, H.; Chen, P.; Cao, R.; Xiong, J.X.; Hu, W.J.; Wang, X.C.; Liang, S.P. Extraction and protein component analysis of venom from the dissected venom glands of Latrodectus tredecimguttatus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 145, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.G.; Yang, J.; Yan, X.J.; Zeng, X.Z.; Wang, X.C.; Liang, S.P. Venom properties of the spider Latrodectus tredecimguttatus and comparison of two venom-collecting methods. Zool. Res. 2009, 30, 381–388. [Google Scholar] [CrossRef]
- Buffkin, D.C.; Russell, F.E.; Deshmukh, A. Preliminary study on the toxicity of black widow spider eggs. Toxicon 1971, 9, 393–402. [Google Scholar] [CrossRef]
- Russell, F.E.; Maretć, Z. Effects of Latrodectus egg poison on web building. Toxicon 1979, 17, 649–650. [Google Scholar] [CrossRef]
- Yan, Y.Z.; Li, J.J.; Zhang, Y.Y.; Peng, X.Z.; Guo, T.Y.; Wang, J.R.; Hu, W.J.; Duan, Z.G.; Wang, X.C. Physiological and biochemical characterization of egg extract of black widow spiders to uncover molecular basis of egg toxicity. Biol. Res. 2014, 47, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Liu, H.; Duan, Z.G.; Cao, R.; Wang, X.C.; Liang, S.P. Protein compositional analysis of the eggs of black widow spider (Latrodectus tredecimguttatus): Implications for the understanding of egg toxicity. J. Biochem. Mol. Toxicol. 2012, 26, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Yan, Y.Z.; Wang, J.R.; Guo, T.Y.; Hu, W.J.; Duan, Z.G.; Wang, X.C.; Liang, S.P. Purification and partial characterization of a novel neurotoxic protein from eggs of black widow spiders (Latrodectus tredecimguttatus). J. Biochem. Mol. Toxicol. 2013, 27, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Yan, Y.Z.; Yu, H.; Peng, X.Z.; Zhang, Y.Y.; Hu, W.J.; Duan, Z.G.; Wang, X.C.; Liang, S.P. Isolation and identification of a sodium channel-inhibiting protein from eggs of black widow spiders. Int. J. Biol. Macromol. 2014, 65, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.; Yu, H.; Peng, X.Z.; Yan, S.; Wang, J.R.; Yan, Y.Z.; Wang, X.C. Isolation and preliminary characterization of proteinaceous toxins with insecticidal and antibacterial activities from black widow spider (L. tredecimguttatus) eggs. Toxins 2015, 7, 886–899. [Google Scholar] [CrossRef] [PubMed]
- Nagalakshmi, U.; Waern, K.; Snyder, M. RNA-Seq: A Method for Comprehensive Transcriptome Analysis. Curr. Protoc. Mol. Biol. 2010, 89, 1–13. [Google Scholar] [CrossRef]
- Hoeijmakers, W.A.; Bártfai, R.; Stunnenberg, H.G. Transcriptome analysis using RNA-Seq. Met. Mol. Biol. 2013, 923, 221–239. [Google Scholar] [CrossRef]
- Xu, D.H.; Wang, X.C. Transcriptome Analysis to Understand the Toxicity of Latrodectus tredecimguttatus Eggs. Toxins 2016, 8, 378. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Chen, Z.W. Isolation, characterization and anti-cancer activity of SK84, a novel glycine-rich antimicrobial peptide from Drosophila virilis. Peptides 2010, 31, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Chen, Z.W.; Wu, Y.; Ding, J.P.; Cederlund, E.; Jörnvall, H.; Bergman, T. An ATPase inhibitory peptide with antibacterial and ion current effects. Biochem. Biophys. Res. Commun. 2014, 446, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.; Meier, I. Scaffolds, levers, rods and springs: Diverse cellular functions of long coiled-coil proteins. Cell. Mol. Life Sci. 2004, 61, 1996–2009. [Google Scholar] [CrossRef] [PubMed]
- Geering, K. Na, K-ATPase. Curr. Opin. Nephrol. Hypertens. 1997, 6, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Aperia, A. New roles for an old enzyme: Na, K-ATPase emerges as an interesting drug target. J. Intern. Med. 2007, 261, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.P. Na+, K+-ATPase: The new face of an old player in pathogenesis and apoptotic/hybrid cell death. Biochem. Pharmacol. 2003, 66, 1601–1609. [Google Scholar] [CrossRef]
- Xiao, A.Y.; Wei, L.; Xia, S.; Rothman, S.; Yu, S.P. Ionic mechanism of ouabain-induced concurrent apoptosis and necrosis in individual cultured cortical neurons. J. Neurosci. 2002, 22, 1350–1362. [Google Scholar] [CrossRef] [PubMed]
- Archibald, J.T.; White, T.D. Rapid reversal of internal Na+ and K+ contents of synaptosomes by ouabain. Nature 1974, 252, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Aperia, A.; Akkuratov, E.E.; Fontana, J.M.; Brismar, H. Na-K-ATPase, a new class of plasma membrane receptors. Am. J. Physiol. Cell Physiol. 2016, 310, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Pierre, S.V.; Xie, Z. The Na, K-ATPase receptor complex: Its organization and membership. Cell Biochem. Biophys. 2006, 46, 303–316. [Google Scholar] [CrossRef]
- Chen, J.Q.; Contreras, R.G.; Wang, R.; Fernandez, S.V.; Shoshani, L.; Russo, I.H.; Cereijido, M.; Russo, J. Sodium/potassium ATPase (Na+, K+-ATPase) and ouabain/related cardiac glycosides: A new paradigm for development of anti- breast cancer drugs? Breast Cancer Res. Treat. 2006, 96, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Felippe Gonçalves-de-Albuquerque, C.; Ribeiro Silva, A.; Ignácio da Silva, C.; Caire Castro-Faria-Neto, H.; Burth, P. Na/K pump and beyond: Na/K-ATPase as a modulator of apoptosis and autophagy. Molecules 2017, 22, 578. [Google Scholar] [CrossRef] [PubMed]
- Alevizopoulos, K.; Calogeropoulou, T.; Lang, F.; Stournaras, C. Na+/K+ ATPase inhibitors in cancer. Curr. Drug Targets 2014, 15, 988–1000. [Google Scholar] [CrossRef] [PubMed]
- Mijatovic, T.; Dufrasne, F.; Kiss, R. Na+/K+-ATPase and cancer. Pharm Pat. Anal. 2012, 1, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Magpusao, A.N.; Omolloh, G.; Johnson, J.; Gascón, J.; Peczuh, M.W.; Fenteany, G. Cardiac glycoside activities link Na+/K+ ATPase ion-transport to breast cancer cell migration via correlative SAR. ACS Chem. Biol. 2015, 10, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Kometiani, P.; Liu, L.; Askari, A. Digitalis-induced signaling by Na+/K+-ATPase in human breast cancer cells. Mol. Pharmacol. 2005, 67, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.W. Cancer and radiation therapy: Current advances and future directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Libério, M.S.; Joanitti, G.A.; Fontes, W.; Castro, M.S. Anticancer peptides and proteins: A panoramic view. Protein Pept. Lett. 2013, 20, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xie, S.H.; Xu, B.; Lu, S.; Liu, P. Digitalis use and the risk of breast cancer: A systematic review and meta-analysis. Drug Saf. 2017, 40, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Ahern, T.P.; Tamimi, R.M.; Rosner, B.A.; Hankinson, S.E. Digoxin use and risk of invasive breast cancer: Evidence from the Nurses’ Health Study and meta-analysis. Breast Cancer Res. Treat. 2014, 144, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Gao, Y.; Qi, Y.; Chen, L.; Ma, Y.; Li, Y. Peptide-based cancer therapy: Opportunity and challenge. Cancer Lett. 2014, 351, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. Why do we study animal toxins? Zool. Res. 2015, 36, 183–222. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.J.; Choi, Y.; Shin, J.M.; Cho, H.J.; Kang, J.H.; Park, K.K.; Choe, J.Y.; Bae, Y.S.; Han, S.M.; Kim, C.H.; et al. Melittin suppresses EGF-induced cell motility and invasion by inhibiting PI3K/Akt/mTOR signaling pathway in breast cancer cells. Food Chem. Toxicol. 2014, 68, 218–225. [Google Scholar] [CrossRef] [PubMed]
- D’Suze, G.; Rosales, A.; Salazar, V.; Sevcik, C. Apoptogenic peptides from Tityus discrepans scorpion venom acting against the SKBR3 breast cancer cell line. Toxicon 2010, 56, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Prajeep, M.; Mechkarska, M.; Arafat, K.; Attoub, S.; Adem, A.; Pla, D.; Calvete, J.J. Peptides with in vitro anti-tumor activity from the venom of the Eastern green mamba, Dendroaspis angusticeps (Elapidae). J. Venom Res. 2014, 5, 16–21. [Google Scholar] [PubMed]
- Cabezón, E.; Runswick, M.J.; Leslie, A.G.; Walker, J.E. The structure of bovine IF1, the regulatory subunit of mitochondrial F-ATPase. EMBO J. 2001, 20, 6990–6996. [Google Scholar] [CrossRef] [PubMed]
- Apostolovic, B.; Danial, M.; Klok, H.A. Coiled coils: Attractive protein folding motifs for the fabrication of self-assembled, responsive and bioactive materials. Chem. Soc. Rev. 2010, 39, 3541–3575. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Collier, J.H. α-Helical coiled-coil peptide materials for biomedical applications. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2017, 9, e1424. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.N.; Zhao, X.J.; Zhao, H.D.; Zhang, W.G.; Li, F.L.; Chen, M.Z.; Li, H.; Li, G. Recombinant Escherichia coli Trx-JZTX-III represses the proliferation of mouse hepatocellular carcinoma cells through induction of cell cycle arrest. Mol. Med. Rep. 2013, 7, 1800–1804. [Google Scholar] [CrossRef] [PubMed]
- El Moussawi, L.; Chakkour, M.; Kreydiyyeh, S.I. Epinephrine modulates Na+/K+ ATPase activity in Caco-2 cells via Src, p38MAPK, ERK and PGE2. PLoS ONE 2018, 13, e0193139. [Google Scholar] [CrossRef] [PubMed]
- El-Zein, O.; Usta, J.; El Moussawi, L.; Kreydiyyeh, S.I. Leptin inhibits the Na+/K+ ATPase in Caco-2 cells via PKC and p38MAPK. Cell. Signal. 2015, 27, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Esmann, M. ATPase and phosphatase activity of Na+, K+-ATPase: Molar and specific activity, protein determination. Methods Enzymol. 1988, 156, 105–109. [Google Scholar] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Cell Cycle Distribution (%) | ||
|---|---|---|---|
| G0/G1 | S | G2/M | |
| Control (0 μM) | 80.80 ± 0.69 | 16.22 ± 1.22 | 2.98 ± 0.7 |
| 25 μM | 81.59 ± 1.65 | 15.81 ± 1.91 | 2.98 ± 0.7 |
| 50 μM | 83.25 ± 0.73 a | 13.83 ± 0.77 a | 2.91 ± 0.15 |
| 100 μM | 89.67 ± 1.39 a | 8.33 ± 0.38 a | 2.00 ± 1.09 |
| Primer Name | Primer Sequence (5′→3′) |
|---|---|
| Latroeggtoxin-V-F | CATGGGATCCTCTCAGGCTGGTGAATGGGGTTCTG |
| Latroeggtoxin-V-R | CCGCTCGAGTTAACCGTCGATTTTTTTTTCTTCTTTAGC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, D.; Tang, X.; Wu, X.; Yu, D.; Tang, P.; Wang, X. Anti-Breast Cancer Activity of Latroeggtoxin-V Mined from the Transcriptome of Spider Latrodectus tredecimguttatus Eggs. Toxins 2018, 10, 451. https://doi.org/10.3390/toxins10110451
Xu D, Tang X, Wu X, Yu D, Tang P, Wang X. Anti-Breast Cancer Activity of Latroeggtoxin-V Mined from the Transcriptome of Spider Latrodectus tredecimguttatus Eggs. Toxins. 2018; 10(11):451. https://doi.org/10.3390/toxins10110451
Chicago/Turabian StyleXu, Dehong, Xiaochao Tang, Xinzhou Wu, Dianmei Yu, Pingping Tang, and Xianchun Wang. 2018. "Anti-Breast Cancer Activity of Latroeggtoxin-V Mined from the Transcriptome of Spider Latrodectus tredecimguttatus Eggs" Toxins 10, no. 11: 451. https://doi.org/10.3390/toxins10110451
APA StyleXu, D., Tang, X., Wu, X., Yu, D., Tang, P., & Wang, X. (2018). Anti-Breast Cancer Activity of Latroeggtoxin-V Mined from the Transcriptome of Spider Latrodectus tredecimguttatus Eggs. Toxins, 10(11), 451. https://doi.org/10.3390/toxins10110451