Oxazolone-Induced Delayed Type Hypersensitivity Reaction in the Adult Yucatan Pigs. A Useful Model for Drug Development and Validation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sensitization and challenging pigs with oxazolone
2.3. Other procedures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protocol for pigs in group 1 |
| Day 0: Intra-dermal sensitization with 2% oxazolone on back |
| Day 6, -12 hr: Bled (interleukin-2 (IL-2), interleukin-8 (IL-8), macrophage inflammatory pretin-1α (MIP-1α), and C-reactive protein (CRP) levels were measured); dosed (PO) |
| Day 6, 0 hr: Epi-dermal challenged with 1% oxazolone on ear |
| Day 6, +3 hr: Dosed (PO) |
| Day 6, +6 hr: Bled (IL-2, IL-8; MIP-1α, and CRP levels were measured) |
| Day 6-8: Pigs were sacrificed. Inflamed ears were drained, left and right ears were harvested and pictures taken. |
| Protocol for pigs in group 2 |
| Day 0: Intra-dermal sensitization with 2% oxazolone on back |
| Day 6, -12 hr: Bled (IL-2, IL-8; MIP-1α, and CRP levels were measured); dosed (PO) |
| Day 6, 0 hr: Epi-dermal challenge with 1% oxazolone on the ears |
| Day 6, +2 hr: Spleens were harvested from sensitized donor pigs |
| Day 6, +3 hr: Dosed (PO) |
| Day 6, +6 hr: Bled (IL-2, IL-8; MIP-1α, and CRP levels are measured); Injected (IV) with fluorescently labeled splenocytes |
| Day 6, +9hr: Pigs were sacrificed. Inflamed ears were drained (IL-2 level was measured, and the number of cells were calculated). Left and right ears were dissected and pictures taken. Dermis was dissected for measuring fluorescence intensity. |
| Spleens were harvested (cell isolation and T cell proliferation after PMA/IONO stimulation was conducted), blood was collected (IL-2, IL-8; MIP-1α, CRP levels were measured). |
2.4. Measurement of inflammatory mediators
2.5. T cell Proliferation and IL-2 release
2.6. Statistical analysis
3. Results
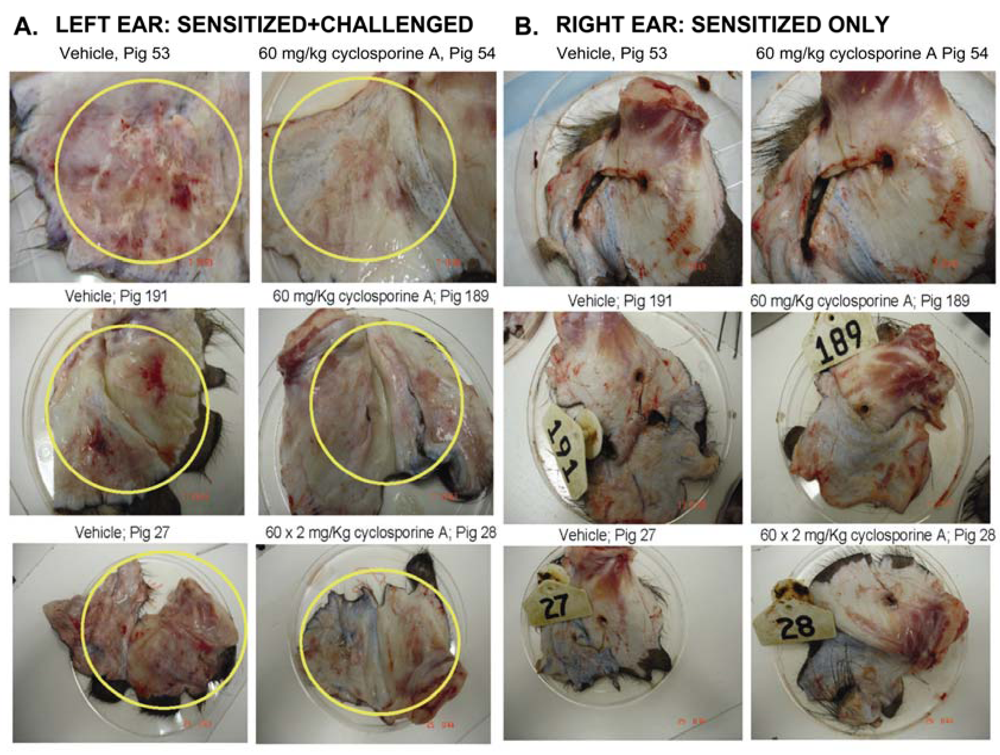
3.1. Effect of CsA on the ear inflammation of pigs in group 1
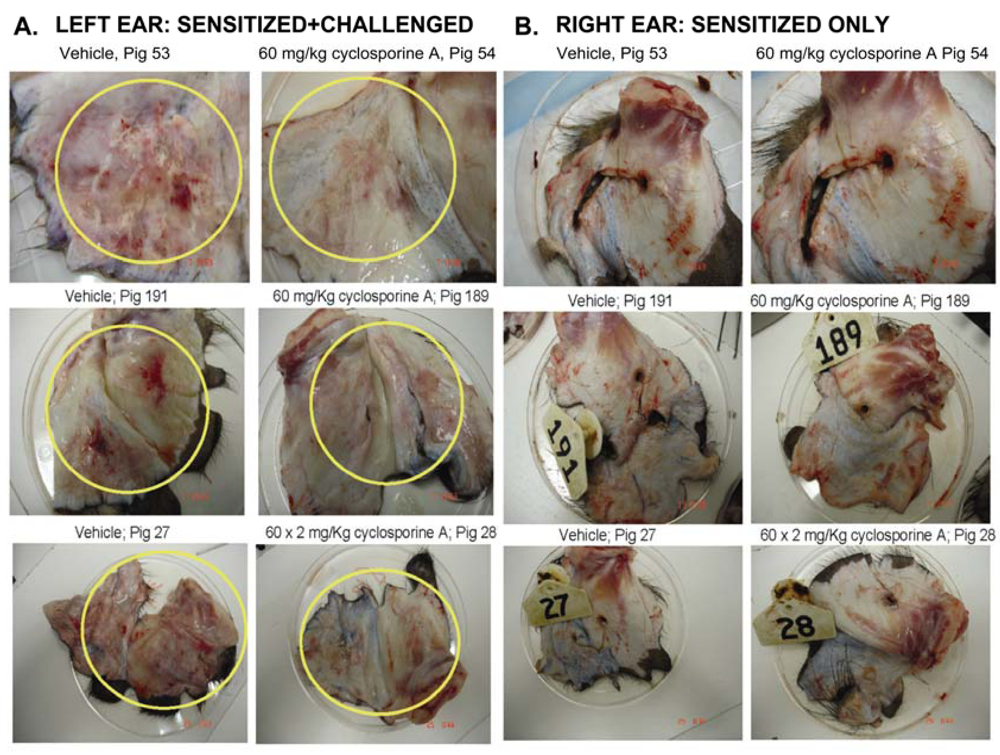
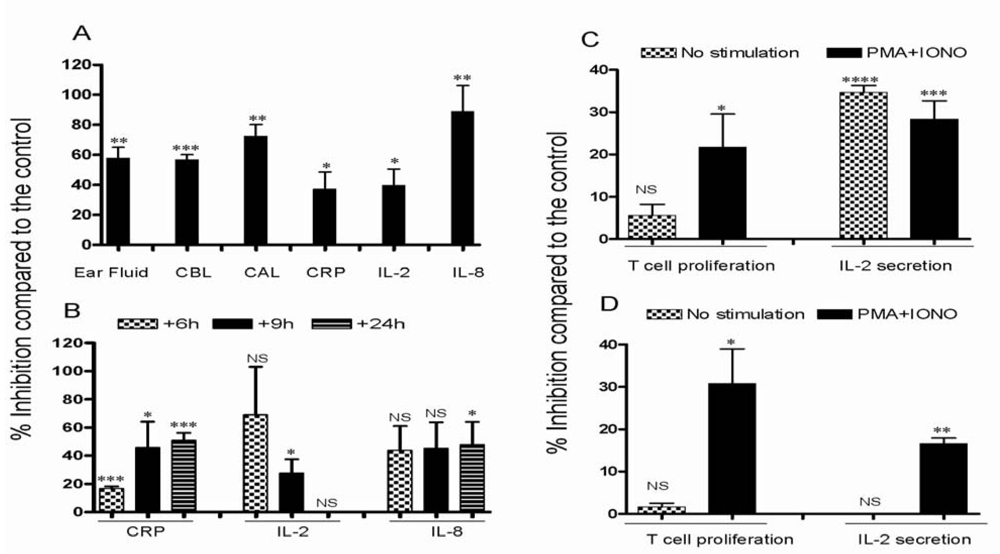
3.2. Effect of CsA on inflammatory reactions in inflamed ears
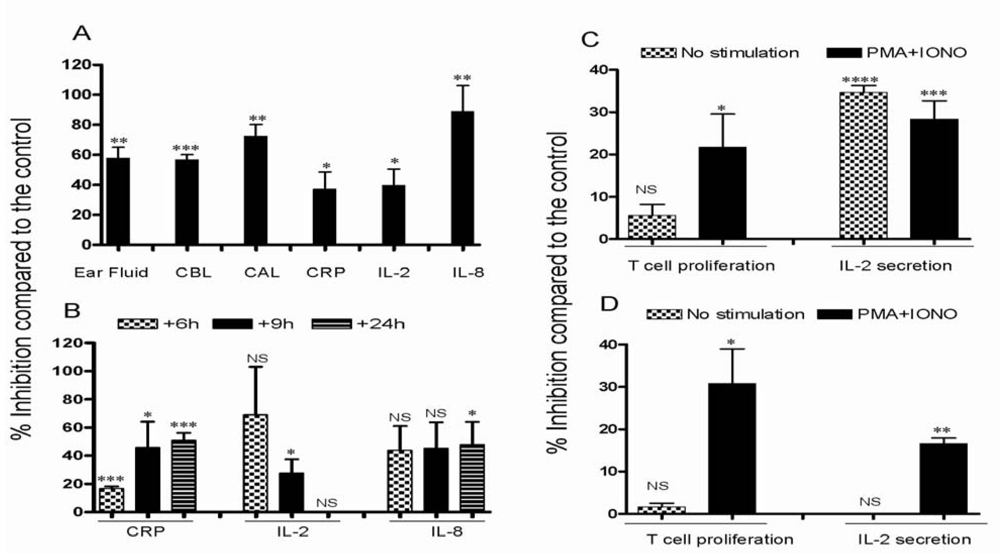
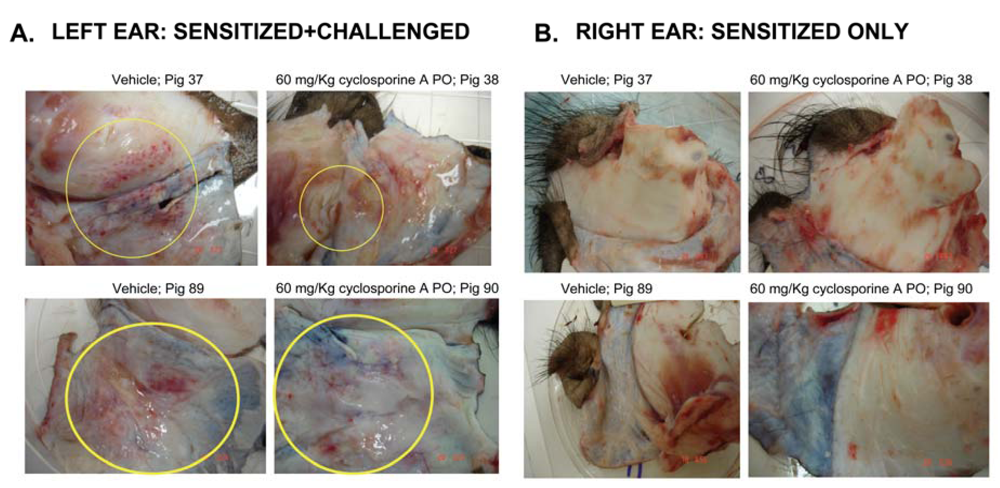
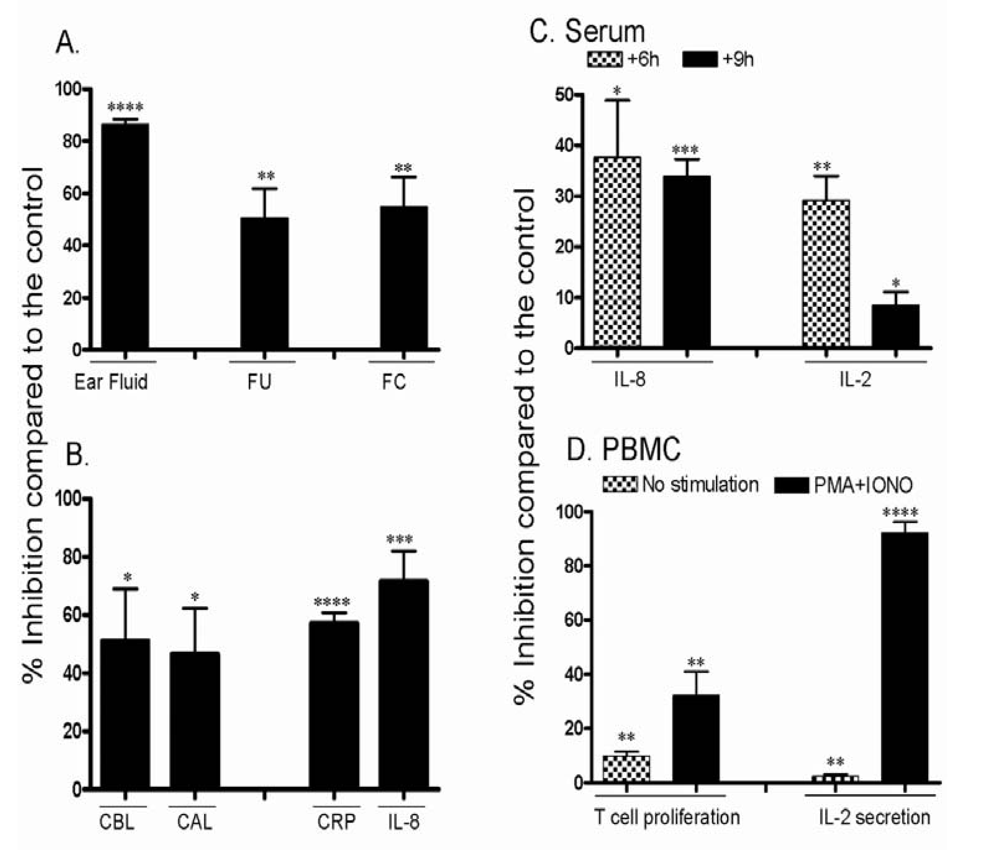
3.3. Effect of CsA on the ear inflammation of pigs in group 2
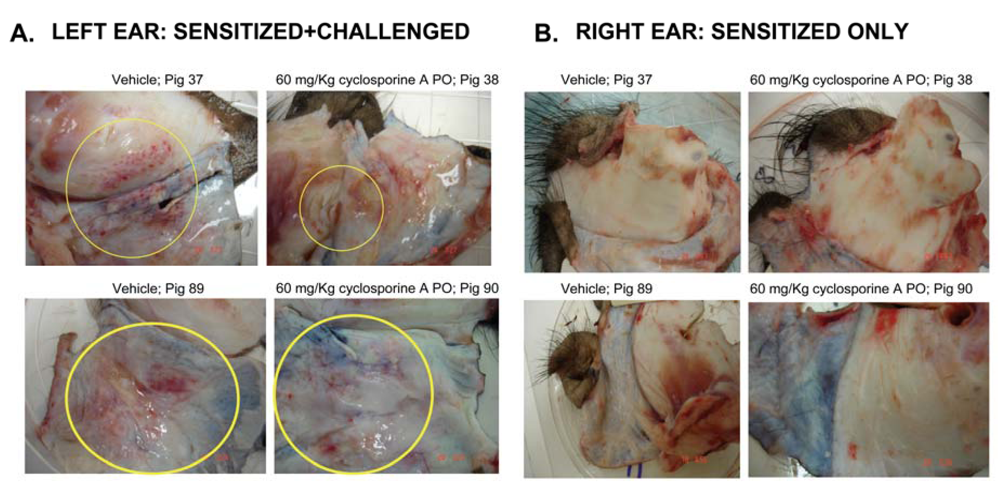
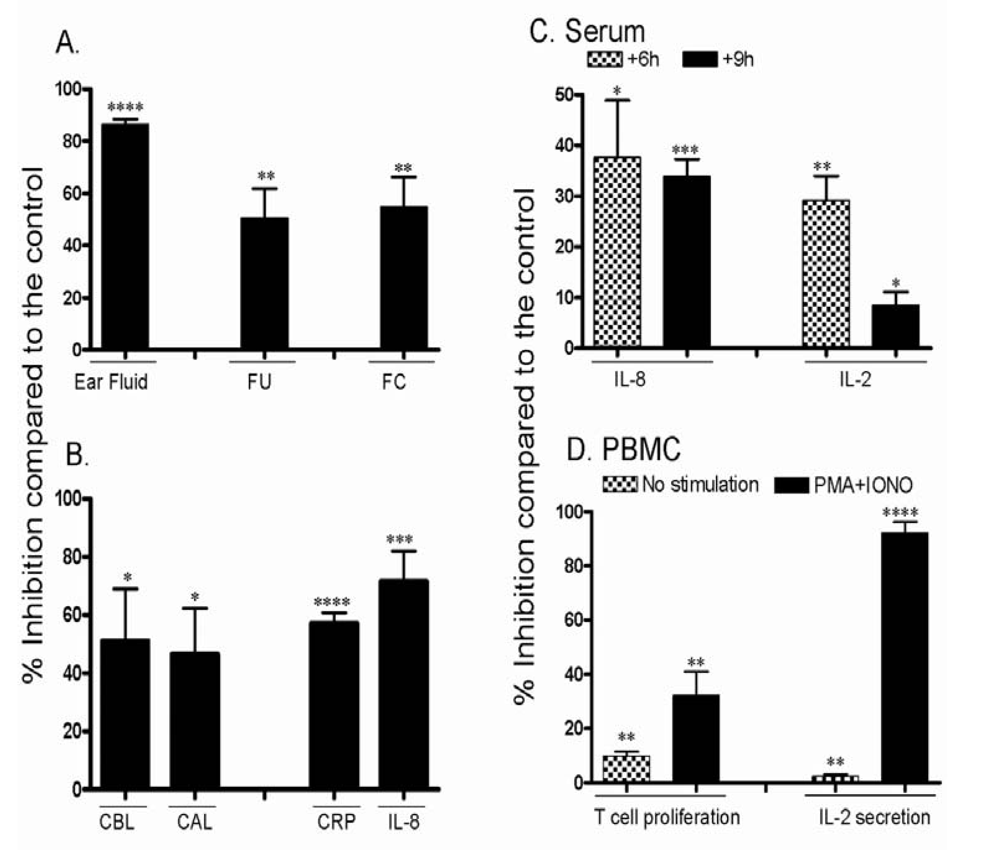
4. Discussion
References
- Di Pierro, F.; d'Atri, G.; Marcucci, F.; Leoni, F. Use of type I and type IV hypersensitivity responses to define the immunopharmacological profile of drugs. J. Pharmacol. Toxicol. Meth. 1997, 37, 91–96. [Google Scholar]
- Todderud, G.; Nair, X.; Lee, D.; Alford, J.; Davern, L.; Stanley, P.; Bachand, C.; Lapointe, P.; Marinier, A.; Martel, A.; Menard, M.; Wright, J.J.; Bajorath, J.; Hollenbaugh, D.; Aruffo, A.; Tramposch, K.M. BMS-190394, a selectin inhibitor, prevents rat cutaneous inflammatory reactions. J. Pharmacol. Exp. Ther. 1997, 282, 1298–1304. [Google Scholar] [PubMed]
- Fukuda, S.; Midoro, K.; Kamei, T.; Gyoten, M.; Kawano, Y.; Ashida, Y.; Nagaya, H. Inhibition of allergic dermal inflammation by the novel imidazopyridazine derivative TAK-427 in a guinea pig experimental model of eczema. J. Pharmacol. Exp. Ther. 2002, 303, 1283–1290. [Google Scholar] [PubMed]
- Zhu, Y.N.; Zhao, W.M.; Yang, Y.F.; Liu, Q.F.; Zhou, Y.; Tian, J.; Ni, J.; Fu, Y. F.; Zhong, X.G.; Tang, W.; Zhou, R.; He, P.L.; Li, X.Y.; Zuo, J.P. Periplocoside E, an effective compound from Periploca sepium Bge, inhibited T cell activation in vitro and in vitro. J. Pharmacol. Exp. Ther. 2006, 316, 662–669. [Google Scholar] [PubMed]
- Särnstrand, B.; Jansson, A.H.; Matuseviciene, G.; Scheynius, A.; Pierrou, S.; Bergstrand, H. N,N'-Diacetyl-L-cystine-the disulfide dimer of N-acetylcysteine-is a potent modulator of contact sensitivity/delayed type hypersensitivity reactions in rodents. J. Pharmacol. Exp. Ther. 1999, 288, 1174–1184. [Google Scholar] [PubMed]
- Saso, Y.; Conner, E.M.; Teegarden, B.R.; Yuan, C.S. S-Adenosyl-L-homocysteine hydrolase inhibitor mediates immunosuppressive effects in vitro: Suppression of delayed type hypersensitivity ear swelling and peptidoglycan polysaccharide-induced arthritis. J. Pharmacol. Exp. Ther. 2001, 296, 106–112. [Google Scholar] [PubMed]
- Cardona, K.; Milas, Z.; Strobert, E.; Cano, J.; Jiang, W.; Safley, S.A.; Gangappa, S.; Hering, B.J.; Weber, C.J.; Pearson, T.C.; Larsen, C.P. Engraftment of adult porcine islet xenografts in diabetic nonhuman primates through targeting of costimulation pathways. Am. J. Transplant. 2007, 7, 2260–2268. [Google Scholar] [PubMed]
- Hermida-Prieto, M.; Domenech, N.; Moscoso, I.; Diaz, T.; Ishii, J.; Salomon, D.R.; Mañez, R. Lack of cross-species transmission of porcine endogenous retrovirus (PERV) to transplant recipients and abattoir workers in contact with pigs. Transplantation 2007, 84, 548–550. [Google Scholar] [PubMed]
- Hoerbelt, R.; Benjamin, L.C.; Shoji, T.; Johnston, D.R.; Muniappan, A.; Guenther, D.A.; Allan, J.S.; Houser, S.L.; Madsen, J.C. Effects of tolerance induction on the actions of interferon-gamma on porcine cardiac allografts. Transplant. Proc. 2006, 38, 3196–3198. [Google Scholar] [PubMed]
- Mezrich, J.D.; Haller, G.W.; Arn, J.S.; Houser, S.L.; Madsen, J.C.; Sachs, D.H. Histocompatible miniature swine: An inbred large-animal model. Transplantation 2003, 75, 904–907. [Google Scholar] [PubMed]
- Wu, G.; Pfeiffer, S.; Schröder, C.; Zhang, T.; Nguyen, B.N.; Kelishadi, S.; Atkinson, J.B.; Schuurman, H.J.; White, D.J.; Azimzadeh, A.M.; Pierson, R.N. Coagulation cascade activation triggers early failure of pig hearts expressing human complement regulatory genes. Xenotransplantation 2007, 14, 34–47. [Google Scholar] [PubMed]
- Fukudo, M.; Yano, I.; Masuda, S.; Okuda, M.; Inui, K. Distinct inhibitory effects of tacrolimus and cyclosporin a on calcineurin phosphatase activity. J. Pharmacol. Exp. Ther. 2005, 312, 816–825. [Google Scholar] [PubMed]
- Mirshahpanah, P.; Li, Y.-Y.Y.; Burkhardt, N.; Asadullah, K.; Zollner, T.M. CCR4 and CCR10 lignads play additive roles in mouse contact hypersensitivity. Exp. Dermatol. 2007, 17, 30–34. [Google Scholar]
- Hioe, K.M.; Jones, J.M. Effects of trimethyltin on the immune system of rats. Toxicol. Lett. 1984, 20, 317–323. [Google Scholar] [PubMed]
- Weiss, L.; Slavin, S.; Reich, S.; Cohen, P.; Shuster, S.; Stern, R.; Kaganovsky, E.; Okon, E.; Rubinstein, A.M.; Naor, D. Induction of resistance to diabetes in non-obese diabetic mice by targeting CD44 with a specific monoclonal antibody. Proc. Natl. Acad. Sci. USA 2000, 97, 285–290. [Google Scholar]
- Grabbe, S.; Schwarz, T. Immunoregulatory mechanisms involved in elicitation of allergic contact hypersensitivity. Immunol. Today 1998, 19, 37–44. [Google Scholar] [PubMed]
- Vana, G.; Meingassner, J.G. Morphologic and immunohistochemical features of experimentally induced allergic contact dermatitis in Göttingen minipigs. Vet. Pathol. 2000, 37, 565–580. [Google Scholar] [PubMed]
- Maghazachi, A.A. Insights into seven and single transmembrane-spanning domain receptors and their signaling pathways in human natural killer cells. Pharmacol. Rev. 2005, 57, 339–357. [Google Scholar] [PubMed]
- Luster, A.D. Chemokines-chemotactic cytokines that mediate inflammation. N. Engl. J. Med. 1998, 338, 436–445. [Google Scholar] [PubMed]
- Reis, F.; Parada, B.; Teixeira, d.L.E.; Garrido, P.; Dias, A.; Piloto, N.; Baptista, S.; Sereno, J.; Eufrásio, P.; Costa, E.; Rocha-Pereira, P.; Santos-Silva, A.; Figueiredo, A.; Mota, A.; Teixeira, F. Hypertension induced by immunosuppressive drugs: A comparative analysis between sirolimus and cyclosporine. Transplant. Proc. 2009, 41, 868–887. [Google Scholar] [PubMed]
- Borel, J.F.; Feurer, C.; Magnée, C.; Stähelin, H. Effects of the new anti-lymphocytic peptide cyclosporin A in animals. Immunology 1977, 32, 1017–1025. [Google Scholar] [PubMed]
- Borel, J.F. Pharmacology of cyclosporine (sandimmune). IV. Pharmacological properties in vitro. Pharmacol. Rev. 1990, 41, 259–371. [Google Scholar] [PubMed]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nuhaily, S.; Damaj, B.B.; Maghazachi, A.A. Oxazolone-Induced Delayed Type Hypersensitivity Reaction in the Adult Yucatan Pigs. A Useful Model for Drug Development and Validation. Toxins 2009, 1, 25-36. https://doi.org/10.3390/toxins1010025
Nuhaily S, Damaj BB, Maghazachi AA. Oxazolone-Induced Delayed Type Hypersensitivity Reaction in the Adult Yucatan Pigs. A Useful Model for Drug Development and Validation. Toxins. 2009; 1(1):25-36. https://doi.org/10.3390/toxins1010025
Chicago/Turabian StyleNuhaily, Samer, Bassam B. Damaj, and Azzam A. Maghazachi. 2009. "Oxazolone-Induced Delayed Type Hypersensitivity Reaction in the Adult Yucatan Pigs. A Useful Model for Drug Development and Validation" Toxins 1, no. 1: 25-36. https://doi.org/10.3390/toxins1010025
APA StyleNuhaily, S., Damaj, B. B., & Maghazachi, A. A. (2009). Oxazolone-Induced Delayed Type Hypersensitivity Reaction in the Adult Yucatan Pigs. A Useful Model for Drug Development and Validation. Toxins, 1(1), 25-36. https://doi.org/10.3390/toxins1010025