Omega–3 Long-Chain Fatty Acids in the Heart, Kidney, Liver and Plasma Metabolite Profiles of Australian Prime Lambs Supplemented with Pelleted Canola and Flaxseed Oils

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location and Animals
2.2. Experimental Design and Diets
2.3. Blood and Visceral Organ Sampling
2.4. Feed Chemical Analysis
2.5. Fatty Acid and Plasma Metabolite Analyses
2.6. Statistical Analysis
3. Results
3.1. Feed Ingredients, Chemical Compositions and Fatty Acid Profiles
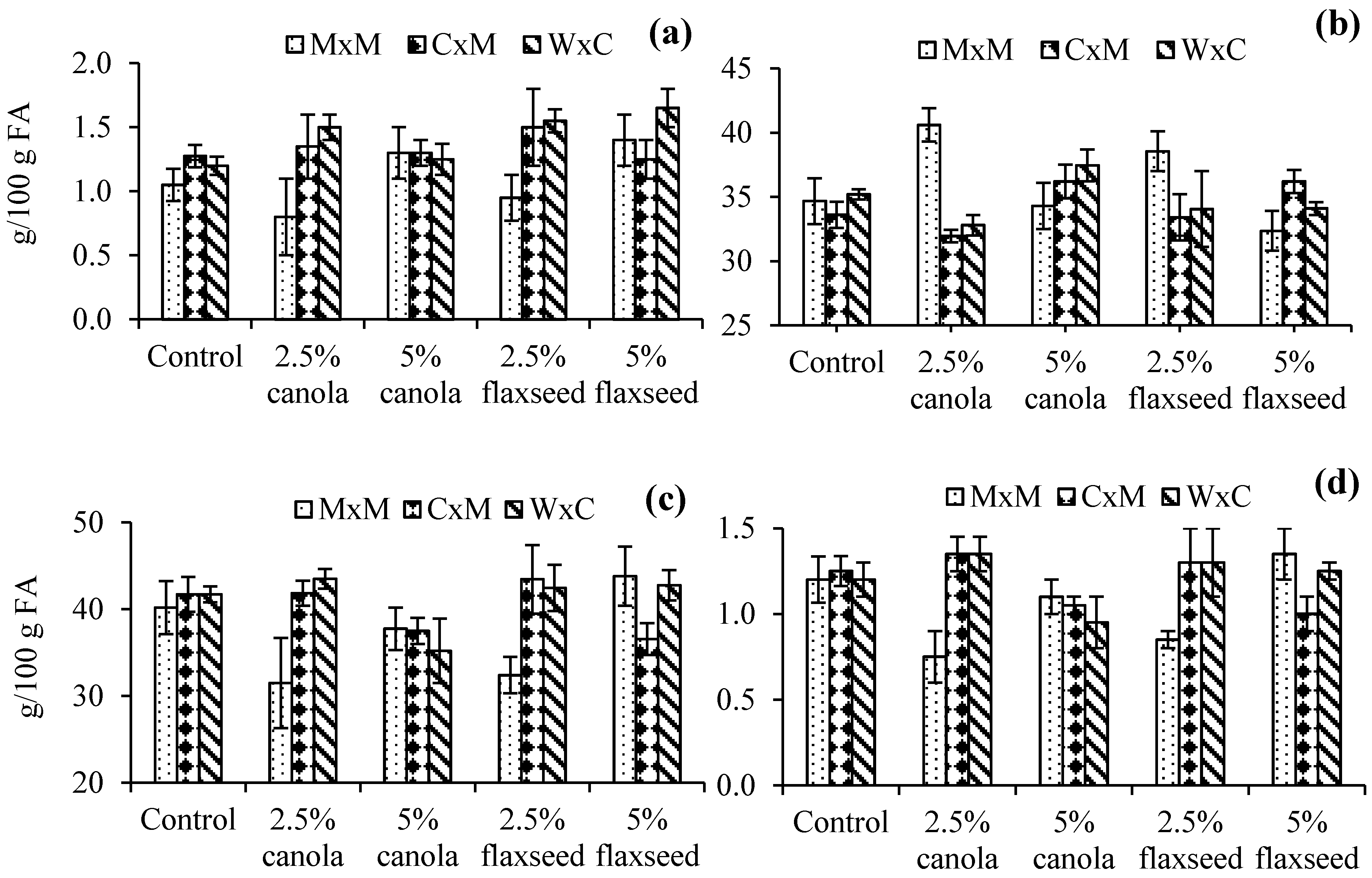
3.2. Heart Fatty Acid Profile
3.3. Liver Fatty Acid Profile
3.4. Kidney Fatty Acid Profile
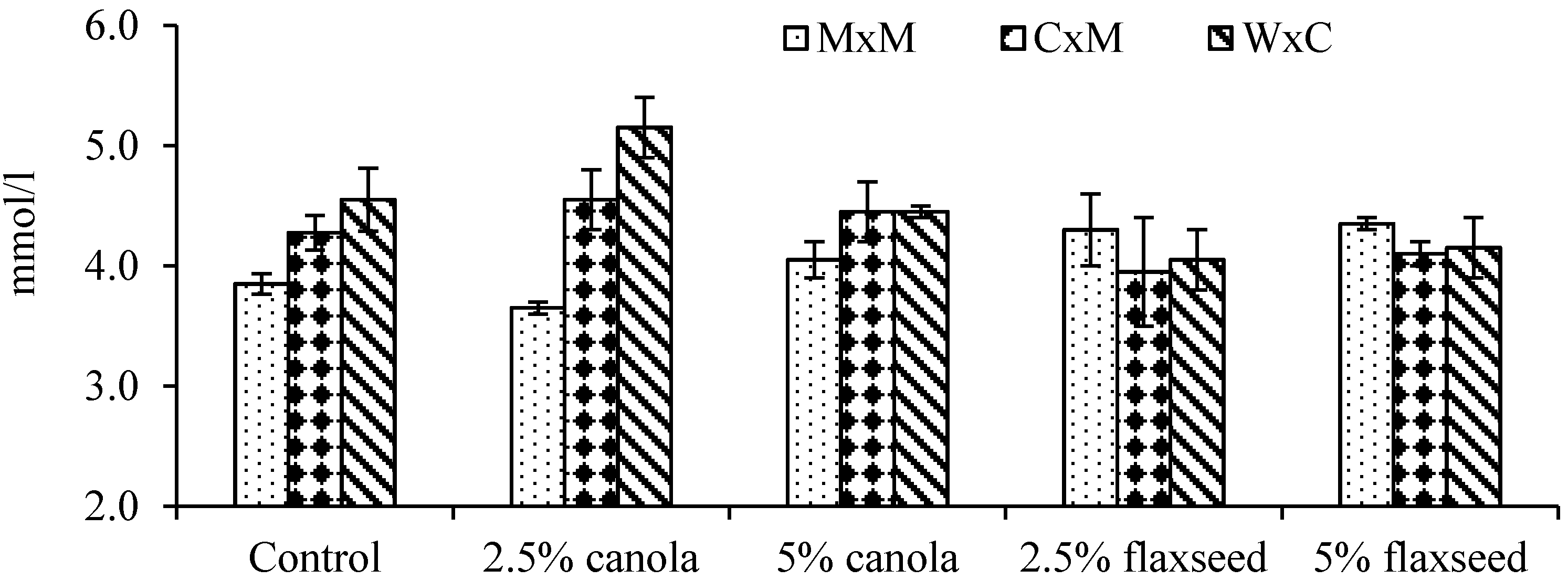
3.5. Plasma Metabolites
4. Discussion
Potential Human Health Implications for Consumption of Lamb Liver and Kidney
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Australian Guide to Healthy Eating. Available online: https://www.eatforhealth.gov.au/guidelines/australian-guide-healthy-eating (accessed on 17 August 2017).
- Ekmekcioglu, C.; Wallner, P.; Kundi, M.; Weisz, U.; Haas, W.; Hutter, H.-P. Red meat, diseases and healthy alternatives: A critical review. Crit. Rev. Food Sci. Nutr. 2016. [Google Scholar] [CrossRef] [PubMed]
- Richi, E.B.; Baumer, B.; Conrad, B.; Darioli, R.; Schmid, A.; Keller, U. Health risks associated with meat consumption: A review of epidemiological studies. Int. J. Vitam. Nutr. Res. 2015, 85, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Enser, M.; Hallett, K.; Hewitt, B.; Fursey, G.; Wood, J. Fatty acid content and composition of english beef, lamb and pork at retail. Meat Sci. 1996, 42, 443–456. [Google Scholar] [CrossRef]
- Adeyemi, K.D.; Sabow, A.B.; Ebrahimi, M.; Samsudin, A.A.; Sazili, A.Q. Fatty acid composition, cholesterol and antioxidant status of infraspinatus muscle, liver and kidney of goats fed blend of palm oil and canola oil. Ital. J. Anim. Sci. 2016, 15, 181–190. [Google Scholar] [CrossRef]
- De Brito, G.F.; Ponnampalam, E.N.; Hopkins, D.L. The effect of extensive feeding systems on growth rate, carcass traits, and meat quality of finishing lambs. Compr. Rev. Food Sci. Food Saf. 2017, 16, 23–38. [Google Scholar] [CrossRef]
- Alvarenga, T.I.R.C.; Chen, Y.; Furusho-Garcia, I.F.; Perez, J.R.O.; Hopkins, D.L. Manipulation of omega-3 PUFAs in lamb: Phenotypic and genotypic views. Compr. Rev. Food Sci. Food Saf. 2015, 14, 189–204. [Google Scholar] [CrossRef]
- Castro, T.; Cabezas, A.; De la Fuente, J.; Isabel, B.; Manso, T.; Jimeno, V. Animal performance and meat characteristics in steers reared in intensive conditions fed with different vegetable oils. Animal 2016, 10, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Bessa, R.J.; Alves, S.P.; Jerónimo, E.; Alfaia, C.M.; Prates, J.A.; Santos-Silva, J. Effect of lipid supplements on ruminal biohydrogenation intermediates and muscle fatty acids in lambs. Eur. J. Lipid Sci. Technol. 2007, 109, 868–878. [Google Scholar] [CrossRef]
- Ding, S.; Meale, S.; Alazzeh, A.; He, M.; Ribeiro, G.; Jin, L.; Wang, Y.; Dugan, M.; Chaves, A.; McAllister, T. Effect of Propionibacterium freudenreichii in diets containing rapeseed or flaxseed oil on in vitro ruminal fermentation, methane production and fatty acid biohydrogenation. Anim. Prod. Sci. 2016. [Google Scholar] [CrossRef]
- Gillingham, L.G.; Gustafson, J.A.; Han, S.-Y.; Jassal, D.S.; Jones, P.J. High-oleic rapeseed (canola) and flaxseed oils modulate serum lipids and inflammatory biomarkers in hypercholesterolaemic subjects. Br. J. Nutr. 2011, 105, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Meale, S.J.; Chaves, A.V.; He, M.L.; Guan, L.L.; McAllister, T.A. Effects of various dietary lipid additives on lamb performance, carcass characteristics, adipose tissue fatty acid composition, and wool characteristics. J. Anim. Sci. 2015, 93, 3110–3120. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, O.; Mendizabal, J.; Insausti, K.; Soret, B.; Purroy, A.; Arana, A. Effect of linseed dietary supplementation on adipose tissue development, fatty acid composition, and lipogenic gene expression in lambs. Livest. Sci. 2015, 178, 345–356. [Google Scholar] [CrossRef]
- Francisco, A.; Alves, S.; Portugal, P.; Pires, V.; Dentinho, M.; Alfaia, C.; Jerónimo, E.; Prates, J.; Santos-Silva, J.; Bessa, R. Effect of feeding lambs with a tanniferous shrub (rockrose) and a vegetable oil blend on fatty acid composition of meat lipids. Animal 2016, 10, 2061–2073. [Google Scholar] [CrossRef] [PubMed]
- Flakemore, A.R.; Balogun, R.O.; McEvoy, P.D.; Malau-Aduli, B.S.; Nichols, P.; Malau-Aduli, A.E.O. Genetic variation in intramuscular fat of prime lambs supplemented with varying concentrations of degummed crude canola oil. Int. J. Nutr. Food Sci. 2014, 3, 203–209. [Google Scholar]
- Annett, R.; Carson, A.; Dawson, L.; Kilpatrick, D. Effects of dam breed and dietary source of n-3 polyunsaturated fatty acids on the growth and carcass characteristics of lambs sourced from hill sheep flocks. Animal 2011, 5, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Demirel, G.; Ozpinar, H.; Nazli, B.; Keser, O. Fatty acids of lamb meat from two breeds fed different forage: Concentrate ratio. Meat Sci. 2006, 72, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Wachira, A.; Sinclair, L.; Wilkinson, R.; Enser, M.; Wood, J.; Fisher, A. Effects of dietary fat source and breed on the carcass composition, n-3 polyunsaturated fatty acid and conjugated linoleic acid content of sheep meat and adipose tissue. Br. J. Nutr. 2002, 88, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F.; Aristoy, M.-C.; Mora, L.; Reig, M. Innovations in value-addition of edible meat by-products. Meat Sci. 2012, 92, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Umaraw, P.; Pathak, V.; Rajkumar, V.; Verma, A.K.; Singh, V.; Verma, A.K. Assessment of fatty acid and mineral profile of Barbari kid in longissimus lumborum muscle and edible byproducts. Small Ruminant Res. 2015, 132, 147–152. [Google Scholar] [CrossRef]
- Rule, D.; Busboom, J.; Kercher, C. Effect of dietary canola on fatty acid composition of bovine adipose tissue, muscle, kidney, and liver. J. Anim. Sci. 1994, 72, 2735–2744. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Adesogan, A.; Badinga, L.; Staples, C. Effects of dietary n-6: n-3 fatty acid ratio on feed intake, digestibility, and fatty acid profiles of the ruminal contents, liver, and muscle of growing lambs. J. Anim. Sci. 2007, 85, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.P.; Trumel, C.; Bézille, P. Clinical biochemistry in sheep: A selected review. Small Ruminant Res. 2010, 92, 10–18. [Google Scholar] [CrossRef]
- Hegarty, R.S.; McFarlane, J.R.; Banks, R.; Harden, S. Association of plasma metabolites and hormones with the growth and composition of lambs as affected by nutrition and sire genetics. Aust. J. Agric. Res. 2006, 57, 683–690. [Google Scholar] [CrossRef]
- Mertens, D.R. Gravimetric determination of amylase-treated neutral detergent fiber in feeds with refluxing in beakers or crucibles: Collaborative study. J. AOAC Int. 2002, 85, 1217–1240. [Google Scholar] [PubMed]
- Garnsworthy, P.; Unal, Y. Estimation of dry-matter intake and digestibility in group-fed dairy cows using near infrared reflectance spectroscopy. Anim. Sci. 2004, 79, 327–334. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; Holman, B.W.B.; Kashani, A.; Nichols, P.D. Sire breed and sex effects on the fatty acid composition and content of heart, kidney, liver, adipose and muscle tissues of purebred and first-cross prime lambs. Anim. Prod. Sci. 2016, 56, 2122–2132. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Clayton, E.H. Graham Centre Monograph No. 4: Long-Chain Omega-3 Polyunsaturated Fatty Acids in Ruminant Nutrition: Benefits to Animals and Humans; Nugent, T., Nicholls, C., Eds.; NSW Department of Primary Industries: Wagga, Australia, 2014.
- Anderson, B.; Kinsella, J.; Watt, B. Comprehensive evaluation of fatty acids in foods. II. Beef products. J. Am. Diet. Assoc. 1975, 67, 35–41. [Google Scholar] [PubMed]
- Food Standards in Australia and New Zealand. Nutrition Information User Guide to Standard 1.2.8—Nutrition Information Requirements Part B—Nutrition Claims. Canberra, Australia, 2012. Available online: http://www.foodstandards.gov.au/code/userguide/Documents/Userguide_Nutrition%20Claims_PartB_March12.pdf (accessed on 15 December 2016).
- Ponnampalam, E.; Butler, K.; Pearce, K.; Mortimer, S.; Pethick, D.; Ball, A.; Hopkins, D. Sources of variation of health claimable long chain omega-3 fatty acids in meat from Australian lambs slaughtered at similar weights. Meat Sci. 2014, 96, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, D.; Clayton, E.; Lamb, T.; van de Ven, R.; Refshauge, G.; Kerr, M.; Bailes, K.; Lewandowski, P.; Ponnampalam, E. The impact of supplementing lambs with algae on growth, meat traits and oxidative status. Meat Sci. 2014, 98, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Nichols, P.D.; Petrie, J.; Singh, S. Long-chain omega-3 oils–An update on sustainable sources. Nutrients 2010, 2, 572–585. [Google Scholar] [CrossRef] [PubMed]
- Byelashov, O.A.; Sinclair, A.J.; Kaur, G. Dietary sources, current intakes, and nutritional role of omega-3 docosapentaenoic acid. Lipid Technol. 2015, 27, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Lemaitre, R.N.; King, I.B.; Song, X.; Huang, H.; Sacks, F.M.; Rimm, E.B.; Wang, M.; Siscovick, D.S. Plasma phospholipid long-chain ω-3 fatty acids and total and cause-specific mortality in older adults: A cohort study. Ann. Intern. Med. 2013, 158, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, Y.; Liang, Y.; Tian, X.; Peng, C.; Ma, K.Y.; Liu, J.; Huang, Y.; Chen, Z.-Y. DPA n-3, DPA n-6 and DHA improve lipoprotein profiles and aortic function in hamsters fed a high cholesterol diet. Atherosclerosis 2012, 221, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Howe, P.; Buckley, J.; Meyer, B. Long-chain omega-3 fatty acids in red meat. Nutr. Diet. 2007, 64, 135–139. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Holman, B.W.; Kerry, J.P. The impact of animal nutrition on muscle composition and meat quality. In Meat Quality. Genetic and Environmental Factors; CRC Press: Boca Raton, FL, USA, 2016; pp. 101–146. [Google Scholar]
- Rahmawaty, S.; Charlton, K.; Lyons-Wall, P.; Meyer, B.J. Dietary intake and food sources of EPA, DPA and DHA in Australian children. Lipids 2013, 48, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Howe, P.; Meyer, B.; Record, S.; Baghurst, K. Dietary intake of long-chain ω-3 polyunsaturated fatty acids: Contribution of meat sources. Nutrition 2006, 22, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Wu, J.H. (n-3) Fatty acids and cardiovascular health: Are effects of EPA and DHA shared or complementary? J. Nutr. 2012, 142, 614S–625S. [Google Scholar] [CrossRef] [PubMed]
- National Health and Medical Research Council. Nutrient Reference Values for Australia and New Zealand Fats: Total Fat & Fatty Acids. Canberra, Australia, 2006. Available online: https://www.nrv.gov.au/sites/default/files/page_pdf/n35-fat_0.pdf (accessed on 15 December 2016).
- Bessa, R.J.; Alves, S.P.; Santos-Silva, J. Constraints and potentials for the nutritional modulation of the fatty acid composition of ruminant meat. Eur. J. Lipid Sci. Technol. 2015, 117, 1325–1344. [Google Scholar] [CrossRef]
- Nguyen, D.V.; Flakemore, A.R.; Otto, J.R.; Ives, S.W.; Smith, R.W.; Nichols, P.D.; Malau-Aduli, A.E.O. Nutritional value and sensory characteristics of meat eating quality of australian prime lambs supplemented with pelleted canola and flaxseed oils: Fatty acid profiles of muscle and adipose tissues. Intern. Med. Rev. 2017, 3, 1–21. [Google Scholar]
- Jerónimo, E.; Alves, S.P.; Martins, S.V.; Prates, J.A.; Bessa, R.J.; Santos-Silva, J. Effect of sodium bentonite and vegetable oil blend supplementation on growth, carcass quality and intramuscular fatty acid composition of lambs. Anim. Feed Sci. Technol. 2010, 158, 136–145. [Google Scholar] [CrossRef]
- Kashani, A.; Holman, B.W.B.; Nichols, P.D.; Malau-Aduli, A.E.O. Effect of level of spirulina supplementation on the fatty acid compositions of adipose, muscle, heart, kidney and liver tissues in Australian dual-purpose lambs. Ann. Anim. Sci. 2015, 15, 945–960. [Google Scholar] [CrossRef]
- Demirel, G.; Wachira, A.; Sinclair, L.; Wilkinson, R.; Wood, J.; Enser, M. Effects of dietary n-3 polyunsaturated fatty acids, breed and dietary vitamin E on the fatty acids of lamb muscle, liver and adipose tissue. Br. J. Nutr. 2004, 91, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Wilde, P.; Dawson, R. The biohydrogenation of α-linoleic acid and oleic acid by rumen micro-organisms. Biochem. J. 1966, 98, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Doreau, M.; Ferlay, A. Digestion and utilisation of fatty acids by ruminants. Anim. Feed Sci. Technol. 1994, 45, 379–396. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; Edriss, M.A.; Siebert, B.D.; Bottema, C.D.K.; Pitchford, W.S. Breed differences and heterosis in triacylglycerol fatty acid composition of bovine adipose tissue. J. Anim. Physiol. Anim. Nutr. 2000, 83, 106–112. [Google Scholar] [CrossRef]
- Sanudo, C.; Enser, M.; Campo, M.; Nute, G.; Marıa, G.; Sierra, I.; Wood, J. Fatty acid composition and sensory characteristics of lamb carcasses from Britain and Spain. Meat Sci. 2000, 54, 339–346. [Google Scholar] [CrossRef]
- Hoffman, L.; Laubscher, L.; Leisegang, K. Nutritional value of cooked offal derived from free-range rams reared in South Africa. Meat Sci. 2013, 93, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Maranesi, M.; Bochicchio, D.; Montellato, L.; Zaghini, A.; Pagliuca, G.; Badiani, A. Effect of microwave cooking or broiling on selected nutrient contents, fatty acid patterns and true retention values in separable lean from lamb rib-loins, with emphasis on conjugated linoleic acid. Food Chem. 2005, 90, 207–218. [Google Scholar] [CrossRef]
- Flakemore, A.R.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Omega-3 fatty acids, nutrient retention values, and sensory meat eating quality in cooked and raw Australian lamb. Meat Sci. 2017, 123, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, L. Nutritional manipulation of the fatty acid composition of sheep meat: A review. J. Agric. Sci. 2007, 145, 419–434. [Google Scholar] [CrossRef]
- Obeidat, B.S.; Awawdeh, M.S.; Telfah, B.T.; Ballou, M.A. Calcium salts of long chain fatty acids in Awassi ewe lambs’ diet: Effects on nutrient intake, digestibility, and blood metabolites. Livest. Sci. 2012, 150, 391–396. [Google Scholar] [CrossRef]
- Bhatt, R.S.; Karim, S.A.; Sahoo, A.; Shinde, A.K. Growth performance of lambs fed diet supplemented with rice bran oil as such or as calcium soap. Asian-Australas. J. Anim. Sci. 2013, 26, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Mao, S.; Zhu, W. 17. A 22: 6 n-3 rich supplement affects the ruminal microbial community and fermentation and alters plasma metabolites. Ann. Anim. Sci. 2016, 16, 533–550. [Google Scholar] [CrossRef]
- Mirzaei, F.; Rezaeian, M.; Towhidi, A.; Nik-Khah, A.; Sereshti, H. Effects of fish oil, safflower oil and monensin supplementation on performance, rumen fermentation parameters and plasma metabolites in chall sheep. Int. J. Vet. Res. 2009, 3, 113–128. [Google Scholar]
- Otto, J.R.; Malau-Aduli, B.S.; Balogun, R.O.; Nish, P.; Malau-Aduli, A.E.O. Effect of crude degummed canola oil and ad libitum grazing on plasma metabolites of primiparous Holstein-Friesian cows in a pasture-based system. BMC Vet. Res. 2014, 10, 224. [Google Scholar] [CrossRef] [PubMed]
- Wachira, A.; Sinclair, L.; Wilkinson, R.; Hallett, K.; Enser, M.; Wood, J. Rumen biohydrogenation of n-3 polyunsaturated fatty acids and their effects on microbial efficiency and nutrient digestibility in sheep. J. Agric. Sci. 2000, 135, 419–428. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; Holman, B.W.B. Effect of spirulina supplementation on plasma metabolites in crossbred and purebred Australian Merino lambs. Int. J. Vet. Sci. Med. 2015, 3, 13–20. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; McEvoy, P.D.; Parsons, D.; Lane, P.A. Breed variation in wool quality, growth and plasma metabolites of prime lamb fed degummed canola. In Proceedings of the 10th World Congress of Genetics Applied to Livestock Production, Vancouver, BC, Canada, 17–22 August 2014; pp. 1–6. [Google Scholar]
- Stahlhut, H.; Whisnant, C.; Lloyd, K.; Baird, E.; Legleiter, L.; Hansen, S.; Spears, J. Effect of chromium supplementation and copper status on glucose and lipid metabolism in angus and simmental beef cows. Anim. Feed Sci. Technol. 2006, 128, 253–265. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Stanley, D.F.; Martin, L.C.; Gilmour, A.R. Genotype and age effects on sheep meat production 1. Production and growth. Aust. J. Exp. Agric. 2007, 47, 1119–1127. [Google Scholar] [CrossRef]
- Butler, W. Nutritional interactions with reproductive performance in dairy cattle. Anim. Reprod. Sci. 2000, 60, 449–457. [Google Scholar] [CrossRef]
- Sunny, N.; Owens, S.; Baldwin, R.; El-Kadi, S.; Kohn, R.; Bequette, B. Salvage of blood urea nitrogen in sheep is highly dependent on plasma urea concentration and the efficiency of capture within the digestive tract. J. Anim. Sci. 2007, 85, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Guo, X.-F.; Sinclair, A.J. Short update on docosapentaenoic acid: a bioactive long-chain n-3 fatty acid. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Phang, M.; Garg, M.L.; Sinclair, A.J. Inhibition of platelet aggregation by omega-3 polyunsaturated fatty acids is gender specific—Redefining platelet response to fish oils. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.-N.; Gladman, S.J.; Dyall, S.C.; Patel, U.; Virani, N.; Kang, J.X.; Priestley, J.V.; Michael-Titus, A.T. Transgenic mice with high endogenous omega-3 fatty acids are protected from spinal cord injury. Neurobiol. Dis. 2013, 51, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.; Kaur, G.; Larsen, A.; Loh, S.P.; Linderborg, K.; Weisinger, H.S.; Turchini, G.M.; Cameron-Smith, D.; Sinclair, A.J. A short-term n-3 DPA supplementation study in humans. Eur. J. Nutr. 2013, 52, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Howes, N.L.; Bekhit, A.E.D.A.; Burritt, D.J.; Campbell, A.W. Opportunities and implications of pasture-based lamb fattening to enhance the long-chain fatty acid composition in meat. Compr. Rev. Food Sci. Food Saf. 2015, 14, 22–36. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Item | Pellet Feed 1 | Lucerne Hay | ||||
|---|---|---|---|---|---|---|
| Control | 2.5C | 5C | 2.5F | 5F | ||
| Ingredients (g/kg) | ||||||
| Wheat | 513 | 537 | 545 | 551 | 465 | ‒ |
| Paddy rice | 260 | 230 | 210 | 220 | 280 | ‒ |
| Lupins | 170 | 151 | 138 | 147 | 148 | ‒ |
| Canola oil (mL/kg) | - | 25 | 50 | - | - | ‒ |
| Flaxseed oil (mL/kg) | - | - | - | 25 | 50 | ‒ |
| Salt | 10 | 10 | 10 | 10 | 10 | ‒ |
| Limestone | 21 | 21 | 21 | 21 | 21 | ‒ |
| Sheep premix | 1 | 1 | 1 | 1 | 1 | ‒ |
| Ammonium sulfate | 12.6 | 12.6 | 12.6 | 12.6 | 12.6 | ‒ |
| Acid buff | 6.2 | 6.2 | 6.2 | 6.2 | 6.2 | ‒ |
| Sodium bicarbonate | 6.2 | 6.2 | 6.2 | 6.2 | 6.2 | ‒ |
| Chemical compositions (% dry matter) 2 | ||||||
| Dry matter, (%) | 89.8 | 90.2 | 87.9 | 90.5 | 89.4 | 89.6 |
| Crude protein | 14.7 | 14.5 | 14.4 | 14.5 | 14.5 | 17.4 |
| NDF | 23.8 | 23.5 | 23.9 | 23.7 | 23.3 | 46.5 |
| ADF | 9.2 | 9.3 | 8.9 | 9.5 | 9.0 | 30.9 |
| NFC | 50.5 | 49.9 | 47.8 | 50.5 | 50.7 | 27.4 |
| Ether extract | 3.0 | 4.6 | 5.7 | 4.2 | 5.1 | 2.4 |
| Ash | 8.0 | 7.5 | 8.2 | 7.1 | 6.4 | 7.2 |
| ME (MJ/kgDM) | 10.7 | 10.9 | 11.1 | 10.8 | 11.1 | 9.8 |
| Fatty acid percentage (g/100 g total FA) 3 | ||||||
| 14:0 | 0.2 | 0.5 | 0.6 | 0.2 | 0.2 | 0.6 |
| 15:0 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.4 |
| 16:0 | 18.2 | 16.9 | 16.5 | 19.1 | 19.8 | 29.6 |
| 17:0 | 0.1 | 0.1 | 0.2 | 0.1 | 0.1 | 0.7 |
| 18:2n-6 | 43.4 | 28.4 | 26.7 | 25.6 | 24.7 | 19.1 |
| 18:3n-3 | 3.5 | 3.6 | 4.3 | 4.9 | 7.2 | 18.8 |
| 18:1n-9 | 23.9 | 38.9 | 37.5 | 32.3 | 34.1 | 5.6 |
| 18:0 | 3.4 | 4.1 | 4.1 | 4.4 | 5.1 | 4.7 |
| 20:3n-6 | 0.3 | 0.4 | 0.4 | 0.4 | 0.5 | 0.4 |
| 20:4n-3 | 0.4 | 0.5 | 0.2 | 0.5 | 0.6 | 0.5 |
| 20:2n-6 | 0.1 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 |
| 20:0 | 0.5 | 0.8 | 0.7 | 0.7 | 0.8 | 1.5 |
| ∑SFA | 24.1 | 23.0 | 25.0 | 26.7 | 28.7 | 47.3 |
| ∑MUFA | 27.5 | 42.6 | 43.3 | 36.3 | 37.5 | 12.7 |
| ∑PUFA | 48.4 | 34.4 | 31.7 | 37.0 | 33.8 | 40.0 |
| PUFA/SFA | 2.0 | 1.5 | 1.3 | 1.4 | 1.2 | 0.9 |
| ∑n-3 PUFA | 3.9 | 4.1 | 4.8 | 5.5 | 7.9 | 20.5 |
| ∑n-6 PUFA | 43.8 | 28.8 | 27.4 | 26 | 25.3 | 19.4 |
| n-6/n-3 | 11.1 | 7.0 | 5.7 | 4.7 | 3.2 | 0.9 |
| Item 1 | Treatment | Breed 2 | SEM 3 | p Value 4 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 2.5C | 5C | 2.5F | 5F | M × M | C × M | W × C | T | B | T × B | ||
| Total lipid (g fat/100 g wet tissue) | 2.6 | 2.7 | 2.8 | 2.9 | 2.4 | 2.8 | 2.7 | 2.5 | 0.09 | 0.35 | 0.23 | 0.16 |
| Percentage (g/100 g total FA) | ||||||||||||
| 14:0 | 0.6 | 0.6 | 0.8 | 0.6 | 0.5 | 0.6 | 0.7 | 0.5 | 0.05 | 0.68 | 0.33 | 0.24 |
| 15:0 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.01 | 0.36 | 0.94 | 0.30 |
| 16:0 | 12.9 | 12.5 | 13.1 | 12.9 | 12.7 | 12.9 | 12.9 | 12.7 | 0.22 | 0.93 | 0.63 | 0.06 |
| 17:0 | 1.1 a | 1.0 a,b | 1.0 a,b | 0.9 a,b | 0.8 b | 1.1 a | 1.0 a,b | 0.9 b | 0.03 | 0.04 | 0.02 | 0.07 |
| 18:2n-6 | 23.8 | 21.0 | 21.0 | 21.9 | 22.4 | 20.9 | 23.1 | 22.9 | 0.48 | 0.11 | 0.06 | 0.07 |
| 18:3n-3 | 2.0 | 1.8 | 2.5 | 2.1 | 2.7 | 2.5 | 2.1 | 1.7 | 0.18 | 0.53 | 0.24 | 0.56 |
| 18:1n-9 | 13.4 | 15.5 | 15.6 | 14.2 | 12.8 | 14.9 | 14.7 | 12.9 | 0.45 | 0.17 | 0.08 | 0.39 |
| 18:0 | 17.2 | 18.4 | 18.4 | 17.9 | 17.5 | 18.3 | 17.3 | 17.6 | 0.27 | 0.35 | 0.12 | 0.06 |
| 20:4n-6 (ARA) | 5.6 | 6.2 | 4.3 | 5.8 | 5.2 | 5.1 | 4.8 | 6.5 | 0.33 | 0.33 | 0.06 | 0.06 |
| 20:5n-3 (EPA) | 1.2 | 1.3 | 1.2 | 1.3 | 1.4 | 1.2 | 1.3 | 1.3 | 0.04 | 0.35 | 0.33 | 0.07 |
| 20:3n-6 | 0.5 | 0.4 | 0.4 | 0.4 | 0.5 | 0.4 | 0.4 | 0.5 | 0.01 | 0.22 | 0.37 | 0.44 |
| 20:4n-3 | 1.4 | 1.4 | 1.5 | 1.6 | 1.7 | 1.4 | 1.5 | 1.6 | 0.06 | 0.65 | 0.47 | 0.49 |
| 20:2n-6 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.01 | 0.81 | 0.77 | 0.91 |
| 20:0 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.01 | 0.99 | 0.74 | 0.87 |
| 22:5n-6 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.01 | 0.25 | 0.23 | 0.23 |
| 22:6n-3 (DHA) | 0.6 | 0.7 | 0.7 | 0.7 | 0.7 | 0.7 | 0.6 | 0.7 | 0.03 | 0.85 | 0.56 | 0.72 |
| 22:4n-6 | 0.2 | 0.2 | 0.1 | 0.2 | 0.2 | 0.1 | 0.1 | 0.2 | 0.01 | 0.72 | 0.20 | 0.57 |
| 22:5n-3 (DPA) | 1.1 | 1.1 | 1.2 | 1.3 | 1.4 | 1.2 b | 1.3 a,b | 1.4 a | 0.05 | 0.19 | 0.04 | 0.02 |
| 22:0 | 0.3 | 0.3 | 0.2 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.01 | 0.74 | 0.22 | 0.83 |
| 23:0 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.01 | 0.37 | 0.56 | 0.74 |
| 24:0 | 0.3 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.01 | 0.85 | 0.46 | 0.67 |
| ∑SFA | 34.5 | 35.1 | 35.9 | 35.2 | 34.2 | 35.5 | 34.5 | 34.8 | 0.46 | 0.63 | 0.31 | 0.01 |
| ∑MUFA | 24.3 | 26.3 | 27.3 | 25.3 | 24.8 | 26.7 a | 25.5 a,b | 23.8 b | 0.50 | 0.27 | 0.04 | 0.30 |
| ∑PUFA | 41.2 | 38.6 | 36.8 | 39.5 | 41.0 | 37.8 | 40.0 | 41.4 | 0.87 | 0.30 | 0.08 | 0.03 |
| PUFA/SFA | 1.2 | 1.2 | 1.0 | 1.2 | 1.2 | 1.1 | 1.2 | 1.2 | 0.04 | 0.52 | 0.06 | 0.01 |
| ∑n-3 PUFA | 6.5 | 6.4 | 6.9 | 6.6 | 7.9 | 7.0 | 6.7 | 6.7 | 0.25 | 0.36 | 0.90 | 0.26 |
| ∑n-6 PUFA | 30.3 | 28.1 | 26.1 | 28.6 | 28.5 | 26.9b | 28.7 a,b | 30.3 a | 0.68 | 0.17 | 0.02 | 0.06 |
| n-6/n-3 | 4.8 a | 4.4 a,b | 4.1 a,b | 4.3 a,b | 3.6 b | 4.0 | 4.4 | 4.6 | 0.15 | 0.02 | 0.28 | 0.83 |
| Content (mg/100 g wet tissue) | ||||||||||||
| 18:3n-3 | 24.1 | 26.7 | 24.6 | 27.5 | 31.0 | 31.7 | 18.7 | 26.7 | 2.19 | 0.83 | 0.09 | 0.81 |
| 20:4n-6 | 77.1 | 101.7 | 59.9 | 68.8 | 71.0 | 68.6 | 60.4 | 99.0 | 7.44 | 0.55 | 0.08 | 0.58 |
| EPA | 13.7 | 15.7 | 15.4 | 18 | 20.5 | 16.1 | 13.1 | 20.3 | 1.37 | 0.64 | 0.07 | 0.56 |
| 22:5n-6 | 1.1 | 1.1 | 1.2 | 1.1 | 0.9 | 1.0 | 1.0 | 1.3 | 0.09 | 0.89 | 0.23 | 0.94 |
| DHA | 7.8 | 8.0 | 8.1 | 9.3 | 10.4 | 9.2 | 6.4 | 10.1 | 0.71 | 0.79 | 0.14 | 0.90 |
| DPA | 14.0 | 15.3 | 16.1 | 18.0 | 20.0 | 15.3 | 13.4 | 20.6 | 1.37 | 0.72 | 0.07 | 0.58 |
| EPA + DHA | 21.5 | 23.7 | 23.5 | 27.3 | 30.9 | 25.3 | 19.5 | 30.4 | 2.02 | 0.67 | 0.09 | 0.72 |
| EPA + DHA + DPA | 35.5 | 39.0 | 39.6 | 45.3 | 50.9 | 40.6 | 32.9 | 51.0 | 3.36 | 0.69 | 0.08 | 0.66 |
| Item 1 | Treatment | Breed | SEM | p Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 2.5C | 5C | 2.5F | 5F | M × M | C × M | W × C | T | B | T × B | ||
| Total lipid (g fat/100 g wet tissue) | 6.3 | 6.6 | 6.2 | 6.3 | 6.4 | 6.4 | 6.6 | 6.0 | 0.16 | 0.99 | 0.37 | 0.74 |
| Percentage (g/100 g total FA) 2 | ||||||||||||
| 14:0 | 0.5 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.5 | 0.4 | 0.02 | 0.44 | 0.22 | 0.22 |
| 15:0 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.01 | 0.22 | 0.12 | 0.59 |
| 16:0 | 18.0 a | 17.4 a,b | 17.4 a,b | 17.3 a,b | 15.9 b | 17.2 | 17.3 | 17.2 | 0.28 | 0.03 | 0.85 | 0.47 |
| 17:0 | 1.4 a | 1.3 a,b | 1.3 ab | 1.3 a,b | 1.2 b | 1.4 | 1.3 | 1.3 | 0.03 | 0.02 | 0.24 | 0.56 |
| 18:2n-6 | 10.5 | 10.0 | 10.1 | 9.7 | 10.0 | 10.5 | 9.7 | 10.1 | 0.20 | 0.68 | 0.35 | 0.59 |
| 18:3n-3 | 2.0 | 2.3 | 2.5 | 2.5 | 2.7 | 2.4 | 2.5 | 2.1 | 0.10 | 0.17 | 0.53 | 0.66 |
| 18:1n-9 | 20.7 | 20.5 | 20.7 | 18.1 | 18.8 | 19.9 | 20.2 | 19.6 | 0.44 | 0.30 | 0.88 | 0.61 |
| 18:0 | 18.9 | 19.6 | 19.3 | 21.7 | 20.4 | 19.4 | 19.9 | 20.2 | 0.41 | 0.17 | 0.70 | 0.36 |
| 20:4n-6 (ARA) | 4.2 | 4.6 | 3.1 | 4.4 | 4.5 | 3.7 | 4.7 | 4.0 | 0.33 | 0.73 | 0.57 | 0.77 |
| 20:5n-3 (EPA) | 1.3 | 1.4 | 1.3 | 1.3 | 1.6 | 1.3 | 1.5 | 1.3 | 0.06 | 0.68 | 0.51 | 0.27 |
| 20:3n-6 | 0.7 | 0.7 | 0.7 | 0.6 | 0.6 | 0.7 | 0.7 | 0.7 | 0.03 | 0.50 | 0.86 | 0.07 |
| 20:4n-3 | 0.2 | 0.1 | 0.2 | 0.2 | 0.1 | 0.2 | 0.2 | 0.2 | 0.01 | 0.15 | 0.47 | 0.82 |
| 20:2n-6 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.01 | 0.27 | 0.30 | 0.54 |
| 20:0 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | ||||
| 22:5n-6 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.01 | 0.33 | 0.26 | 0.70 |
| 22:6n-3 (DHA) | 2.4 b | 2.5 b | 2.6 a,b | 2.9 a,b | 3.4 a | 2.9 | 2.6 | 2.5 | 0.13 | 0.03 | 0.50 | 0.18 |
| 22:4n-6 | 0.5 | 0.4 | 0.4 | 0.4 | 0.3 | 0.4 | 0.4 | 0.5 | 0.02 | 0.26 | 0.32 | 0.66 |
| 22:5n-3 (DPA) | 3.3 | 3.1 | 3.4 | 3.4 | 3.7 | 3.1 | 3.3 | 3.6 | 0.12 | 0.64 | 0.24 | 0.47 |
| 22:0 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.01 | 0.62 | 0.99 | 0.92 |
| 23:0 | 0.4 | 0.4 | 0.4 | 0.3 | 0.3 | 0.4 | 0.4 | 0.4 | 0.01 | 0.71 | 0.83 | 0.21 |
| 24:0 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.01 | 0.80 | 0.84 | 0.06 |
| ∑SFA | 40.8 | 41.3 | 41.4 | 44.1 | 40.6 | 41.2 | 41.6 | 41.8 | 0.54 | 0.23 | 0.90 | 0.12 |
| ∑MUFA | 32.6 | 32.0 | 33.1 | 29.2 | 31.1 | 32.1 | 31.5 | 31.6 | 0.54 | 0.29 | 0.91 | 0.68 |
| ∑PUFA | 26.6 | 26.7 | 25.5 | 26.7 | 28.3 | 26.7 | 26.9 | 26.6 | 0.59 | 0.92 | 0.98 | 0.47 |
| PUFA/SFA | 0.7 | 0.6 | 0.6 | 0.6 | 0.7 | 0.6 | 0.6 | 0.6 | 0.07 | 0.69 | 0.74 | 0.21 |
| ∑n-3 PUFA | 9.2 b | 9.6 a,b | 9.7 a,b | 10.3 a,b | 11.5 a | 9.9 | 10.0 | 9.7 | 0.32 | 0.04 | 0.99 | 0.26 |
| ∑n-6 PUFA | 16.5 | 16.3 | 14.8 | 15.6 | 15.9 | 15.9 | 16.0 | 15.9 | 0.40 | 0.73 | 0.96 | 0.67 |
| n-6/n-3 | 1.8 a | 1.7 a,b | 1.6 a,b | 1.6 a,b | 1.4 b | 1.6 | 1.7 | 1.7 | 0.05 | 0.03 | 0.96 | 0.57 |
| Content (mg/100 g wet tissue) | ||||||||||||
| 18:3n-3 | 79.3 | 86.7 | 115.7 | 111.7 | 118.9 | 107.9 | 99.6 | 88.2 | 7.20 | 0.36 | 0.77 | 0.57 |
| 20:4n-6 | 186.0 | 176.6 | 135.6 | 225.2 | 202.5 | 177.0 | 192.6 | 184.2 | 19.89 | 0.85 | 0.96 | 0.88 |
| EPA | 50.9 | 53.5 | 58.2 | 64.8 | 71.6 | 56.9 | 61.2 | 56.0 | 4.73 | 0.86 | 0.90 | 0.59 |
| 22:5n-6 | 9.7 | 9.8 | 9.7 | 7.4 | 8.0 | 10.6 | 7.8 | 8.9 | 0.72 | 0.77 | 0.30 | 0.91 |
| DHA | 97.2 | 99.1 | 108.8 | 142.0 | 146.8 | 128.7 | 110.0 | 106.0 | 9.23 | 0.49 | 0.71 | 0.75 |
| DPA | 131.1 | 118.0 | 156.0 | 167.4 | 161.4 | 140.5 | 138.6 | 153.4 | 11.95 | 0.76 | 0.77 | 0.69 |
| EPA + DHA | 148.1 | 152.5 | 167.0 | 206.9 | 218.3 | 185.6 | 171.2 | 162.0 | 13.53 | 0.61 | 0.88 | 0.72 |
| EPA + DHA + DPA | 279.1 | 270.5 | 323.0 | 374.3 | 379.8 | 326.1 | 309.9 | 315.5 | 24.89 | 0.70 | 0.97 | 0.74 |
| Item 1 | Treatment | Breed | SEM | p Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 2.5C | 5C | 2.5F | 5F | M × M | C × M | W × C | T | B | T × B | ||
| Total lipid (g fat/100 g wet tissue) | 3.0 | 3.1 | 2.8 | 3.0 | 2.9 | 3.0 | 3.0 | 3.0 | 0.05 | 0.53 | 0.65 | 0.42 |
| Percentage (g/100 g total FA)2 | ||||||||||||
| 14:0 | 0.4 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.02 | 0.44 | 0.77 | 0.65 |
| 15:0 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.01 | 0.68 | 0.58 | 0.68 |
| 16:0 | 17.0 | 17.2 | 16.5 | 16.4 | 16.0 | 16.6 | 17.0 | 16.3 | 0.19 | 0.53 | 0.47 | 0.94 |
| 17:0 | 1.2 a | 1.1 b | 1.1 b | 1.0 b,c | 0.9 c | 1.1 a | 1.1a | 1.0b | 0.02 | 0.00 | 0.02 | 0.07 |
| 18:2n-6 | 12.7 | 12.6 | 12.5 | 12.7 | 12.6 | 12.6 | 12.4 | 12.8 | 0.17 | 0.93 | 0.84 | 0.90 |
| 18:3n-3 | 1.1 | 1.1 | 1.2 | 1.2 | 1.5 | 1.2 | 1.3 | 1.1 | 0.06 | 0.22 | 0.49 | 0.11 |
| 18:1n-9 | 11.9 | 12.7 | 11.7 | 11.3 | 11.6 | 12.1 | 12.4 | 11.1 | 0.25 | 0.52 | 0.12 | 0.51 |
| 18:0 | 17.4 | 17.5 | 17.9 | 17.5 | 17.8 | 17.7 | 17.0 | 18.1 | 0.18 | 0.89 | 0.16 | 0.96 |
| 20:4n-6 | 13.8 | 13.7 | 13.2 | 12.7 | 12.1 | 12.5 b | 13.1 a,b | 14.2 a | 0.29 | 0.27 | 0.04 | 0.69 |
| 20:5n-3 (EPA) | 1.9 b | 2.0 b | 2.3 a,b | 2.5 a | 2.8 a | 2.3 | 2.0 | 2.3 | 0.09 | 0.01 | 0.08 | 0.80 |
| 20:3n-6 | 0.9 | 0.8 | 0.8 | 0.9 | 0.8 | 0.8 | 0.8 | 0.9 | 0.03 | 0.86 | 0.11 | 0.72 |
| 20:4n-3 | 0.6 | 0.7 | 0.7 | 0.7 | 0.7 | 0.7 | 0.6 | 0.6 | 0.02 | 0.63 | 0.17 | 0.71 |
| 20:2n-6 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.4 | 0.01 | 0.91 | 0.65 | 0.27 |
| 20:0 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.01 | 0.30 | 0.29 | 0.32 |
| 22:5n-6 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.01 | 0.65 | 0.79 | 0.85 |
| 22:6n-3 (DHA) | 2.3 | 2.3 | 2.4 | 2.4 | 2.4 | 2.5 a | 2.4 a | 2.1 b | 0.07 | 0.88 | 0.03 | 0.11 |
| 22:4n-6 | 0.4 a | 0.4 a | 0.3 a,b | 0.3 a,b | 0.2 b | 0.3 | 0.4 | 0.4 | 0.02 | 0.04 | 0.14 | 0.88 |
| 22:5n-3 (DPA) | 2.7 b | 2.8 a,b | 3.1 a,b | 3.2 a | 3.3 a | 2.8 b | 2.9 b | 3.2 a | 0.07 | 0.04 | 0.05 | 0.29 |
| 22:0 | 1.2 | 1.2 | 1.1 | 1.1 | 1.1 | 1.1 | 1.1 | 1.2 | 0.04 | 0.85 | 0.33 | 0.93 |
| 23:0 | 0.4 | 0.4 | 0.4 | 0.3 | 0.3 | 0.4 | 0.3 | 0.3 | 0.01 | 0.48 | 0.09 | 0.30 |
| 24:0 | 1.4 | 1.3 | 1.3 | 1.2 | 1.2 | 1.3 | 1.2 | 1.4 | 0.05 | 0.77 | 0.22 | 0.99 |
| ∑SFA | 40.5 | 40.5 | 40.0 | 39.4 | 39.6 | 40.4 | 39.7 | 40.3 | 0.26 | 0.70 | 0.64 | 0.94 |
| ∑MUFA | 21.1 | 21.4 | 20.5 | 21.7 | 22.0 | 21.5 | 22.6 | 19.7 | 0.43 | 0.88 | 0.06 | 0.97 |
| ∑PUFA | 38.4 | 38.1 | 39.5 | 38.9 | 38.4 | 38.1 | 37.7 | 40.0 | 0.40 | 0.87 | 0.10 | 0.79 |
| PUFA/SFA | 0.9 | 0.9 | 1.0 | 1.0 | 1.0 | 0.9 | 0.9 | 1.0 | 0.01 | 0.93 | 0.45 | 0.82 |
| ∑n-3 PUFA | 8.7 b | 8.8 b | 9.8 a,b | 10.0 a | 10.3 a | 9.5 | 8.8 | 9.7 | 0.19 | 0.01 | 0.06 | 0.70 |
| ∑n-6 PUFA | 28.2 | 27.5 | 28.0 | 27.1 | 26.1 | 26.7 | 27.1 | 28.7 | 0.38 | 0.49 | 0.22 | 0.86 |
| n-6/n-3 | 3.3 a | 3.2 a,b | 2.9 a,b,c | 2.7 b,c | 2.5 c | 2.8 | 3.1 | 3.0 | 0.08 | 0.02 | 0.39 | 0.92 |
| Content (mg/100 g wet tissue) | ||||||||||||
| 18:3n-3 | 18.6 | 19.7 | 19.9 | 23.6 | 24.6 | 22.0 | 22.5 | 18.6 | 1.72 | 0.78 | 0.63 | 0.18 |
| 20:4n-6 | 243.6 | 245.0 | 223.2 | 204.3 | 202.7 | 234.5 | 207.0 | 239.6 | 9.35 | 0.46 | 0.66 | 0.48 |
| EPA | 33.7 | 37.6 | 38.1 | 41.7 | 45.5 | 43.0 | 32.3 | 39.9 | 2.00 | 0.39 | 0.17 | 0.86 |
| 22:5n-6 | 2.2 a | 2.2 a | 1.3 b | 1.5 a,b | 1.2 b | 1.8 | 1.6 | 1.9 | 0.13 | 0.02 | 0.82 | 0.67 |
| DHA | 39.2 | 39.2 | 39.3 | 40.5 | 42.1 | 46.5 a | 33.0 b | 40.9 a,b | 1.87 | 0.98 | 0.05 | 0.28 |
| DPA | 50.3 | 49.7 | 50.6 | 51.1 | 51.3 | 52.7 | 45.5 | 53.5 | 1.93 | 0.92 | 0.44 | 0.35 |
| EPA + DHA | 72.9 | 76.8 | 77.4 | 82.2 | 87.6 | 89.5 a | 65.3 b | 80.8 a,b | 3.45 | 0.87 | 0.05 | 0.70 |
| EPA + DHA + DPA | 123.2 | 126.5 | 128.0 | 133.3 | 138.9 | 142.2 | 110.8 | 134.3 | 5.14 | 0.96 | 0.16 | 0.56 |
| Item 1 | Treatment | Breed | SEM | Normal Range | p Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 2.5C | 5C | 2.5F | 5F | M × M | C × M | W × C | T | B | T × B | |||
| Cholesterol | 1.3 | 1.5 | 1.5 | 1.4 | 1.2 | 1.3 | 1.4 | 1.4 | 0.04 | 1.1–1.5 | 0.13 | 0.62 | 0.92 |
| Urea | 7.8 | 7.7 | 7.5 | 7.6 | 7.5 | 7.8 | 7.4 | 7.3 | 0.16 | 2.8−7.2 | 0.27 | 0.44 | 0.93 |
| Calcium | 2.6 | 2.6 | 2.7 | 2.6 | 2.7 | 2.7 | 2.7 | 2.6 | 0.02 | 2.4−3.2 | 0.26 | 0.15 | 0.61 |
| Magnesium | 1.1 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.1 | 0.02 | 0.8−1.2 | 0.83 | 0.64 | 0.62 |
| BHB | 0.3 | 0.3 | 0.4 | 0.3 | 0.4 | 0.3 | 0.4 | 0.3 | 0.02 | 0.0−0.8 | 0.66 | 0.85 | 0.95 |
| Glucose | 4.2 | 4.5 | 4.3 | 4.1 | 4.2 | 4.0 b | 4.3 a,b | 4.5 a | 0.07 | 2.8−4.5 | 0.48 | 0.03 | 0.04 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, D.V.; Le, V.H.; Nguyen, Q.V.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Omega–3 Long-Chain Fatty Acids in the Heart, Kidney, Liver and Plasma Metabolite Profiles of Australian Prime Lambs Supplemented with Pelleted Canola and Flaxseed Oils. Nutrients 2017, 9, 893. https://doi.org/10.3390/nu9080893
Nguyen DV, Le VH, Nguyen QV, Malau-Aduli BS, Nichols PD, Malau-Aduli AEO. Omega–3 Long-Chain Fatty Acids in the Heart, Kidney, Liver and Plasma Metabolite Profiles of Australian Prime Lambs Supplemented with Pelleted Canola and Flaxseed Oils. Nutrients. 2017; 9(8):893. https://doi.org/10.3390/nu9080893
Chicago/Turabian StyleNguyen, Don V., Van H. Le, Quang V. Nguyen, Bunmi S. Malau-Aduli, Peter D. Nichols, and Aduli E. O. Malau-Aduli. 2017. "Omega–3 Long-Chain Fatty Acids in the Heart, Kidney, Liver and Plasma Metabolite Profiles of Australian Prime Lambs Supplemented with Pelleted Canola and Flaxseed Oils" Nutrients 9, no. 8: 893. https://doi.org/10.3390/nu9080893
APA StyleNguyen, D. V., Le, V. H., Nguyen, Q. V., Malau-Aduli, B. S., Nichols, P. D., & Malau-Aduli, A. E. O. (2017). Omega–3 Long-Chain Fatty Acids in the Heart, Kidney, Liver and Plasma Metabolite Profiles of Australian Prime Lambs Supplemented with Pelleted Canola and Flaxseed Oils. Nutrients, 9(8), 893. https://doi.org/10.3390/nu9080893