Effects of Six-Week Ginkgo biloba Supplementation on Aerobic Performance, Blood Pro/Antioxidant Balance, and Serum Brain-Derived Neurotrophic Factor in Physically Active Men



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design
2.3. Biochemical Analyses
2.4. Statistical Analysis
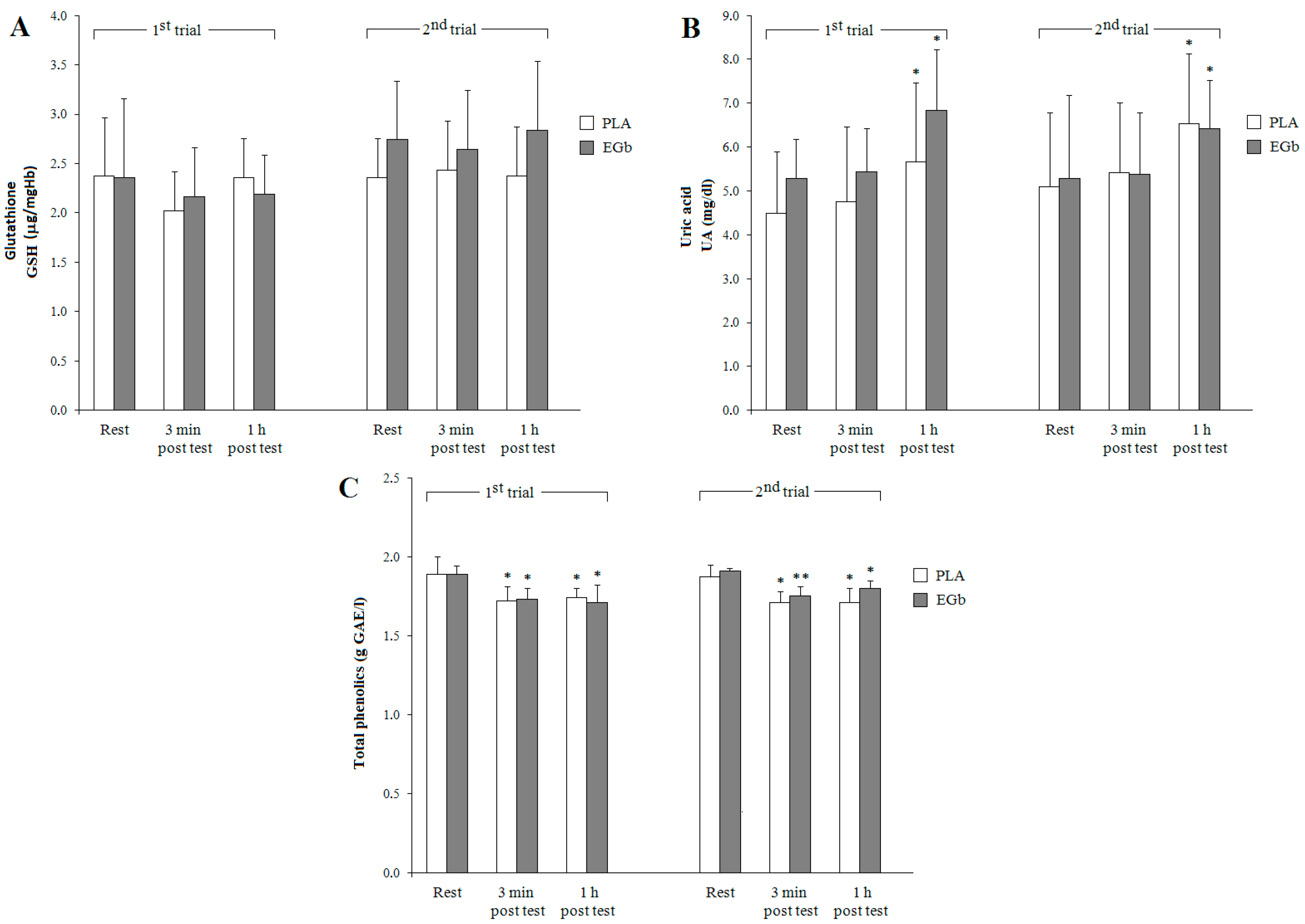
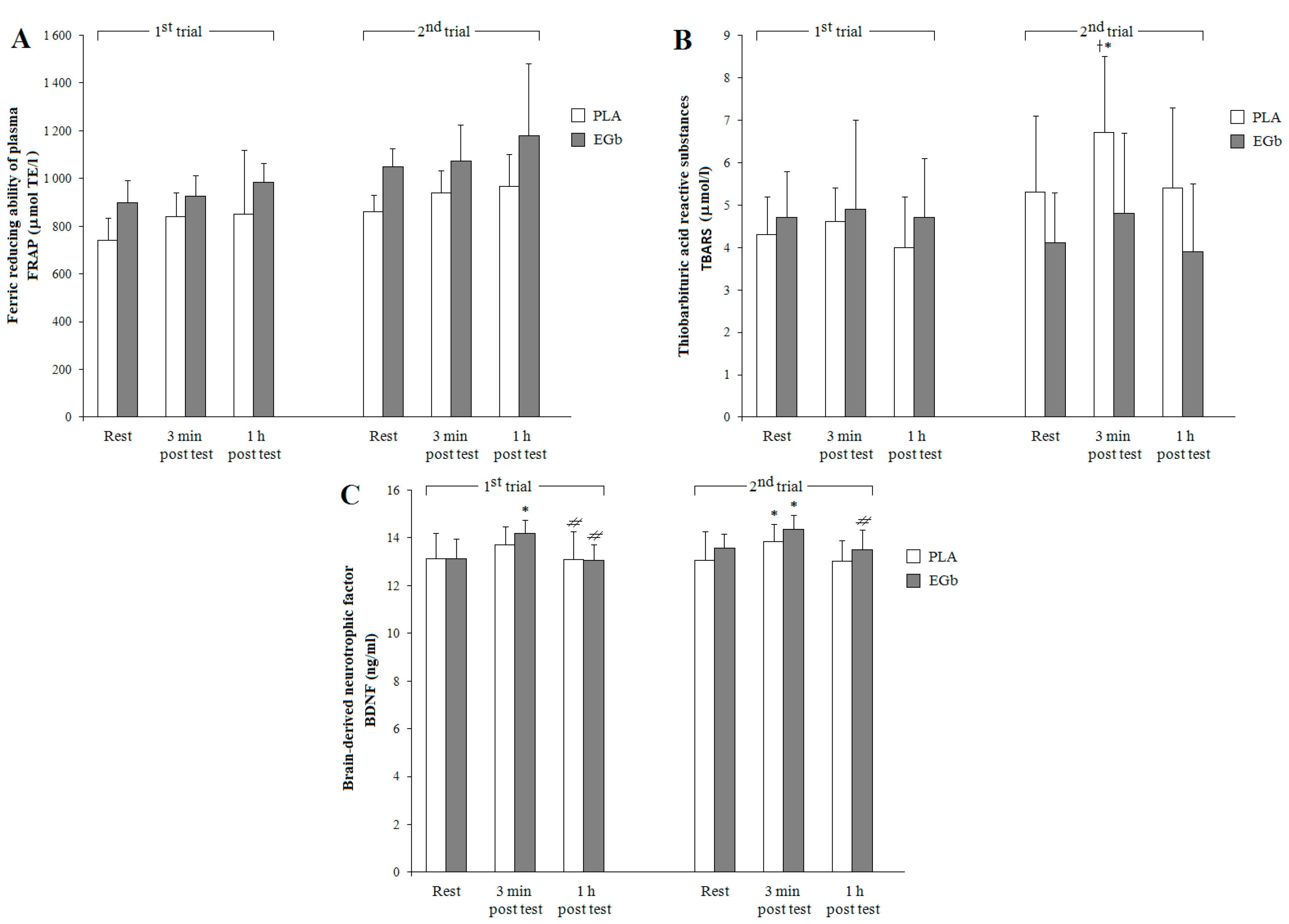
3. Results
3.1. Subjects
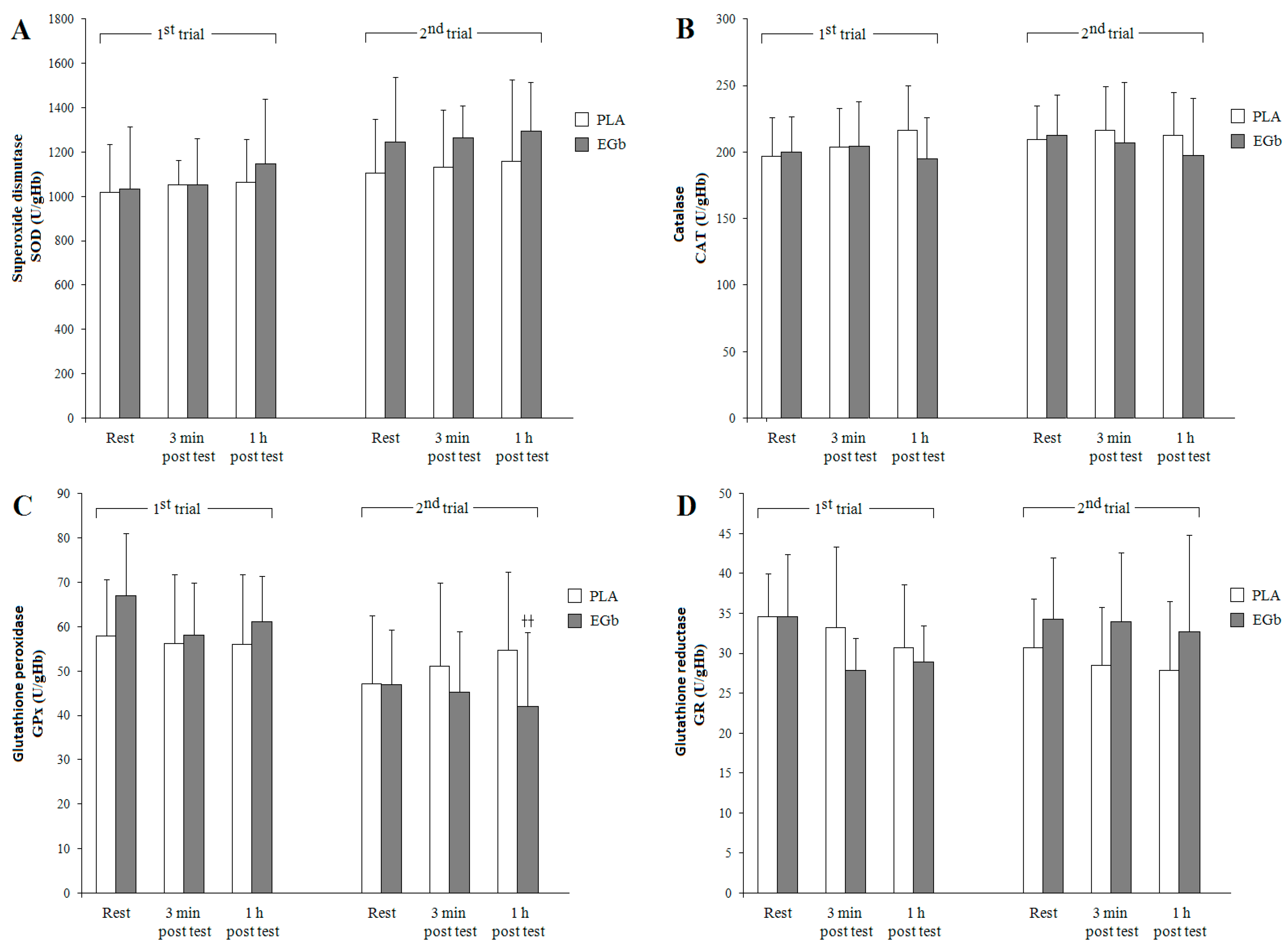
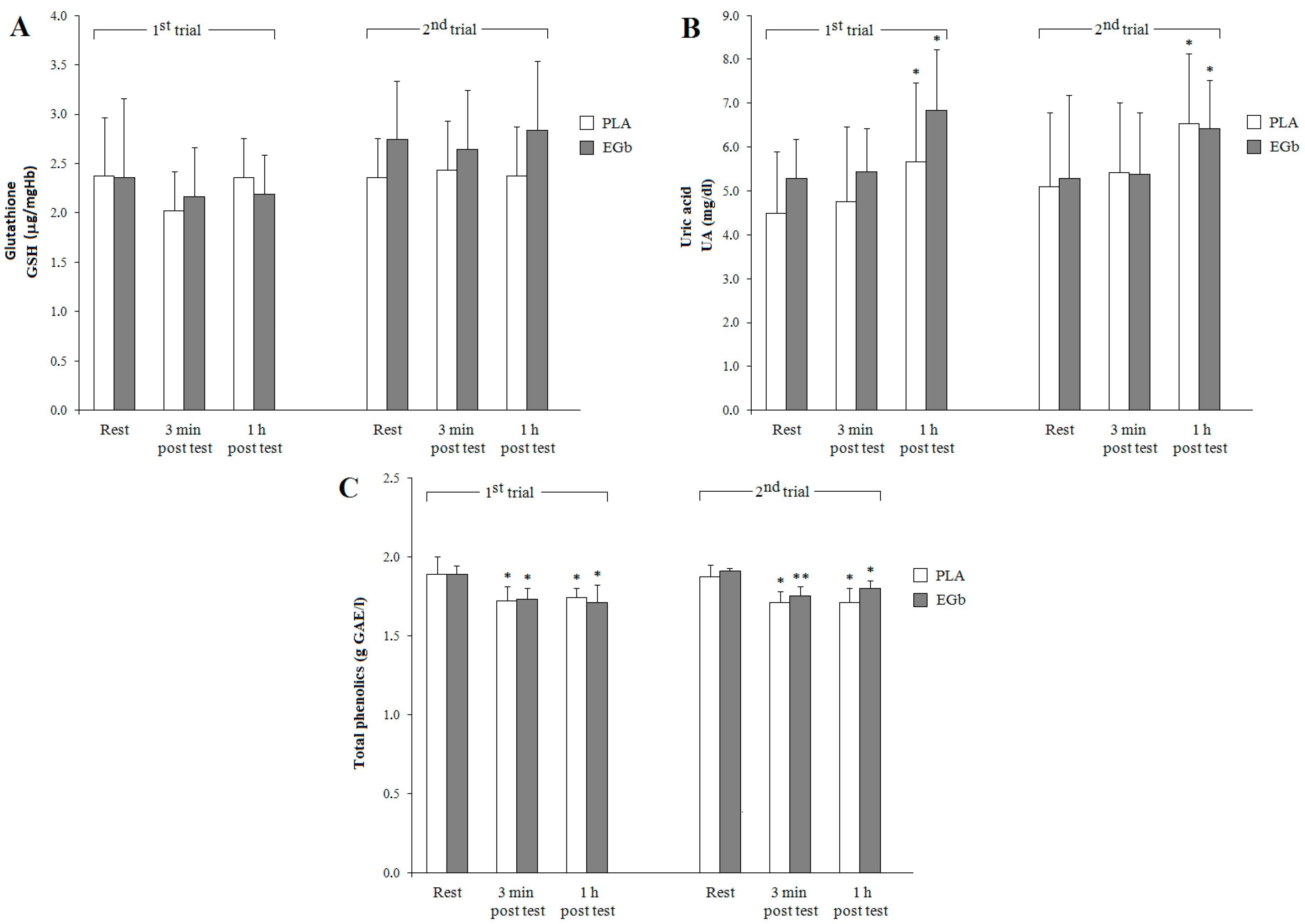
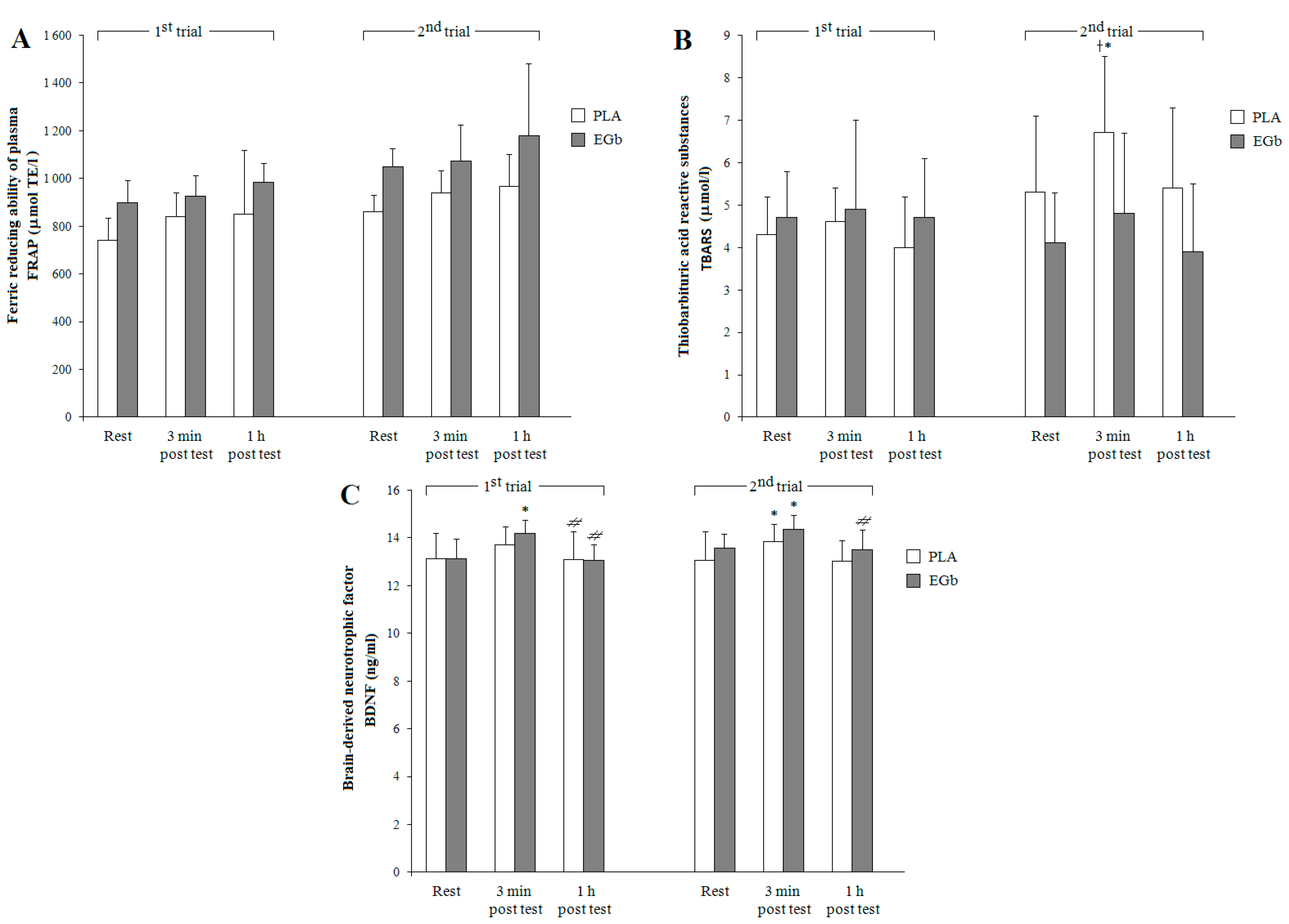
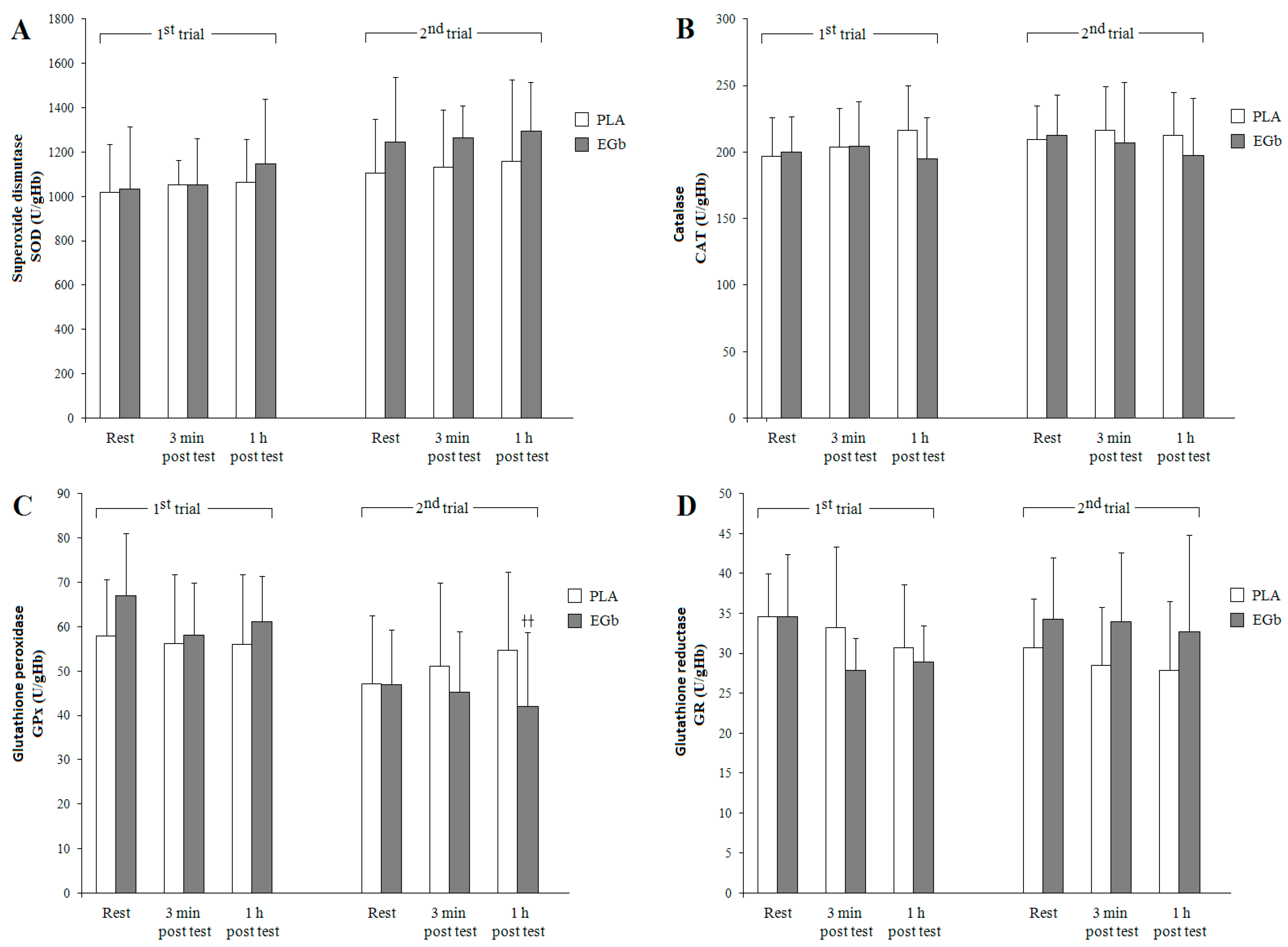
3.2. Antioxidant Capacities
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and human health: Prevention of disease and mechanisms of action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef] [PubMed]
- Willcox, J.K.; Ash, S.L.; Catignani, G.L. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr. 2004, 44, 275–295. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E., Jr.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 526, 673–751. [Google Scholar] [PubMed]
- Croft, K.D. Dietary polyphenols: Antioxidants or not? Arch. Biochem. Biophys. 2016, 595, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Droy-Lefaix, M.T. Effect of the antioxidant action of Ginkgo biloba extract (EGb 761) on aging and oxidative stress. Age 1997, 29, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Holst, B. Dietary reference intake (DRI) value for dietary polyphenols: Are we heading in the right direction? Br. J. Nutr. 2008, 99 (Suppl. 3), S55–S58. [Google Scholar] [CrossRef] [PubMed]
- Ude, C.; Schubert-Zsilavecz, M.; Wurglics, M. Ginkgo biloba extracts: A review of the pharmacokinetics of the active ingredients. Clin. Pharmacokinet. 2013, 52, 727–749. [Google Scholar] [CrossRef] [PubMed]
- Mullaicharam, A.R. A review on evidence based practice of Ginkgo biloba in brain health. Int. J. Pharmaceut. Chem. Anal. 2013, 1, 24–30. [Google Scholar]
- Kuo, Y.C.; Lin, J.C.; Bernard, J.R.; Liao, Y.H. Green tea extract supplementation dose not hamper endurance-training adaptation but improves antioxidant capacity in sedentary men. Appl. Physiol. Nutr. Metab. 2015, 40, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Spencer, J.P.E.; Rice-Evans, C. Flavonoids: Antioxidants or signalin g molecules? Free Radic. Biol. Med. 2004, 36, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Manach, C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr. 2005, 81, 243S–255S. [Google Scholar] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [PubMed]
- Bell, L.; Lamport, J.; Butler, L.T.; Williams, C.M. A review of the cognitive effects observed in humans following acute supplementation with flavonoids, and their associated mechanisms of action. Nutrients 2015, 7, 10290–10306. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [PubMed]
- DeFeudis, F.V. Bilobalide and neuroprotection. Pharmacol. Res. 2002, 46, 565–568. [Google Scholar] [CrossRef]
- DeFeudis, F.V.; Drieu, K. Ginkgo biloba extract (EGb 761) and CNS functions: Basic studies and clinical applications. Curr. Drug Targets 2000, 1, 25–58. [Google Scholar] [CrossRef] [PubMed]
- Autry, A.E.; Monteggia, L.M. Brain-derived neurotrophic factor and neuropsychiatric disorders. Pharmacol. Rev. 2012, 64, 238–258. [Google Scholar] [CrossRef] [PubMed]
- Cotman, C.W.; Berchtold, N.C.; Christie, L.A. Exercise builds brain health: Key roles of growth factor cascades and inflammation. Trends Neurosci. 2007, 30, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Beutler, E.; Duron, O.; Kelly, B.M. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Glatzle, G.; Korner, W.F.; Christeller, S.; Wiss, O. Method for the detection of biochemical riboflavin deficiency stimulation of NADPH2-dependent glutathione reductase from human erythrocytes by FAD in vitro. Investigations on the vitamin B2 status-in healthy people and geriatric patients. Int. Z. Vitaminforsch. 1970, 40, 166–183. [Google Scholar] [PubMed]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Davis, J.M.; Carlstedt, C.J.; Chen, S.; Carmichael, M.D.; Murphy, E.A. The dietary flavonoid quercetin increases VO2max and endurance capacity. Int. J. Sport Nutr. Exerc. Metab. 2010, 20, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.C.; Lonac, M.C.; Johnson, T.K.; Schweder, M.M.; Bell, C. Epigallocatechin-3-gallate increases maximal oxygen uptake in adult humans. Med. Sci. Sports Exerc. 2010, 42, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Williams, M. Dietary supplements and sports performance: Herbals. J. Int. Soc. Sports Nutr. 2006, 3, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Peters, H.; Kieser, M.; Hölscher, U. Demonstration of the efficacy of ginkgo biloba special extract EGb 761 on intermittent claudication—A placebo-controlled, double-blind multicenter trial. Vasa 1998, 27, 106–110. [Google Scholar] [PubMed]
- Nicolaï, S.P.A.; Kruidenier, L.M.; Bendermacher, B.L.W.; Prins, M.H.; Stockmans, R.A.; Broos, P.P.H.L.; Teijink, J.A.W. Ginkgo biloba for intermittent claudication. Cochrane Database Syst. Rev. 2013, 6, CD006888. [Google Scholar] [CrossRef]
- Kressler, J.; Millard-Stafford, M.; Warren, G.L. Quercetin and endurance exercise capacity: A systemic review and meta-analysis. Med. Sci. Sports Exerc. 2011, 43, 2396–2404. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, D.M.; Lacerte, G.; Goulet, E.D.B. Effects of quercetin supplementation on endurance performance and maximal oxygen consumption: A meta-analysis. Int. J. Sport. Nutr. Exerc. Metab. 2013, 23, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Mahady, G.B. Ginkgo biloba: A review of quality, safety, and efficacy. Nutr. Clin. Care 2001, 4, 140–147. [Google Scholar] [CrossRef]
- Coimbra, S.; Castro, E.; Rocha-Pereira, P.; Rebelo, I.; Rocha, S.; Santos-Silva, A. The effect of green tea in oxidative stress. Clin. Nutr. 2006, 25, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Abreu, I.A.; Cabelli, D.E. Superoxide dismutases—A review of the metal-associated mechanistic variations. Biochim. Biophys. Acta 2010, 1804, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.R. The glutathione peroxidases. Cell Mol. Life Sci. 2000, 57, 1825–1835. [Google Scholar] [CrossRef] [PubMed]
- Moskaug, J.Ø.; Carlsen, H.; Myhrstad, M.C.W.; Blomhoff, R. Polyphenols and glutathione synthesis regulation. Am. J. Clin. Nutr. 2005, 81, 277S–283S. [Google Scholar] [PubMed]
- Hellsten, Y.; Richter, E.A.; Kiens, B.; Bangsbo, J. AMP deamination and purine exchange in human skeletal muscle during and after intense exercise. J. Physiol. 1999, 520, 909–920. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, E.P.; Burini, R.C. High plasma uric acid concentration: Causes and consequences. Diabetol. Metab. Syndr. 2012, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Dudzińska, W.; Lubkowska, A.; Dolegowska, B.; Suska, M.; Janiak, M. Uridine—An indicator of post-exercise uric acid concentration and blood pressure. Physiol. Res. 2015, 64, 467–477. [Google Scholar] [PubMed]
- Hellsten-Westing, Y.; Ekblom, B.; Sjödin, B. The metabolic relation between hypoxanthine and uric acid in man following maximal short-distance running. Acta Physiol. Scand. 1989, 137, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Witkowska, A.M.; Zujko, M.E.; Waśkiewicz, A.; Terlikowska, K.M.; Piotrowski, W. Comparison of various databases for estimation of dietary polyphenol intake in the population of Polish adults. Nutrients 2015, 7, 9299–9308. [Google Scholar] [CrossRef] [PubMed]
- Hollman, P.C.H.; Cassidy, A.; Compe, B.; Heinonen, M.; Richelle, M.; Richling, E.; Serafini, M.; Scalbert, A.; Sies, H.; Vidry, S. The biological relevance of direct antioxidant effects of polyphenols for cardiovascular health in humans is not established. J. Nutr. 2011, 141, 989S–1009S. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Davies, K.J.A.; Ursini, F. How do nutritional antioxidants really work: Nucleophilic tone and para-hormesis versus free radical scavenging in vivo. Free Radic. Biol. Med. 2014, 66, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Jówko, E.; Długołęcka, B.; Makaruk, B.; Cieśliński, I. The effect of green tea extract supplementation on exercise-induced oxidative stress parameters in male sprinters. Eur. J. Nutr. 2015, 54, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Bartosz, G. Total antioxidant capacity. Adv. Clin. Chem. 2003, 37, 219–292. [Google Scholar] [PubMed]
- Kamińska, J.; Podgórski, T.; Pawlak, M. Variability of selected hematological and biochemical markers in marathon runners. Trends Sport Sci. 2015, 3, 125–132. [Google Scholar]
- Rassmussen, P.; Brassard, P.; Adser, H.; Pedersen, M.V.; Leick, L.; Hart, E.; Secher, N.H.; Pedersen, B.K.; Pilegaard, H. Evidence for a release of brain-derived neurotrophic factor from the brain during exercise. Exp. Physiol. 2009, 94, 1062–1069. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Voss, M.V.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; Kim, J.S.; Heo, S.; Alves, H.; White, S.M.; et al. Exercise training increases size of hippocampus and improves memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3017–3022. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Larsen, K.T.; Ried-Larsen, M.; Møller, N.C.; Andersen, L.B. The effects of physical activity and exercise on brain-derived neurotrophic factor in healthy humans: A review. Scand. J. Med. Sci. Sports 2014, 24, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I. The impact of exercise training on basal BDNF in athletic adolescents. J. Phys. Ther. Sci. 2016, 28, 3056–3069. [Google Scholar] [CrossRef] [PubMed]
- Szuhany, K.L.; Bugatti, M.; Otto, M.W. A meta-analytic review of the effects of exercise on brain-derived neurotrophic factor. J. Psychiatr. Res. 2015, 60, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Gomes da Silva, S.; Arida, R.M. Physical activity and brain development. Expert Rev. Neurother. 2015, 15, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.P.; Abd El Mohsena, M.M.; Minihanea, A.M.; Mathers, J.C. Biomarkers of the intake of dietary polyphenols: Strengths, limitations and application in nutrition research. Br. J. Nutr. 2008, 99, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Vega, S.R.; Strüder, H.K.; Wahrmann, B.V.; Schmidt, A.; Bloch, W.; Hollmann, W. Acute BDNF and cortisol response to low intensity exercise and following ramp incremental exercise to exhaustion in humans. Brain Res. 2006, 1121, 59–65. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Group | Age (Years) | Height (cm) | Body Weight (kg) | VO2max (mL/min/kg) | |
|---|---|---|---|---|---|
| First Trial | Second Trial | ||||
| PLA | 22.3 ± 1.1 | 179.7 ± 7.8 | 79.7 ± 10.8 | 45.7 ± 5.1 | 46.1 ± 4.1 |
| EGb | 22.5 ± 0.9 | 182.1 ± 5.9 | 80.1 ± 6.8 | 46.2 ± 4.8 | 48.9 ± 5.8 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadowska-Krępa, E.; Kłapcińska, B.; Pokora, I.; Domaszewski, P.; Kempa, K.; Podgórski, T. Effects of Six-Week Ginkgo biloba Supplementation on Aerobic Performance, Blood Pro/Antioxidant Balance, and Serum Brain-Derived Neurotrophic Factor in Physically Active Men. Nutrients 2017, 9, 803. https://doi.org/10.3390/nu9080803
Sadowska-Krępa E, Kłapcińska B, Pokora I, Domaszewski P, Kempa K, Podgórski T. Effects of Six-Week Ginkgo biloba Supplementation on Aerobic Performance, Blood Pro/Antioxidant Balance, and Serum Brain-Derived Neurotrophic Factor in Physically Active Men. Nutrients. 2017; 9(8):803. https://doi.org/10.3390/nu9080803
Chicago/Turabian StyleSadowska-Krępa, Ewa, Barbara Kłapcińska, Ilona Pokora, Przemysław Domaszewski, Katarzyna Kempa, and Tomasz Podgórski. 2017. "Effects of Six-Week Ginkgo biloba Supplementation on Aerobic Performance, Blood Pro/Antioxidant Balance, and Serum Brain-Derived Neurotrophic Factor in Physically Active Men" Nutrients 9, no. 8: 803. https://doi.org/10.3390/nu9080803
APA StyleSadowska-Krępa, E., Kłapcińska, B., Pokora, I., Domaszewski, P., Kempa, K., & Podgórski, T. (2017). Effects of Six-Week Ginkgo biloba Supplementation on Aerobic Performance, Blood Pro/Antioxidant Balance, and Serum Brain-Derived Neurotrophic Factor in Physically Active Men. Nutrients, 9(8), 803. https://doi.org/10.3390/nu9080803