Modulatory Effects of Guarana (Paullinia cupana) on Adipogenesis

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. 3T3L1 Cell Culture
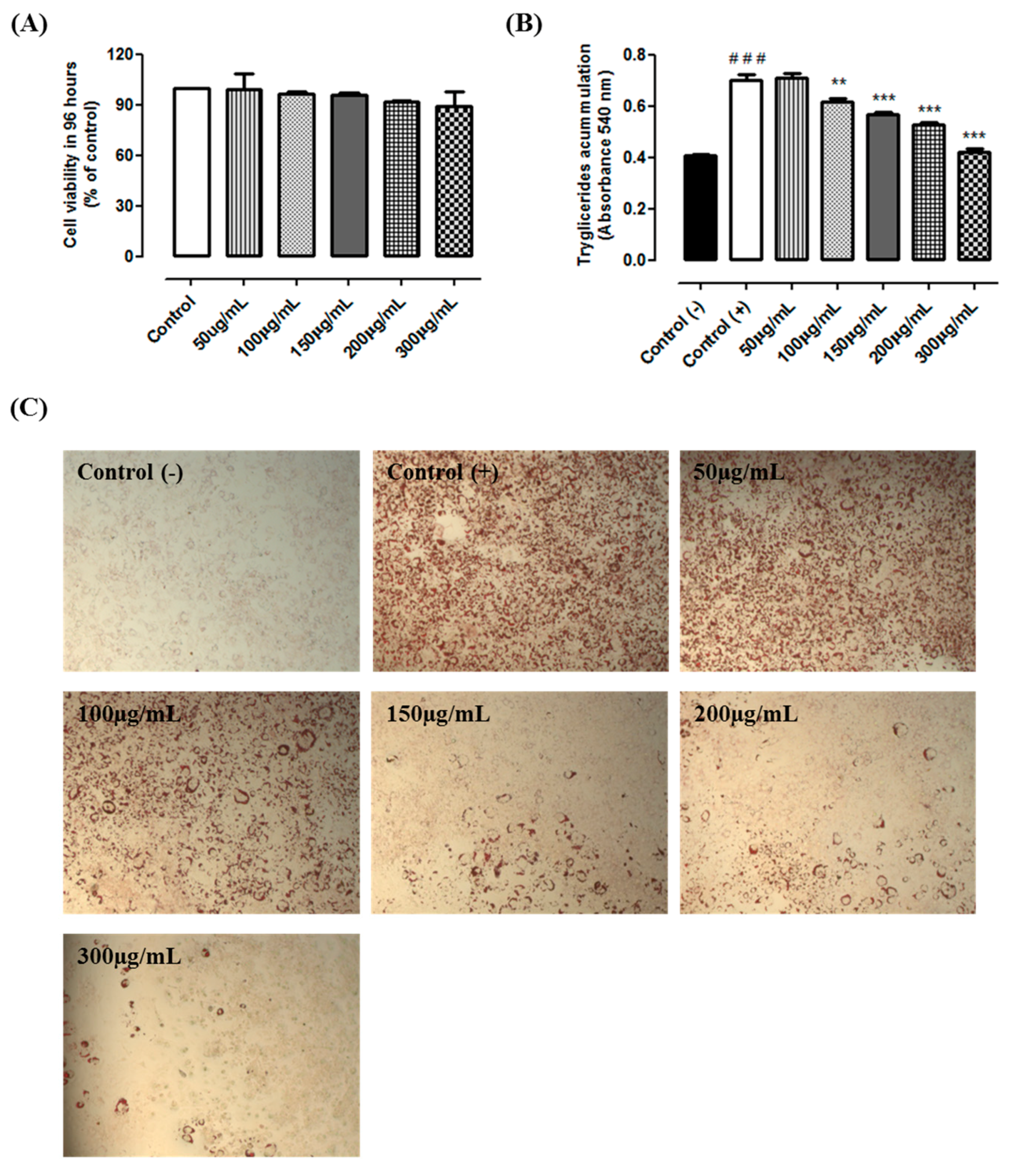
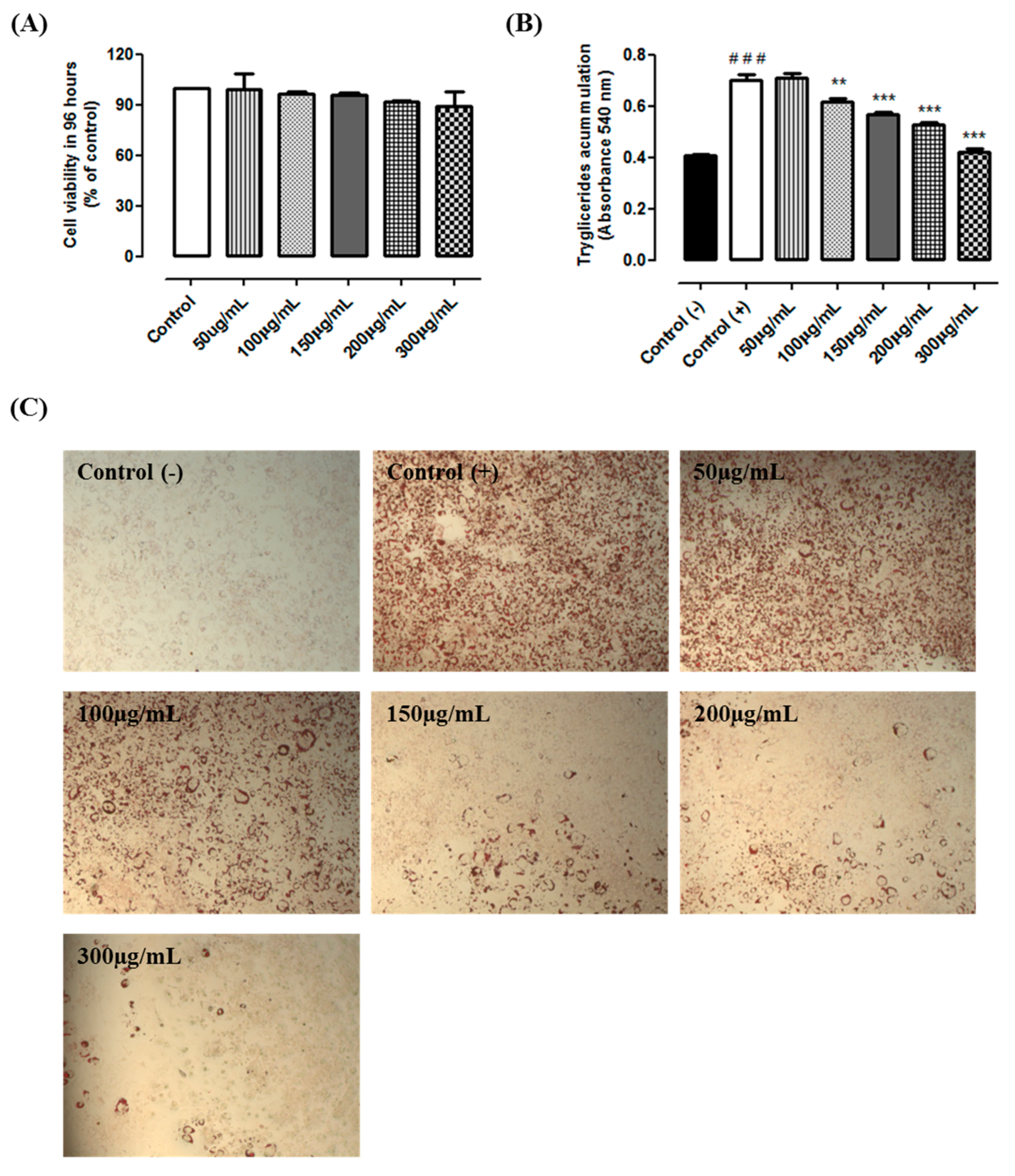
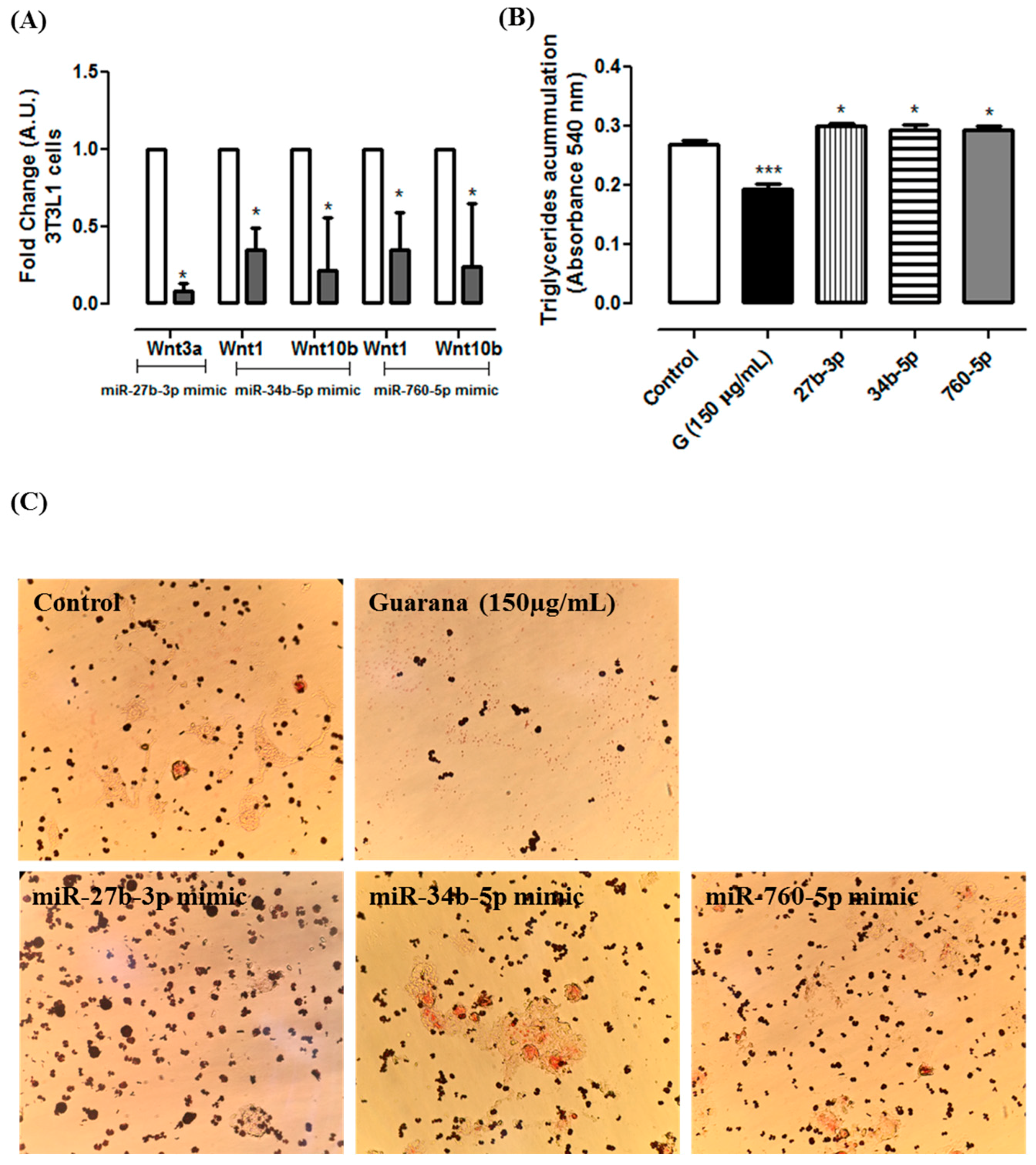
2.2. Cytotoxicity and Triacylglycerol Accumulation
2.3. Experimental Design
2.4. Total Phenolic Content
2.5. Total Flavonoid Content
2.6. Caffeine Quantification by Mass Spectrometry
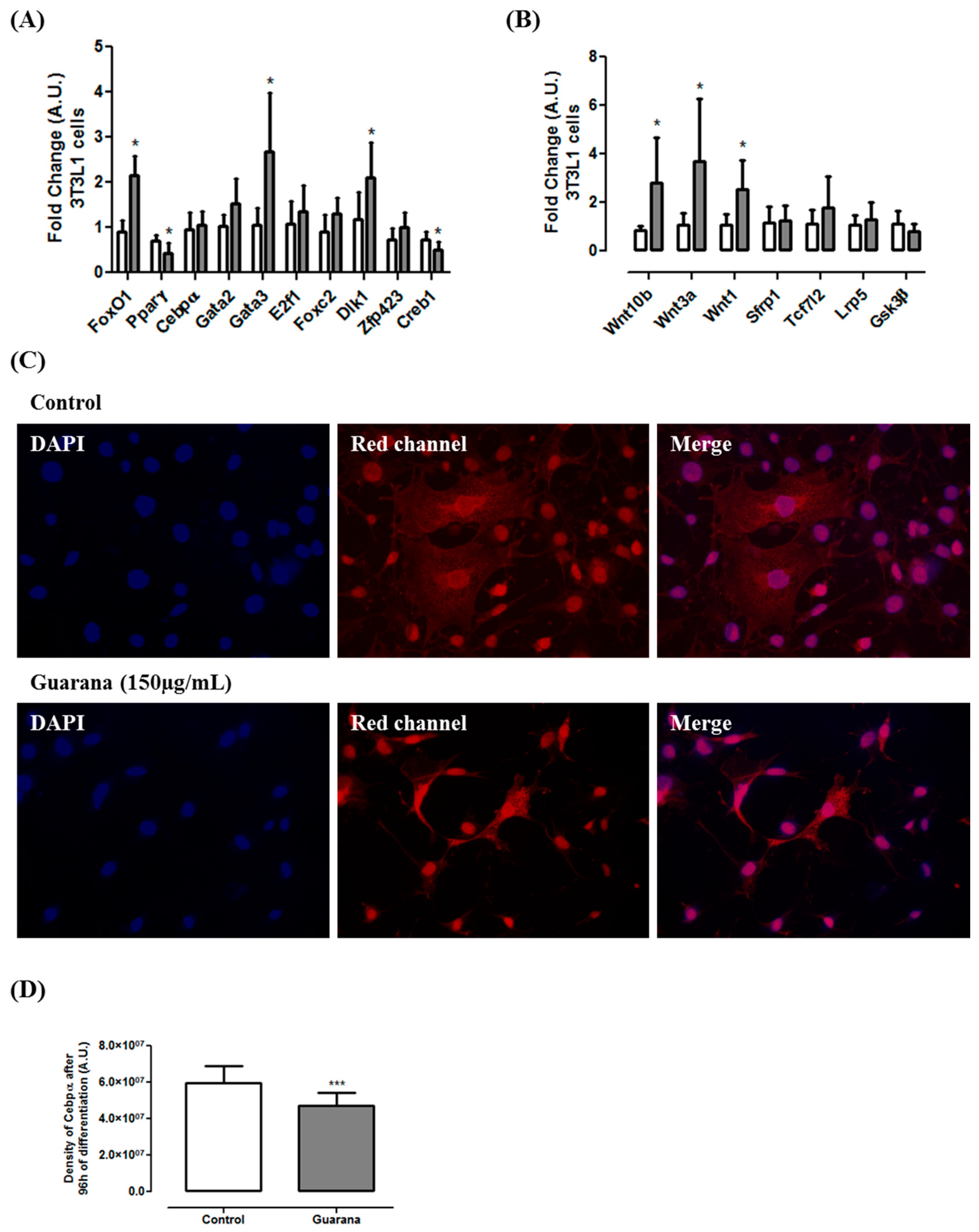
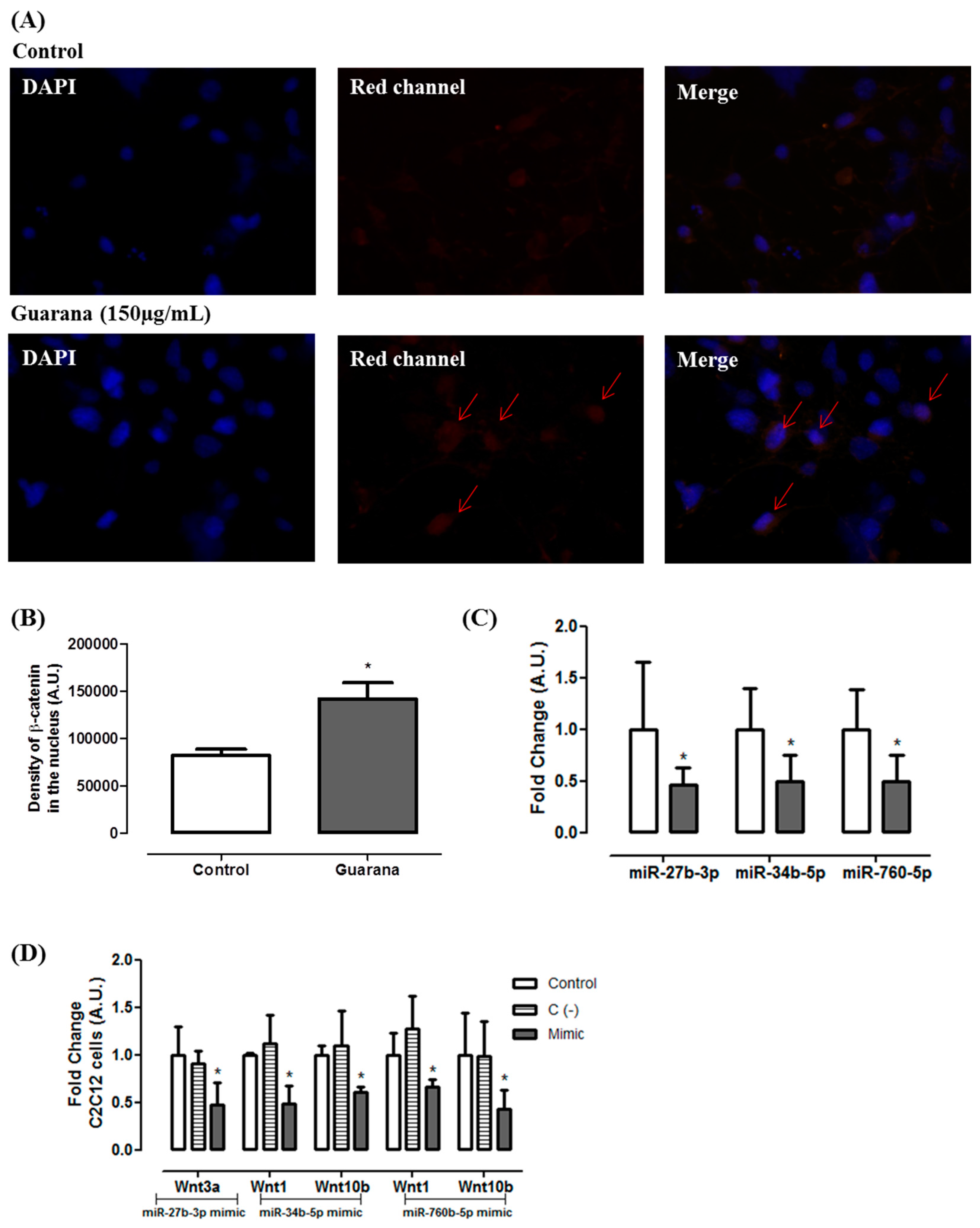
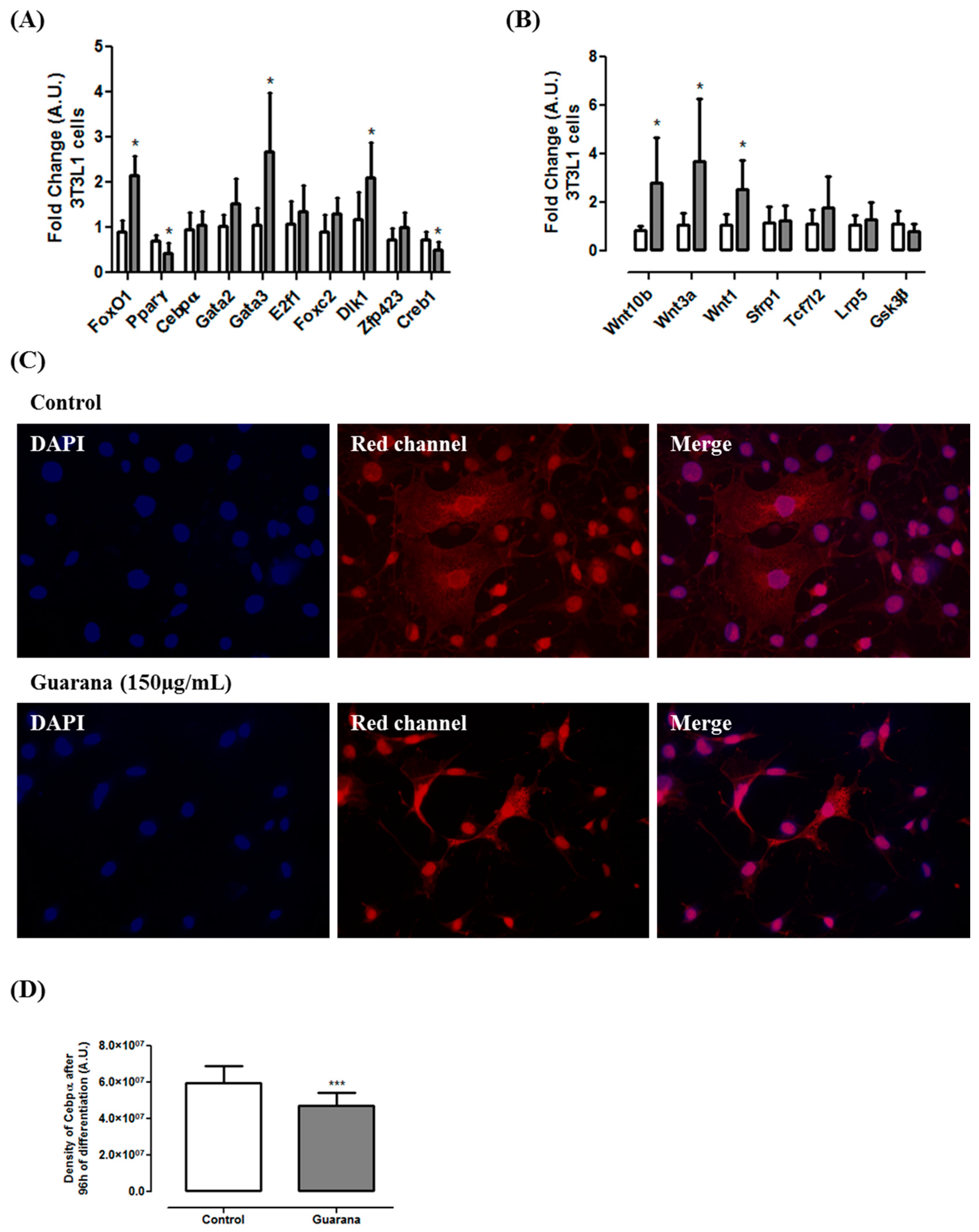
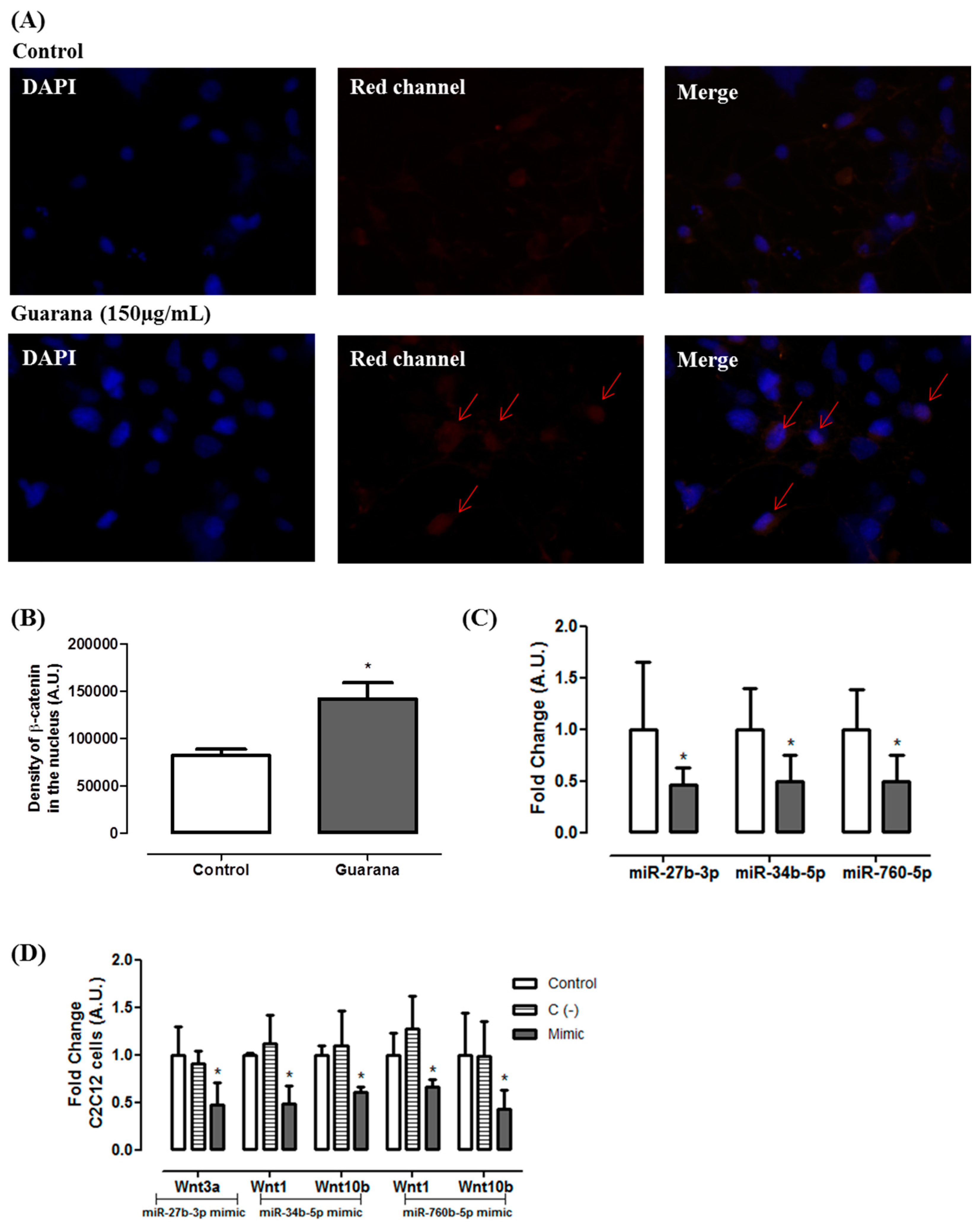
2.7. mRNA and miRNA Expression
2.8. miR Mimics Transfections
2.9. Immunocytochemistry
2.10. Statistical Analysis
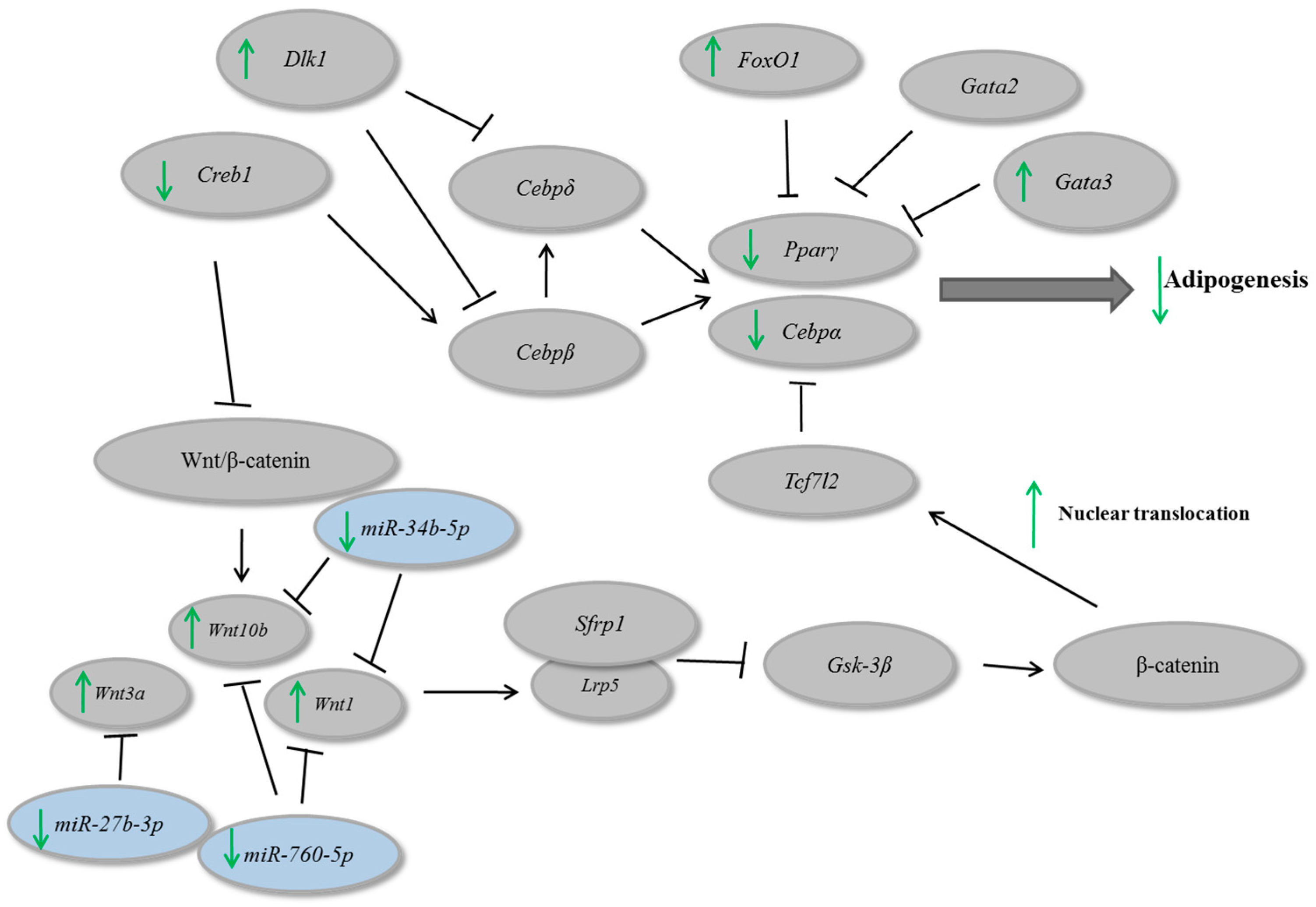
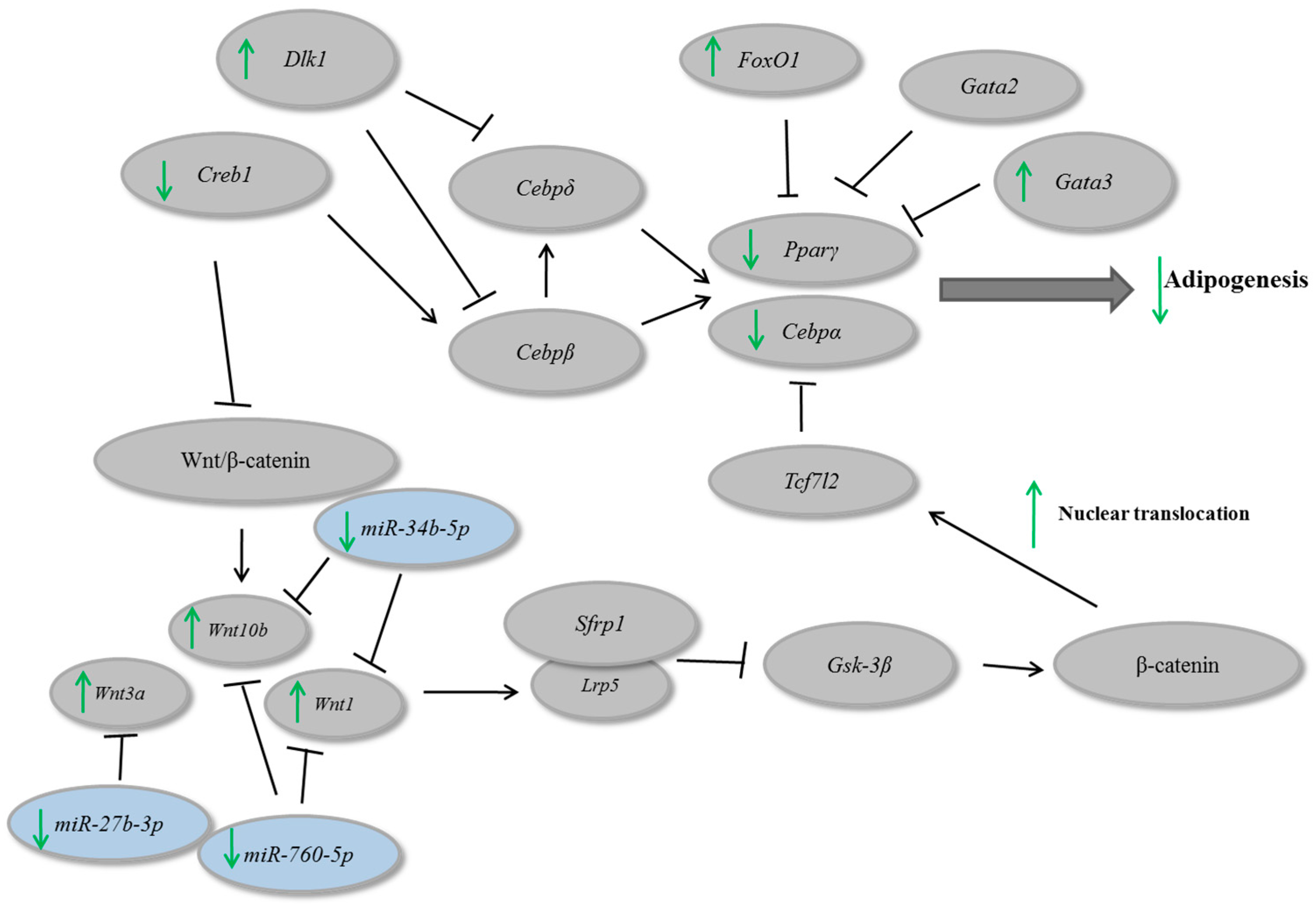
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization. Overweight and Obesity. 2014. Available online: http://www.who.int/gho/ncd/risk_factors/overweight/en/ (accessed on 21 March 2017).
- Phillips, C.M. Metabolically healthy obesity across the life course: Epidemiology, determinants, and implications. Ann. N. Y. Acad. Sci. 2017, 1391, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.; Netterstrøm, M.K. Metabolically healthy obesity and ischemic heart disease: A 10-year follow-up of the Inter99 study. J. Clin. Endocrinol. Metab. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell. Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Kloosterman, W.P.; Plasterk, R.H. The diverse functions of microRNAs in animal development and disease. Dev. Cell 2006, 11, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Xu, Y. miR-27a attenuates adipogenesis and promotes osteogenesis in steroid-induced rat BMSCs by targeting PPARγ and GREM1. Sci. Rep. 2016, 2, 38491. [Google Scholar] [CrossRef] [PubMed]
- Angelo, P.C.; Nunes-Silva, C.G.; Nunes-Silva, M.M.; Brígido, J.S.N.; Azevedo, E.N.; Assunção, A.R.B.; Sousa, F.J.B.; Patrício, M.M.; Rego, J.C.C.; Peixoto, W.P.; et al. Guarana (Paullinia cupana var. sorbilis), an anciently consumed stimulant from the Amazon rain forest: The seeded-fruit transcriptome. Plant Cell Rep. 2008, 27, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Krewer Cda, C.; Ribeiro, E.E. Habitual intake of guarana and metabolic morbidities: An epidemiological study of an elderly Amazonian population. Phytother. Res. 2011, 25, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Portella Rde, L.; Barcelos, R.P. Guaraná (Paullinia cupana Kunth) effects on LDL oxidation in elderly people: An in vitro and in vivo study. Lipids Health Dis. 2013, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Harrold, J.A.; Hughes, G.M. Acute effects of a herb extract formulation and inulin fibre on appetite, energy intake and food choice. Appetite 2013, 62, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.; Gotardo, E.M. Effects of yerba maté, a plant extract formulation (“YGD”) and resveratrol in 3T3-L1 adipogenesis. Molecules 2014, 19, 16909–16924. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteau Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Da Silva, L.A.L.; Pezzini, B.R. Spectrophotometric determination of the total flavonoid content in Ocimum basilicum L. (Lamiaceae) leaves. Pharmacogn. Mag. 2015, 11, 96–101. [Google Scholar] [PubMed]
- Arçari, D.P.; Santos, J.C. The in vitro and in vivo effects of yerba mate (Ilex paraguariensis) extract on adipogenesis. Food Chem. 2013, 141, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Yoon, B.K. Caffeine inhibits adipogenesis through modulation of mitotic clonal expansion and the AKT/GSK3 pathway in 3T3-L1 adipocytes. BMB Rep. 2016, 49, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, H.; Hashimoto, T. Inhibitory effects of caffeine and its metabolites on intracellular lipid accumulation in murine 3T3-L1 adipocytes. Biofactors 2008, 34, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Su, S.H.; Shyu, H.W. Caffeine inhibits adipogenic differentiation of primary adipose-derived stem cells and bone marrow stromal cells. Toxicol. In Vitro 2013, 27, 1830–1837. [Google Scholar] [CrossRef] [PubMed]
- Finelli, C.; Tarantino, G. Have guidelines addressing physical activity been established in nonalcoholic fatty liver disease? World J. Gastroenterol. 2012, 18, 6790–6800. [Google Scholar] [CrossRef] [PubMed]
- Colquitt, J.L.; Loveman, E. Diet, physical activity, and behavioural interventions for the treatment of overweight or obesity in preschool children up to the age of 6 years. Cochrane Database Syst. Rev. 2016, 10, CD012105. [Google Scholar]
- Williams, C.L.; Hayman, L.L. Cardiovascular health in childhood: A statement for health professionals from the Committee on Atherosclerosis, Hypertension, and Obesity in the Young (AHOY) of the Council on Cardiovascular Disease in the Young. Am. Heart Assoc. Circ. 2002, 106, 143–160. [Google Scholar] [CrossRef]
- Raynor, H.A.; Champagn, C.M. Position of the academy of nutrition and dietetics: Interventions for the treatment of overweight and obesity in adults. J. Acad. Nutr. Diet. 2016, 116, 129–147. [Google Scholar] [CrossRef] [PubMed]
- Babu, P.V.; Liu, D. Green tea catechins and cardiovascular health: An update. Curr. Med. Chem. 2008, 15, 1840–1850. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Ye, X. Green tea polyphenols reduced fat deposits in high fat-fed rats via ERK1/2-PPARγ-adiponectin pathway. PLoS ONE 2013, 8, e53796. [Google Scholar] [CrossRef]
- Cunha, C.A.; Lira, F.S. Green tea extract supplementation induces the lipolytic pathway, attenuates obesity, and reduces low-grade inflammation in mice fed a high-fat diet. Mediators Inflamm. 2013, 2013, 635470. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.E.; Hemati, N. Inhibition of adipogenesis by Wnt signaling. Science 2000, 289, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.H.; Butte, A.J. Prediction of preadipocyte differentiation by gene expression reveals role of insulin receptor substrates and necdin. Nat. Cell Biol. 2005, 7, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Walkey, C.J. Transcriptional regulation of adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence 5′→3′ |
|---|---|
| FoxO1 Fw | ACATTTCGTCCTCGAAGCAG |
| FoxO1 Rv | CAGGTCATCCTGCTCTGTCA |
| Pparγ Fw | GATGGAAGACCACTCGCATT |
| Pparγ Rv | AACCATTGGGTCAGCTCTTG |
| Cepbα Fw | GGGACCATTAGCCTTGTGTG |
| Cebpα Rv | CTCTGGGATGGATCGATTGT |
| Gata2 Fw | TGAAGAAGGAAGGGATCCAG |
| Gata2 Rv | TGGAGAGCTCCTCGAAACAT |
| Gata3 Fw | AAGTGCAAAAAGGTGCATGA |
| Gata3 Rv | CAGGGATGACATGTGTCTGG |
| E2f1 Fw | AGCCTAGGGATTCAGGGTGT |
| E2f1 Rv | TGGATCGTGCTATTCCAATG |
| Foxc2 Fw | AGGAGGCCGAGAAGAAAGTC |
| Foxc2 Rv | GCTCAGCGTCTCCACCTT |
| Dlk1 Fw | GGAAAAATTGGCCCCTTTAG |
| Dlk1 Rv | TGGAGTCTTGGGCTAGGGTA |
| Zfp423 Fw | CTTCGAGTCTCTGGCAGACC |
| Zfp423 Rv | CTTGCTGGAGGGAGATGAAG |
| Creb1 Fw | TTTGTCCTTGCTTTCCGAAT |
| Creb1 Rv | CACTTTGGCTGGACATCTTG |
| Wnt10b Fw | TTCTCTCGGGATTTCTTGGA |
| Wnt10b Rv | CACTTCCGCTTCAGGTTTTC |
| Wnt3a Fw | GCACTAGTGTTGAGGCAATGGTCACCAG |
| Wnt3a Rv | ATACGCGTGAACGCAAAGTTCCAGGCAG |
| Wnt1 Fw | GCACTAGTAGGGTTCATAGCGATCCATC |
| Wnt1 Rv | ATACGCGTCAAGGAAAGGTGATAATACC |
| Sfrp1 Fw | GCACTAGTATGGCATGTTGGCTGCTCTG |
| Sfrp1 Rv | ATACGCGTACCTGGGAATCACTATTAAC |
| Tcf7l2 Fw | CCCCTGCTTGATTGAAGTG |
| Tcf7l2 Rv | GGCGGCACAAAATTAAAGAG |
| Lpr5 Fw | AAGACCCTGCTTGAGGACAA |
| Lpr5 Rv | TTGACCTTGTGGACCCTTTC |
| Gsk3β Fw | CTCCTCATGCTCGGATTCA |
| Gsk3β Rv | TGCAGAAGCAGCATTATTGG |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, N.D.S.; Numata, E.D.P.; Mesquita, L.M.d.S.; Dias, P.H.; Vilegas, W.; Gambero, A.; Ribeiro, M.L. Modulatory Effects of Guarana (Paullinia cupana) on Adipogenesis. Nutrients 2017, 9, 635. https://doi.org/10.3390/nu9060635
Lima NDS, Numata EDP, Mesquita LMdS, Dias PH, Vilegas W, Gambero A, Ribeiro ML. Modulatory Effects of Guarana (Paullinia cupana) on Adipogenesis. Nutrients. 2017; 9(6):635. https://doi.org/10.3390/nu9060635
Chicago/Turabian StyleLima, Natália Da Silva, Erica De Paula Numata, Leonardo Mendes de Souza Mesquita, Pollyana Hammoud Dias, Wagner Vilegas, Alessandra Gambero, and Marcelo Lima Ribeiro. 2017. "Modulatory Effects of Guarana (Paullinia cupana) on Adipogenesis" Nutrients 9, no. 6: 635. https://doi.org/10.3390/nu9060635
APA StyleLima, N. D. S., Numata, E. D. P., Mesquita, L. M. d. S., Dias, P. H., Vilegas, W., Gambero, A., & Ribeiro, M. L. (2017). Modulatory Effects of Guarana (Paullinia cupana) on Adipogenesis. Nutrients, 9(6), 635. https://doi.org/10.3390/nu9060635