Diet Hypotheses in Light of the Microbiota Revolution: New Perspectives

{kind=link}
Abstract
:1. Introduction
2. Nutrients and Epidemiology of Allergy
3. Milk and Epidemiology of Allergy
3.1. Breastfeeding
3.2. Unpasteurized Cow’s Milk
4. Dietary Fiber and the Lung Function
5. Dietary Fiber, Short-Chain Fatty Acids, and Susceptibility to Diseases: Lessons Learned from Animal Studies
6. Dietary Fats and Susceptibility to Diseases
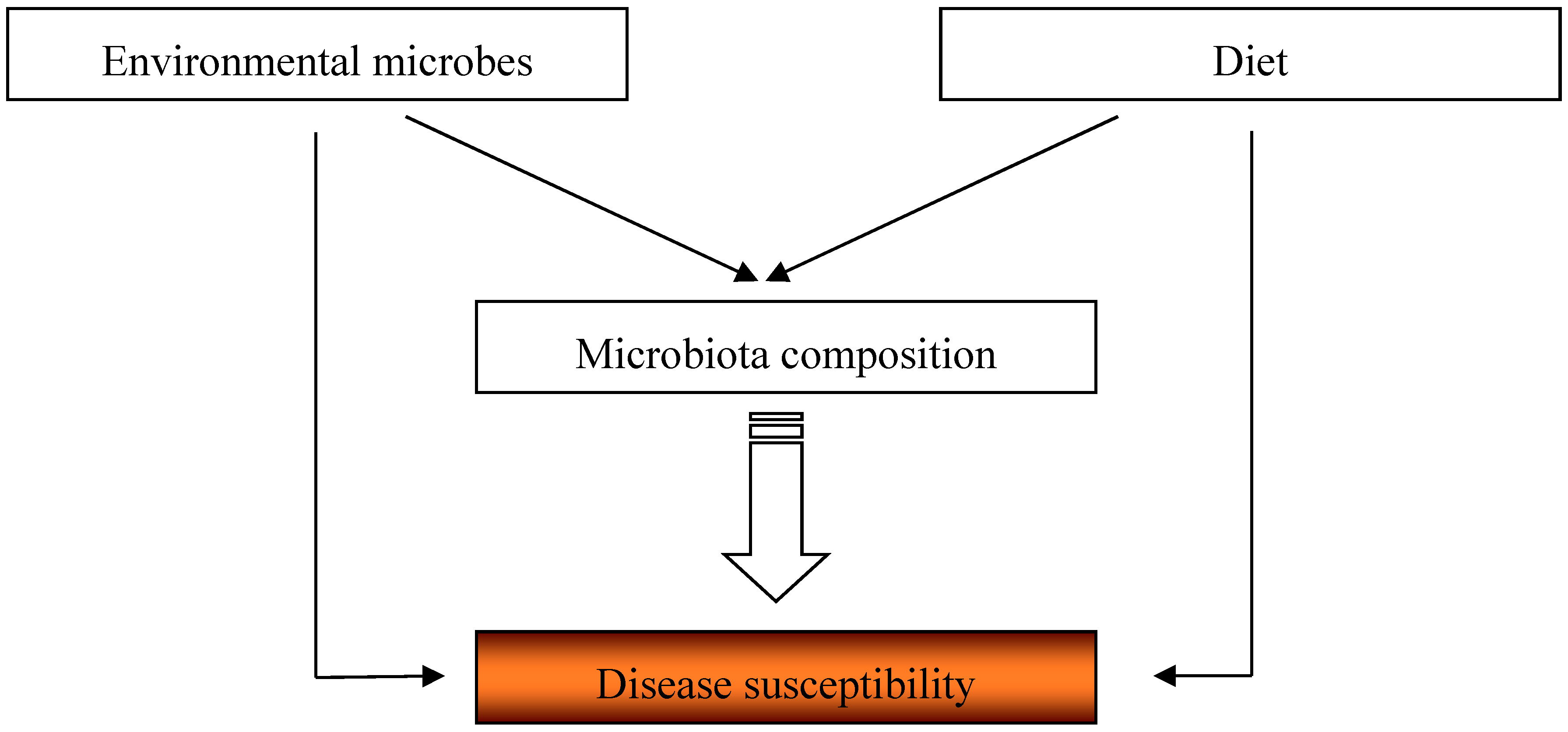
7. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Strachan, D.P. Hay fever, hygiene, and household size. BMJ 1989, 299, 1259–1260. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.A.; Martinelli, R.; Brunet, L.R. Innate immune responses to mycobacteria and the downregulation of atopic responses. Curr. Opin. Allergy Clin. Immunol. 2003, 3, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Von Hertzen, L.; Hanski, I.; Haahtela, T. Natural immunity. Biodiversity loss and inflammatory diseases are two global megatrends that might be related. EMBO Rep. 2011, 12, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Profet, M. The function of allergy: Immunological defense against toxins. Q. Rev. Biol. 1991, 66, 23–62. [Google Scholar] [CrossRef] [PubMed]
- Devereux, G.; Seaton, A. Diet as a risk factor for atopy and asthma. J. Allergy Clin. Immunol. 2005, 115, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Devereux, G. Early life events in asthma—Diet. Pediatr. Pulmonol. 2007, 42, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Ellwood, P.E.; Asher, M.I. Diet and asthma: Looking back, moving forward. Respir. Res. 2009, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and evolution of the western diet: Health implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar] [PubMed]
- Cordain, L.; Watkins, B.A.; Florant, G.L.; Kelher, M.; Rogers, L.; Li, Y. Fatty acid analysis of wild ruminant tissues: Evolutionary implications for reducing diet-related chronic disease. Eur. J. Clin. Nutr. 2002, 56, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Rule, D.C.; Broughton, K.S.; Shellito, S.M.; Maiorano, G. Comparison of muscle fatty acid profiles and cholesterol concentrations of bison, beef cattle, elk, and chicken. J. Anim. Sci. 2002, 80, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Burney, P. A diet rich in sodium may potentiate asthma. Epidemiologic evidence for a new hypothesis. Chest 1987, 91, 143S–148S. [Google Scholar] [CrossRef] [PubMed]
- Javaid, A.; Cushley, M.J.; Bone, M.F. Effect of dietary salt on bronchial reactivity to histamine in asthma. BMJ 1988, 297, 454. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.; Weiss, S.T. Dietary factors and their relation to respiratory symptoms. The second national health and nutrition examination survey. Am. J. Epidemiol. 1990, 132, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Carey, O.J.; Locke, C.; Cookson, J.B. Effect of alterations of dietary sodium on the severity of asthma in men. Thorax 1993, 48, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Pistelli, R.; Forastiere, F.; Corbo, G.M.; Dell’Orco, V.; Brancato, G.; Agabiti, N.; Pizzabiocca, A.; Perucci, C.A. Respiratory symptoms and bronchial responsiveness are related to dietary salt intake and urinary potassium excretion in male children. Eur. Respir. J. 1993, 6, 517–522. [Google Scholar] [PubMed]
- Sparrow, D.; O’Connor, G.T.; Rosner, B.; Weiss, S.T. Methacholine airway responsiveness and 24-hour urine excretion of sodium and potassium. The normative aging study. Am. Rev. Respir. Dis. 1991, 144, 722–725. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, D.; Heimer, D. Effect of dietary sodium on the severity of bronchial asthma. Thorax 1992, 47, 360–362. [Google Scholar] [CrossRef] [PubMed]
- Devereux, G.; Beach, J.R.; Bromly, C.; Avery, A.J.; Ayatollahi, S.M.; Williams, S.M.; Stenton, S.C.; Bourke, S.J.; Hendrick, D.J. Effect of dietary sodium on airways responsiveness and its importance in the epidemiology of asthma: An evaluation in three areas of northern England. Thorax 1995, 50, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Rubin, R.N.; Navon, L.; Cassano, P.A. Relationship of serum antioxidants to asthma prevalence in youth. Am. J. Respir. Crit. Care Med. 2004, 169, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.D.; Welch, A.A.; Bingham, S.A.; Luben, R.N.; Day, N.E.; Khaw, K.T.; Lomas, D.A.; Wareham, N.J. Dietary antioxidants and asthma in adults. Thorax 2006, 61, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Picado, C.; Deulofeu, R.; Lleonart, R.; Agusti, M.; Mullol, J.; Quinto, L.; Torra, M. Dietary micronutrients/antioxidants and their relationship with bronchial asthma severity. Allergy 2001, 56, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Gao, X.; Li, W.; Zhu, Y.; Thompson, P.J. Observational studies on the effect of dietary antioxidants on asthma: A meta-analysis. Respirology 2008, 13, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.; Rowe, B.H.; Ram, F.S. Vitamin C supplementation for asthma. Cochrane Database Syst. Rev. 2001, CD000993. [Google Scholar]
- Fogarty, A.; Lewis, S.A.; Scrivener, S.L.; Antoniak, M.; Pacey, S.; Pringle, M.; Britton, J. Oral magnesium and vitamin C supplements in asthma: A parallel group randomized placebo-controlled trial. Clin. Exp. Allergy 2003, 33, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Troisi, R.J.; Willett, W.C.; Weiss, S.T.; Trichopoulos, D.; Rosner, B.; Speizer, F.E. A prospective study of diet and adult-onset asthma. Am. J. Respir. Crit. Care Med. 1995, 151, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Bodner, C.; Godden, D.; Brown, K.; Little, J.; Ross, S.; Seaton, A. Antioxidant intake and adult-onset wheeze: A case-control study. Eur. Respir. J. 1999, 13, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Phinney, S.D.; Halpern, G.M.; Gershwin, M.E. Asthma mortality: Another opinion—Is it a matter of life and bread? J. Asthma 1993, 30, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Hodge, L.; Peat, J.K.; Salome, C. Increased consumption of polyunsaturated oils may be a cause of increased prevalence of childhood asthma. Aust. N. Z. J. Med. 1994, 24, 727. [Google Scholar] [CrossRef] [PubMed]
- Black, P.N.; Sharpe, S. Dietary fat and asthma: Is there a connection? Eur. Respir. J. 1997, 10, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Dunder, T.; Kuikka, L.; Turtinen, J.; Rasanen, L.; Uhari, M. Diet, serum fatty acids, and atopic diseases in childhood. Allergy 2001, 56, 425–428. [Google Scholar] [CrossRef] [PubMed]
- Bolte, G.; Frye, C.; Hoelscher, B.; Meyer, I.; Wjst, M.; Heinrich, J. Margarine consumption and allergy in children. Am. J. Respir. Crit. Care Med. 2001, 163, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Trak-Fellermeier, M.A.; Brasche, S.; Winkler, G.; Koletzko, B.; Heinrich, J. Food and fatty acid intake and atopic disease in adults. Eur. Respir. J. 2004, 23, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Haby, M.M.; Peat, J.K.; Marks, G.B.; Woolcock, A.J.; Leeder, S.R. Asthma in preschool children: Prevalence and risk factors. Thorax 2001, 56, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Fluge, O.; Omenaas, E.; Eide, G.E.; Gulsvik, A. Fish consumption and respiratory symptoms among young adults in a Norwegian community. Eur. Respir. J. 1998, 12, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Bolte, G.; Kompauer, I.; Fobker, M.; Cullen, P.; Keil, U.; Mutius, E.; Weiland, S.K. Fatty acids in serum cholesteryl esters in relation to asthma and lung function in children. Clin. Exp. Allergy 2006, 36, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Almqvist, C.; Garden, F.; Xuan, W.; Mihrshahi, S.; Leeder, S.R.; Oddy, W.; Webb, K.; Marks, G.B.; Team, C. Omega-3 and omega-6 fatty acid exposure from early life does not affect atopy and asthma at age 5 years. J. Allergy Clin. Immunol. 2007, 119, 1438–1444. [Google Scholar] [CrossRef] [PubMed]
- Takemura, Y.; Sakurai, Y.; Honjo, S.; Tokimatsu, A.; Gibo, M.; Hara, T.; Kusakari, A.; Kugai, N. The relationship between fish intake and the prevalence of asthma: The Tokorozawa childhood asthma and pollinosis study. Prev. Med. 2002, 34, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Broadfield, E.C.; McKeever, T.M.; Whitehurst, A.; Lewis, S.A.; Lawson, N.; Britton, J.; Fogarty, A. A case-control study of dietary and erythrocyte membrane fatty acids in asthma. Clin. Exp. Allergy 2004, 34, 1232–1236. [Google Scholar] [CrossRef] [PubMed]
- Grulee, C.G.; Sanford, H.N. The influence of breast and artificial feeding on infantile eczema. J. Pediatr. 1936, 9, 223–225. [Google Scholar] [CrossRef]
- Saarinen, U.M.; Kajosaari, M.; Backman, A.; Siimes, M.A. Prolonged breast-feeding as prophylaxis for atopic disease. Lancet 1979, 2, 163–166. [Google Scholar] [CrossRef]
- Hide, D.W.; Guyer, B.M. Clinical manifestations of allergy related to breast and cows’ milk feeding. Arch. Dis. Child. 1981, 56, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.L.; Holberg, C.J.; Martinez, F.D.; Morgan, W.J.; Taussig, L.M. Breast feeding and lower respiratory tract illness in the first year of life. Group health medical associates. BMJ 1989, 299, 946–949. [Google Scholar] [CrossRef] [PubMed]
- Dell, S.; To, T. Breastfeeding and asthma in young children: Findings from a population-based study. Arch. Pediatr. Adolesc. Med. 2001, 155, 1261–1265. [Google Scholar] [CrossRef] [PubMed]
- Kull, I.; Wickman, M.; Lilja, G.; Nordvall, S.L.; Pershagen, G. Breast feeding and allergic diseases in infants-a prospective birth cohort study. Arch. Dis. Child. 2002, 87, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Saarinen, U.M.; Kajosaari, M. Breastfeeding as prophylaxis against atopic disease: Prospective follow-up study until 17 years old. Lancet 1995, 346, 1065–1069. [Google Scholar] [CrossRef]
- Tariq, S.M.; Matthews, S.M.; Hakim, E.A.; Stevens, M.; Arshad, S.H.; Hide, D.W. The prevalence of and risk factors for atopy in early childhood: A whole population birth cohort study. J. Allergy Clin. Immunol. 1998, 101, 587–593. [Google Scholar] [CrossRef]
- Oddy, W.H.; Holt, P.G.; Sly, P.D.; Read, A.W.; Landau, L.I.; Stanley, F.J.; Kendall, G.E.; Burton, P.R. Association between breast feeding and asthma in 6 year old children: Findings of a prospective birth cohort study. BMJ 1999, 319, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Scholtens, S.; Wijga, A.H.; Brunekreef, B.; Kerkhof, M.; Hoekstra, M.O.; Gerritsen, J.; Aalberse, R.; de Jongste, J.C.; Smit, H.A. Breast feeding, parental allergy and asthma in children followed for 8 years. The piama birth cohort study. Thorax 2009, 64, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Hide, D.W.; Guyer, B.M. Clinical manifestations of allergy related to breast- and cow’s milk-feeding. Pediatrics 1985, 76, 973–975. [Google Scholar] [PubMed]
- Wright, A.L.; Holberg, C.J.; Taussig, L.M.; Martinez, F.D. Factors influencing the relation of infant feeding to asthma and recurrent wheeze in childhood. Thorax 2001, 56, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Takemura, Y.; Sakurai, Y.; Honjo, S.; Kusakari, A.; Hara, T.; Gibo, M.; Tokimatsu, A.; Kugai, N. Relation between breastfeeding and the prevalence of asthma : The tokorozawa childhood asthma and pollinosis study. Am. J. Epidemiol. 2001, 154, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Sears, M.R.; Greene, J.M.; Willan, A.R.; Taylor, D.R.; Flannery, E.M.; Cowan, J.O.; Herbison, G.P.; Poulton, R. Long-term relation between breastfeeding and development of atopy and asthma in children and young adults: A longitudinal study. Lancet 2002, 360, 901–907. [Google Scholar] [CrossRef]
- Burgess, S.W.; Dakin, C.J.; O’Callaghan, M.J. Breastfeeding does not increase the risk of asthma at 14 years. Pediatrics 2006, 117, e787–e792. [Google Scholar] [CrossRef] [PubMed]
- Gdalevich, M.; Mimouni, D.; Mimouni, M. Breast-feeding and the risk of bronchial asthma in childhood: A systematic review with meta-analysis of prospective studies. J. Pediatr. 2001, 139, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Gdalevich, M.; Mimouni, D.; David, M.; Mimouni, M. Breast-feeding and the onset of atopic dermatitis in childhood: A systematic review and meta-analysis of prospective studies. J. Am. Acad. Dermatol. 2001, 45, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Mimouni Bloch, A.; Mimouni, D.; Mimouni, M.; Gdalevich, M. Does breastfeeding protect against allergic rhinitis during childhood? A meta-analysis of prospective studies. Acta Paediatr. 2002, 91, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Dogaru, C.M.; Nyffenegger, D.; Pescatore, A.M.; Spycher, B.D.; Kuehni, C.E. Breastfeeding and childhood asthma: Systematic review and meta-analysis. Am. J. Epidemiol. 2014, 179, 1153–1167. [Google Scholar] [CrossRef] [PubMed]
- Peat, J.K.; Allen, J.; Oddy, W.; Webb, K. Breastfeeding and asthma: Appraising the controversy. Pediatr. Pulmonol. 2003, 35, 331–334. [Google Scholar] [CrossRef] [PubMed]
- Oddy, W.H.; Peat, J.K. Breastfeeding, asthma, and atopic disease: An epidemiological review of the literature. J. Hum. Lact. 2003, 19, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Rogier, E.W.; Frantz, A.L.; Bruno, M.E.; Wedlund, L.; Cohen, D.A.; Stromberg, A.J.; Kaetzel, C.S. Secretory antibodies in breast milk promote long-term intestinal homeostasis by regulating the gut microbiota and host gene expression. Proc. Natl. Acad. Sci. USA 2014, 111, 3074–3079. [Google Scholar] [CrossRef] [PubMed]
- Chichlowski, M.; German, J.B.; Lebrilla, C.B.; Mills, D.A. The influence of milk oligosaccharides on microbiota of infants: Opportunities for formulas. Annu. Rev. Food Sci. Technol. 2011, 2, 331–351. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, L.; Langa, S.; Martin, V.; Maldonado, A.; Jimenez, E.; Martin, R.; Rodriguez, J.M. The human milk microbiota: Origin and potential roles in health and disease. Pharmacol. Res. 2013, 69, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kaetzel, C.S. Cooperativity among secretory IgA, the polymeric immunoglobulin receptor, and the gut microbiota promotes host-microbial mutualism. Immunol. Lett. 2014, 162, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Rogier, E.W.; Frantz, A.L.; Bruno, M.E.; Wedlund, L.; Cohen, D.A.; Stromberg, A.J.; Kaetzel, C.S. Lessons from mother: Long-term impact of antibodies in breast milk on the gut microbiota and intestinal immune system of breastfed offspring. Gut Microbes 2014, 5, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Julia, V.; Macia, L.; Dombrowicz, D. The impact of diet on asthma and allergic diseases. Nat. Rev. Immunol. 2015, 15, 308–322. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.A.; Iyengar, R.S. Breast milk, microbiota, and intestinal immune homeostasis. Pediatr. Res. 2015, 77, 220–228. [Google Scholar] [PubMed]
- Perkin, M.R.; Strachan, D.P. Which aspects of the farming lifestyle explain the inverse association with childhood allergy? J. Allergy Clin. Immunol. 2006, 117, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Riedler, J.; Braun-Fahrlander, C.; Eder, W.; Schreuer, M.; Waser, M.; Maisch, S.; Carr, D.; Schierl, R.; Nowak, D.; von Mutius, E.; et al. Exposure to farming in early life and development of asthma and allergy: A cross-sectional survey. Lancet 2001, 358, 1129–1133. [Google Scholar] [CrossRef]
- Waser, M.; Michels, K.B.; Bieli, C.; Floistrup, H.; Pershagen, G.; von Mutius, E.; Ege, M.; Riedler, J.; Schram-Bijkerk, D.; Brunekreef, B.; et al. Inverse association of farm milk consumption with asthma and allergy in rural and suburban populations across Europe. Clin. Exp. Allergy 2007, 37, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Wickens, K.; Lane, J.M.; Fitzharris, P.; Siebers, R.; Riley, G.; Douwes, J.; Smith, T.; Crane, J. Farm residence and exposures and the risk of allergic diseases in New Zealand children. Allergy 2002, 57, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Loss, G.; Apprich, S.; Waser, M.; Kneifel, W.; Genuneit, J.; Buchele, G.; Weber, J.; Sozanska, B.; Danielewicz, H.; Horak, E.; et al. The protective effect of farm milk consumption on childhood asthma and atopy: The Gabriela study. J. Allergy Clin. Immunol. 2011, 128, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Van Neerven, R.J.; Knol, E.F.; Heck, J.M.; Savelkoul, H.F. Which factors in raw cow’s milk contribute to protection against allergies? J. Allergy Clin. Immunol. 2012, 130, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Brick, T.; Schober, Y.; Bocking, C.; Pekkanen, J.; Genuneit, J.; Loss, G.; Dalphin, J.C.; Riedler, J.; Lauener, R.; Nockher, W.A.; et al. Omega-3 fatty acids contribute to the asthma-protective effect of unprocessed cow’s milk. J. Allergy Clin. Immunol. 2016, 137, 1699–1706. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, B.; Pfaffl, M.W.; Dumpler, J.; von Mutius, E.; Ege, M.J. MicroRNA in native and processed cow’s milk and its implication for the farm milk effect on asthma. J. Allergy Clin. Immunol. 2016, 137, 1893–1895. [Google Scholar] [CrossRef] [PubMed]
- Quigley, L.; O’Sullivan, O.; Stanton, C.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. The complex microbiota of raw milk. FEMS Microbiol. Rev. 2013, 37, 664–698. [Google Scholar] [CrossRef] [PubMed]
- Braun-Fahrlander, C.; von Mutius, E. Can farm milk consumption prevent allergic diseases? Clin. Exp. Allergy 2011, 41, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Halnes, I.; Baines, K.J.; Berthon, B.S.; MacDonald-Wicks, L.K.; Gibson, P.G.; Wood, L.G. Soluble fibre meal challenge reduces airway inflammation and expression of gpr43 and gpr41 in asthma. Nutrients 2017, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Kan, H.; Stevens, J.; Heiss, G.; Rose, K.M.; London, S.J. Dietary fiber, lung function, and chronic obstructive pulmonary disease in the atherosclerosis risk in communities study. Am. J. Epidemiol. 2008, 167, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Varraso, R.; Willett, W.C.; Camargo, C.A., Jr. Prospective study of dietary fiber and risk of chronic obstructive pulmonary disease among US women and men. Am. J. Epidemiol. 2010, 171, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Berthon, B.S.; Macdonald-Wicks, L.K.; Gibson, P.G.; Wood, L.G. Investigation of the association between dietary intake, disease severity and airway inflammation in asthma. Respirology 2013, 18, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Root, M.M.; Houser, S.M.; Anderson, J.J.; Dawson, H.R. Healthy eating index 2005 and selected macronutrients are correlated with improved lung function in humans. Nutr. Res. 2014, 34, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Roediger, W.E. The colonic epithelium in ulcerative colitis: An energy-deficiency disease? Lancet 1980, 2, 712–715. [Google Scholar] [CrossRef]
- Gilbert, K.M.; Weigle, W.O. Th1 cell anergy and blockade in G1a phase of the cell cycle. J. Immunol. 1993, 151, 1245–1254. [Google Scholar] [PubMed]
- Siavoshian, S.; Blottiere, H.M.; Bentouimou, N.; Cherbut, C.; Galmiche, J.P. Butyrate enhances major histocompatibility complex class I, HLA-DR and ICAM-1 antigen expression on differentiated human intestinal epithelial cells. Eur. J. Clin. Investig. 1996, 26, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Bohmig, G.A.; Krieger, P.M.; Saemann, M.D.; Wenhardt, C.; Pohanka, E.; Zlabinger, G.J. N-butyrate downregulates the stimulatory function of peripheral blood-derived antigen-presenting cells: A potential mechanism for modulating T-cell responses by short-chain fatty acids. Immunology 1997, 92, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Segain, J.P.; Raingeard de la Bletiere, D.; Bourreille, A.; Leray, V.; Gervois, N.; Rosales, C.; Ferrier, L.; Bonnet, C.; Blottiere, H.M.; Galmiche, J.P. Butyrate inhibits inflammatory responses through NFkB inhibition: Implications for Crohn’s disease. Gut 2000, 47, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Tedelind, S.; Westberg, F.; Kjerrulf, M.; Vidal, A. Anti-inflammatory properties of the short-chain fatty acids acetate and propionate: A study with relevance to inflammatory bowel disease. World J. Gastroenterol. 2007, 13, 2826–2832. [Google Scholar] [PubMed]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.Y.; Lannoy, V.; Decobecq, M.E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Van Damme, J.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.; Shim, R.; Robert, R.; et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly, Y.M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. Gpr109a is a G-protein-coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of GPR109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; Ian McKenzie, C.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors gpr43 and gpr109a facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Vuillermin, P.J.; Goverse, G.; Vinuesa, C.G.; Mebius, R.E.; Macia, L.; Mackay, C.R. Dietary fiber and bacterial SCFA enhance oral tolerance and protect against food allergy through diverse cellular pathways. Cell. Rep. 2016, 15, 2809–2824. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Von Mutius, E.; Schwartz, J.; Neas, L.M.; Dockery, D.; Weiss, S.T. Relation of body mass index to asthma and atopy in children: The national health and nutrition examination study III. Thorax 2001, 56, 835–838. [Google Scholar] [CrossRef] [PubMed]
- Gilliland, F.D.; Berhane, K.; Islam, T.; McConnell, R.; Gauderman, W.J.; Gilliland, S.S.; Avol, E.; Peters, J.M. Obesity and the risk of newly diagnosed asthma in school-age children. Am. J. Epidemiol. 2003, 158, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Flaherman, V.; Rutherford, G.W. A meta-analysis of the effect of high weight on asthma. Arch. Dis. Child. 2006, 91, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Visness, C.M.; London, S.J.; Daniels, J.L.; Kaufman, J.S.; Yeatts, K.B.; Siega-Riz, A.M.; Calatroni, A.; Zeldin, D.C. Association of childhood obesity with atopic and nonatopic asthma: Results from the national health and nutrition examination survey 1999–2006. J. Asthma 2010, 47, 822–829. [Google Scholar] [CrossRef] [PubMed]
- Johnston, R.A.; Zhu, M.; Rivera-Sanchez, Y.M.; Lu, F.L.; Theman, T.A.; Flynt, L.; Shore, S.A. Allergic airway responses in obese mice. Am. J. Respir. Crit. Care Med. 2007, 176, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Calixto, M.C.; Lintomen, L.; Schenka, A.; Saad, M.J.; Zanesco, A.; Antunes, E. Obesity enhances eosinophilic inflammation in a murine model of allergic asthma. Br. J. Pharmacol. 2010, 159, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Dietze, J.; Bocking, C.; Heverhagen, J.T.; Voelker, M.N.; Renz, H. Obesity lowers the threshold of allergic sensitization and augments airway eosinophilia in a mouse model of asthma. Allergy 2012, 67, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Lee, H.J.; Chang, Y.J.; Pichavant, M.; Shore, S.A.; Fitzgerald, K.A.; Iwakura, Y.; Israel, E.; Bolger, K.; Faul, J.; et al. Interleukin-17-producing innate lymphoid cells and the NLRP3 inflammasome facilitate obesity-associated airway hyperreactivity. Nat. Med. 2014, 20, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Sideleva, O.; Dixon, A.E. The many faces of asthma in obesity. J. Cell. Biochem. 2014, 115, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Baffi, C.W.; Winnica, D.E.; Holguin, F. Asthma and obesity: Mechanisms and clinical implications. Asthma Res. Pract. 2015, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Leiria, L.O.; Martins, M.A.; Saad, M.J. Obesity and asthma: Beyond TH2 inflammation. Metabolism 2015, 64, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Shore, S.A. Obesity, asthma, and the microbiome. Physiology 2016, 31, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.G.; Garg, M.L.; Gibson, P.G. A high-fat challenge increases airway inflammation and impairs bronchodilator recovery in asthma. J. Allergy Clin. Immunol. 2011, 127, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Myles, I.A.; Fontecilla, N.M.; Janelsins, B.M.; Vithayathil, P.J.; Segre, J.A.; Datta, S.K. Parental dietary fat intake alters offspring microbiome and immunity. J. Immunol. 2013, 191, 3200–3209. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, K.L. The science of fatty acids and inflammation. Adv. Nutr. 2015, 6, 293S–301S. [Google Scholar] [CrossRef] [PubMed]
- Bilal, S.; Haworth, O.; Wu, L.; Weylandt, K.H.; Levy, B.D.; Kang, J.X. Fat-1 transgenic mice with elevated omega-3 fatty acids are protected from allergic airway responses. Biochim. Biophys. Acta 2011, 1812, 1164–1169. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, A.; Hamazaki, T.; Ohshita, A.; Kohno, N.; Sakai, K.; Zhao, G.D.; Katayama, H.; Hiwada, K. Effect of aerosolized docosahexaenoic acid in a mouse model of atopic asthma. Int. Arch. Allergy Immunol. 2000, 123, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.; Fortin, S.; Cantin, A.M.; Rousseau, E. Docosahexaenoic acid derivative prevents inflammation and hyperreactivity in lung: Implication of PKC-potentiated inhibitory protein for heterotrimeric myosin light chain phosphatase of 17 Kd in asthma. Am. J. Respir. Cell. Mol. Biol. 2011, 45, 366–375. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wypych, T.P.; Marsland, B.J. Diet Hypotheses in Light of the Microbiota Revolution: New Perspectives. Nutrients 2017, 9, 537. https://doi.org/10.3390/nu9060537
Wypych TP, Marsland BJ. Diet Hypotheses in Light of the Microbiota Revolution: New Perspectives. Nutrients. 2017; 9(6):537. https://doi.org/10.3390/nu9060537
Chicago/Turabian StyleWypych, Tomasz P., and Benjamin J. Marsland. 2017. "Diet Hypotheses in Light of the Microbiota Revolution: New Perspectives" Nutrients 9, no. 6: 537. https://doi.org/10.3390/nu9060537
APA StyleWypych, T. P., & Marsland, B. J. (2017). Diet Hypotheses in Light of the Microbiota Revolution: New Perspectives. Nutrients, 9(6), 537. https://doi.org/10.3390/nu9060537