Suppression of Oxidative Stress and NFκB/MAPK Signaling by Lyophilized Black Raspberries for Esophageal Cancer Prevention in Rats



Abstract
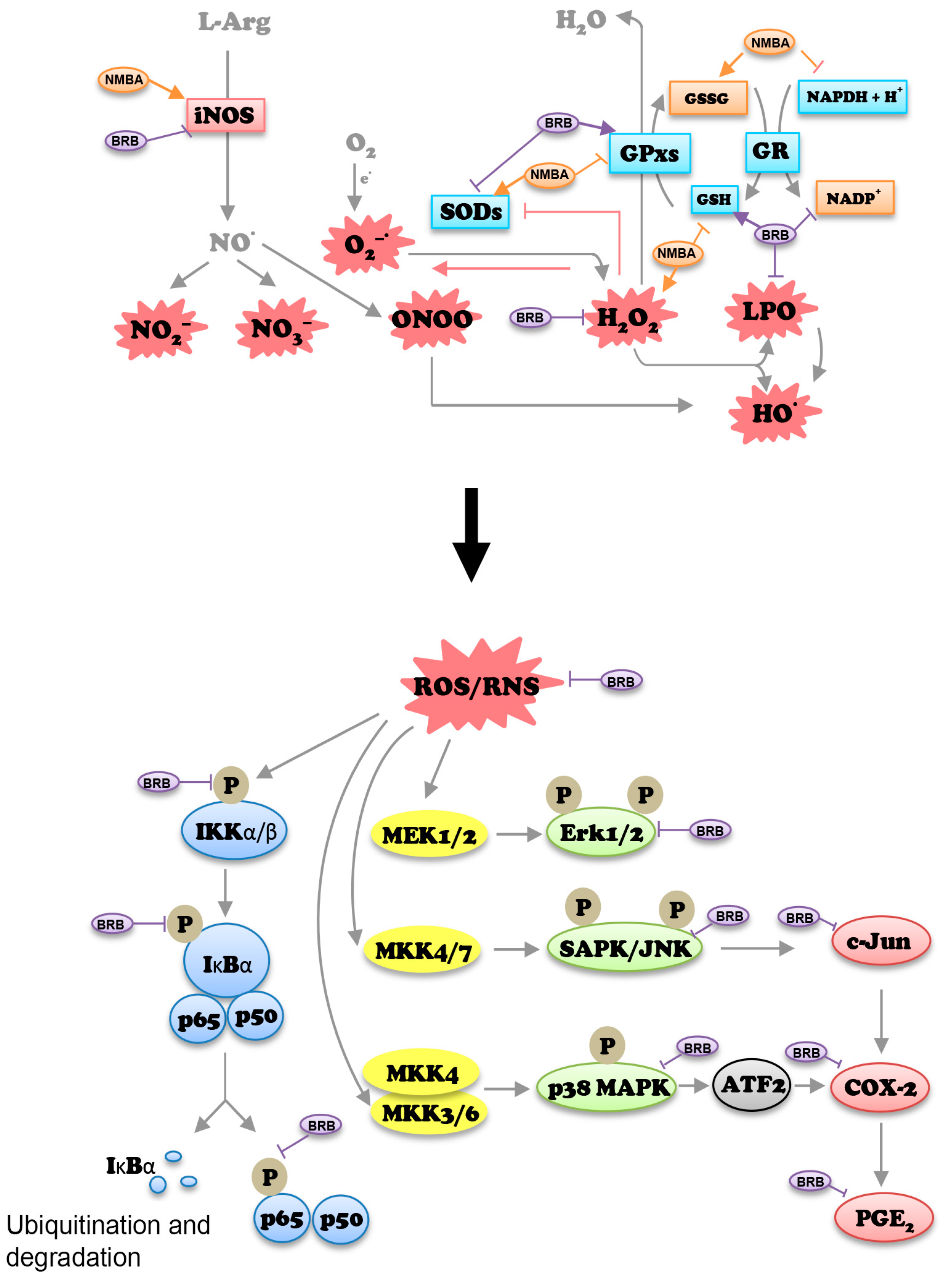
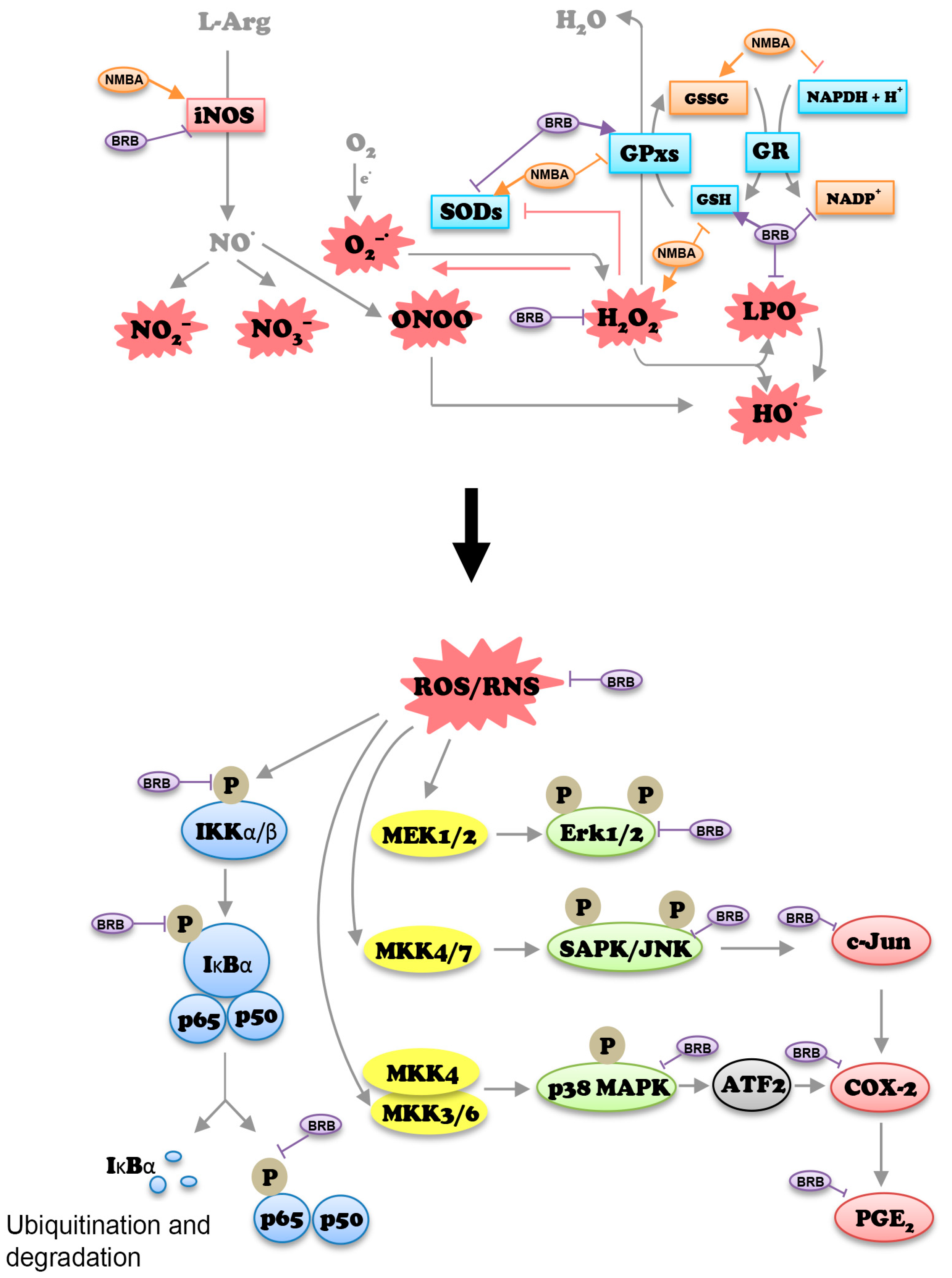
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. In Vivo Model of ESCC
2.3. Immunohistochemistry
2.4. Real-Time RT-PCR
2.5. Western Blot Analysis
2.6. Measurements of H2O2 and LPO
2.7. GPx and SOD2 Activity Assay
2.8. GSSG/GSH and NADP+/NADPH Ratios
2.9. Statistical Analysis
3. Results
3.1. BRB Inhibits NMBA-Induced Tumor Development
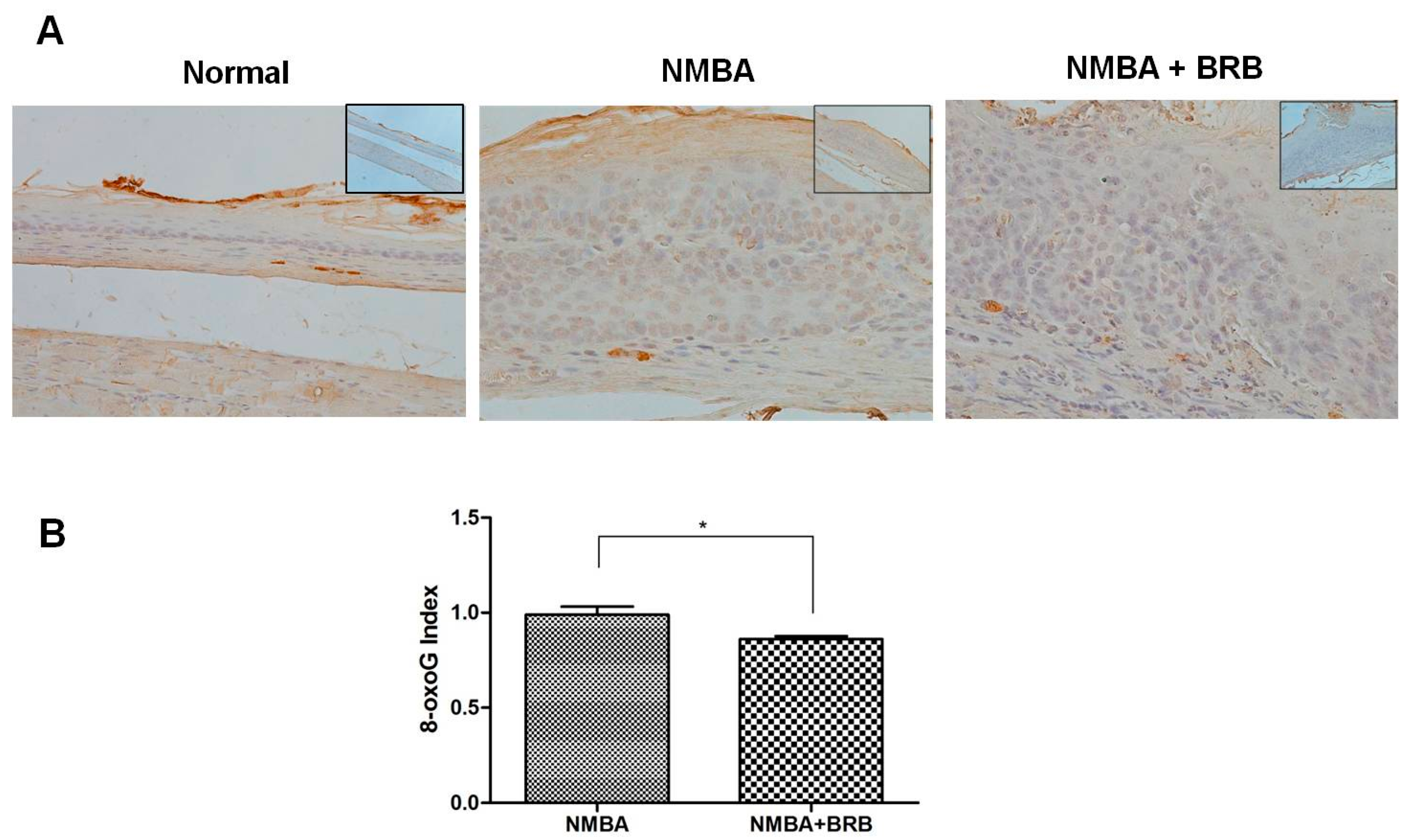
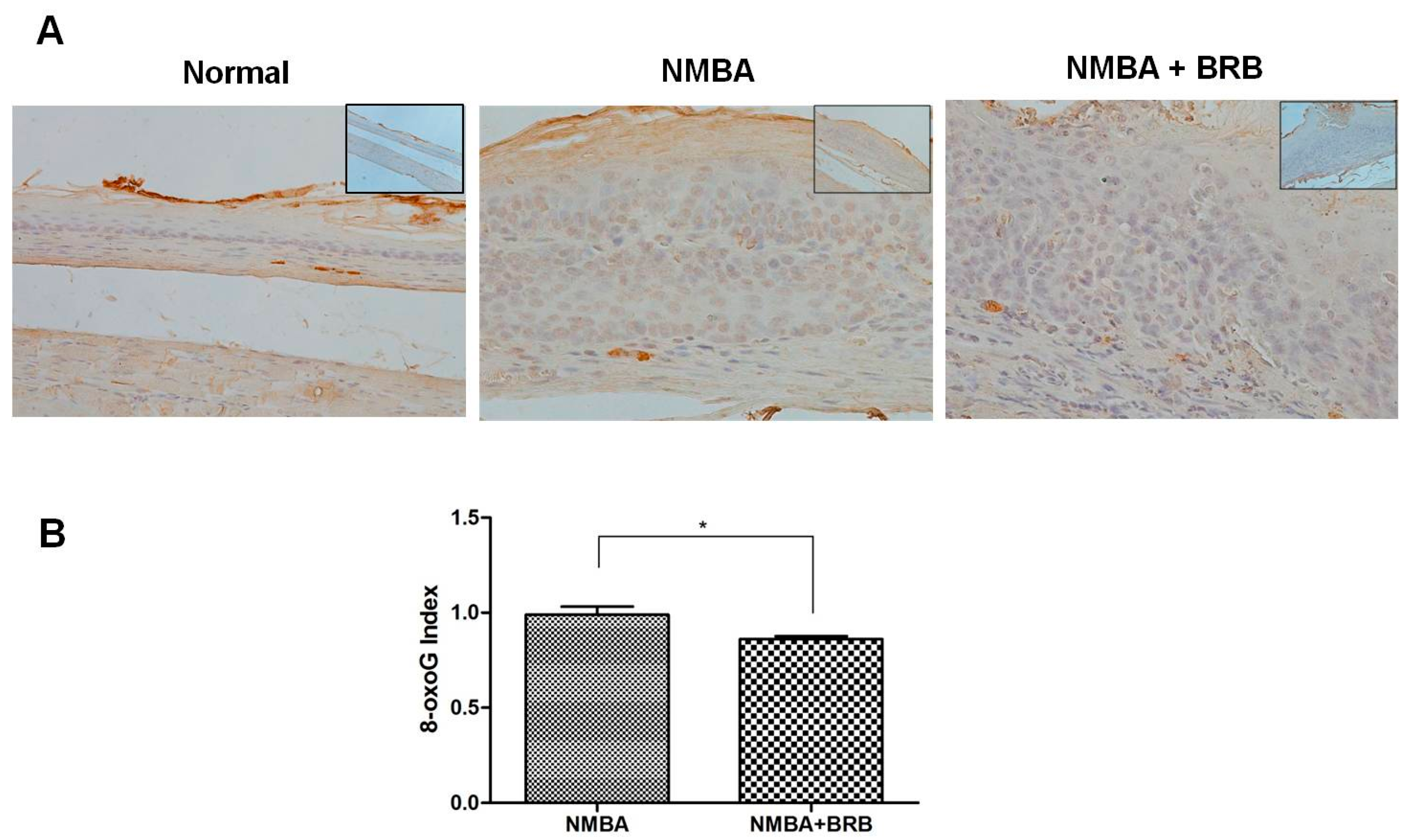
3.2. BRB Decreases Level of 8-OxoG
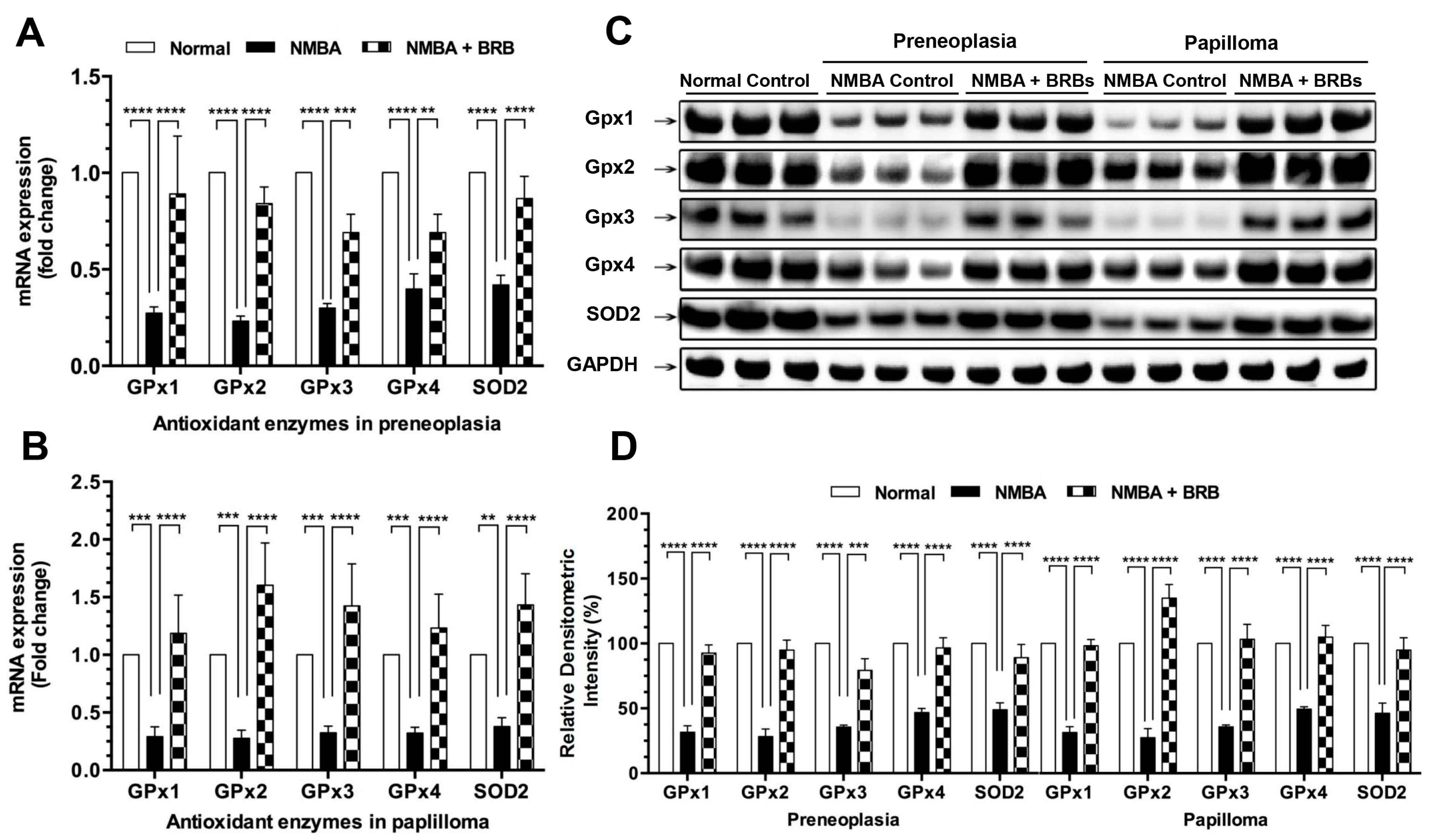
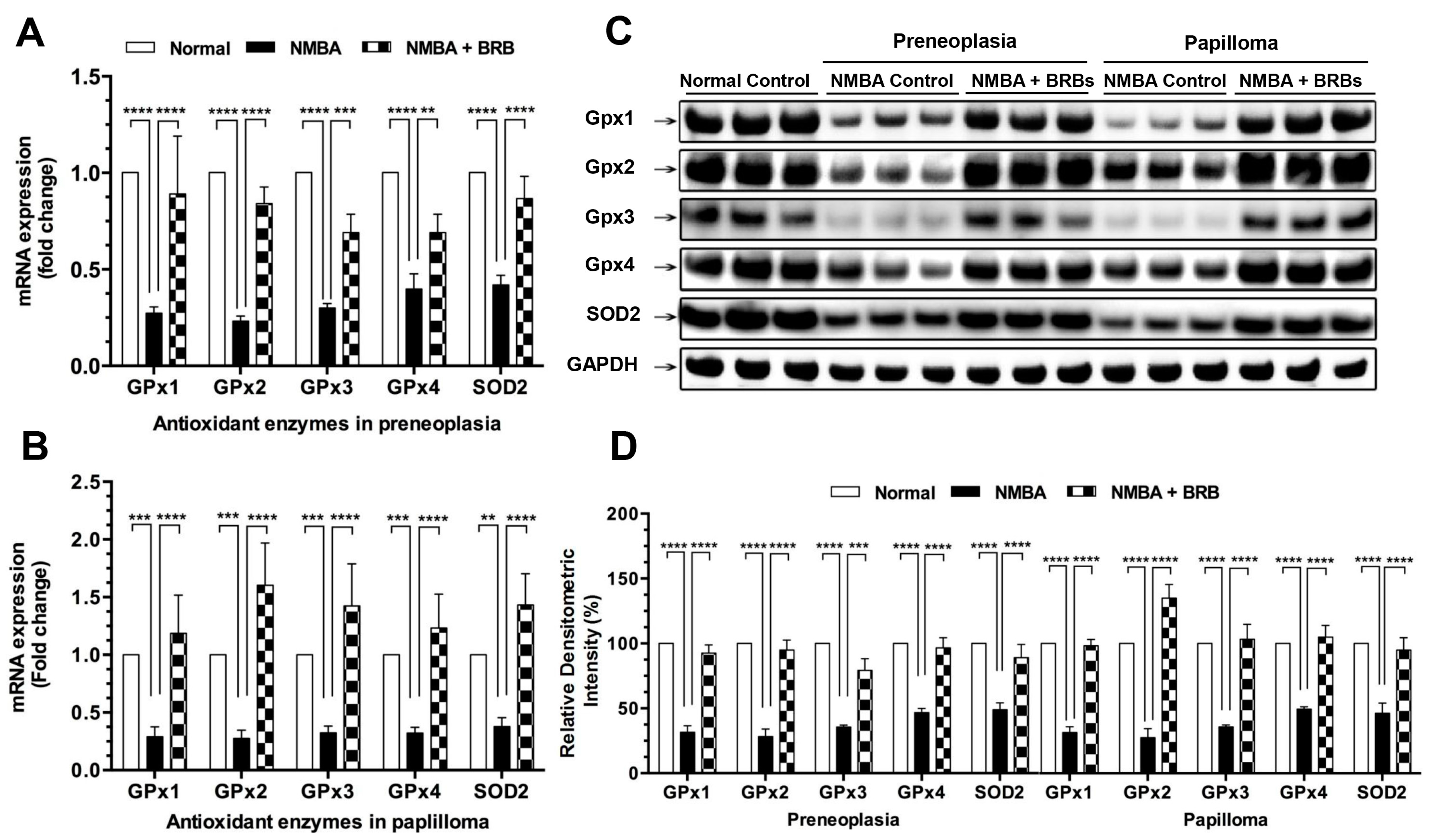
3.3. BRB Increases Expression Levels of GPx and SOD2
3.4. BRB Reduces H2O2 and LPO Concentrations
3.5. BRB Enhances GPx and SOD2 Activities
3.6. BRB Reduces GSSG/GSH and NADP+/NADPH Ratios
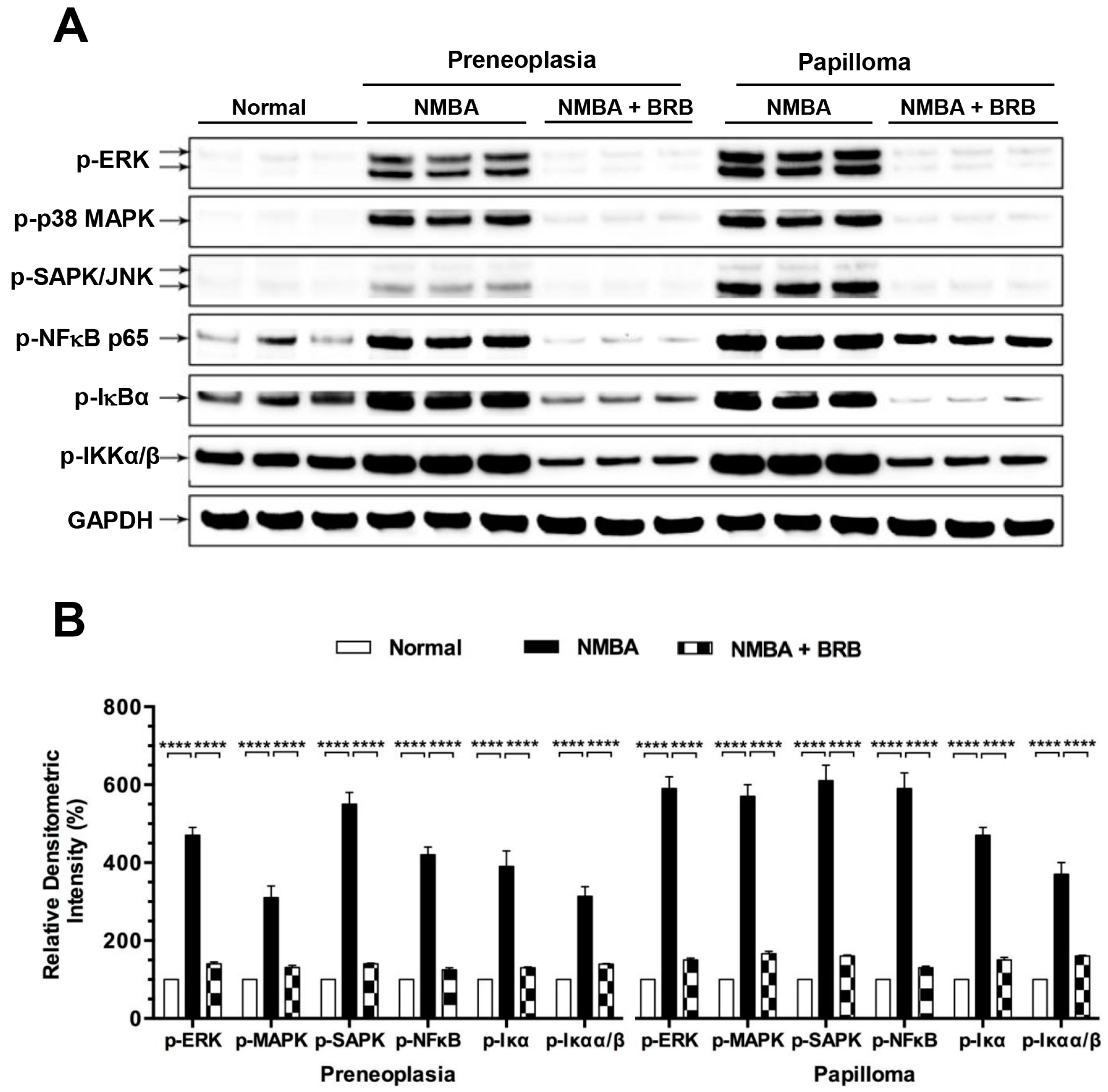
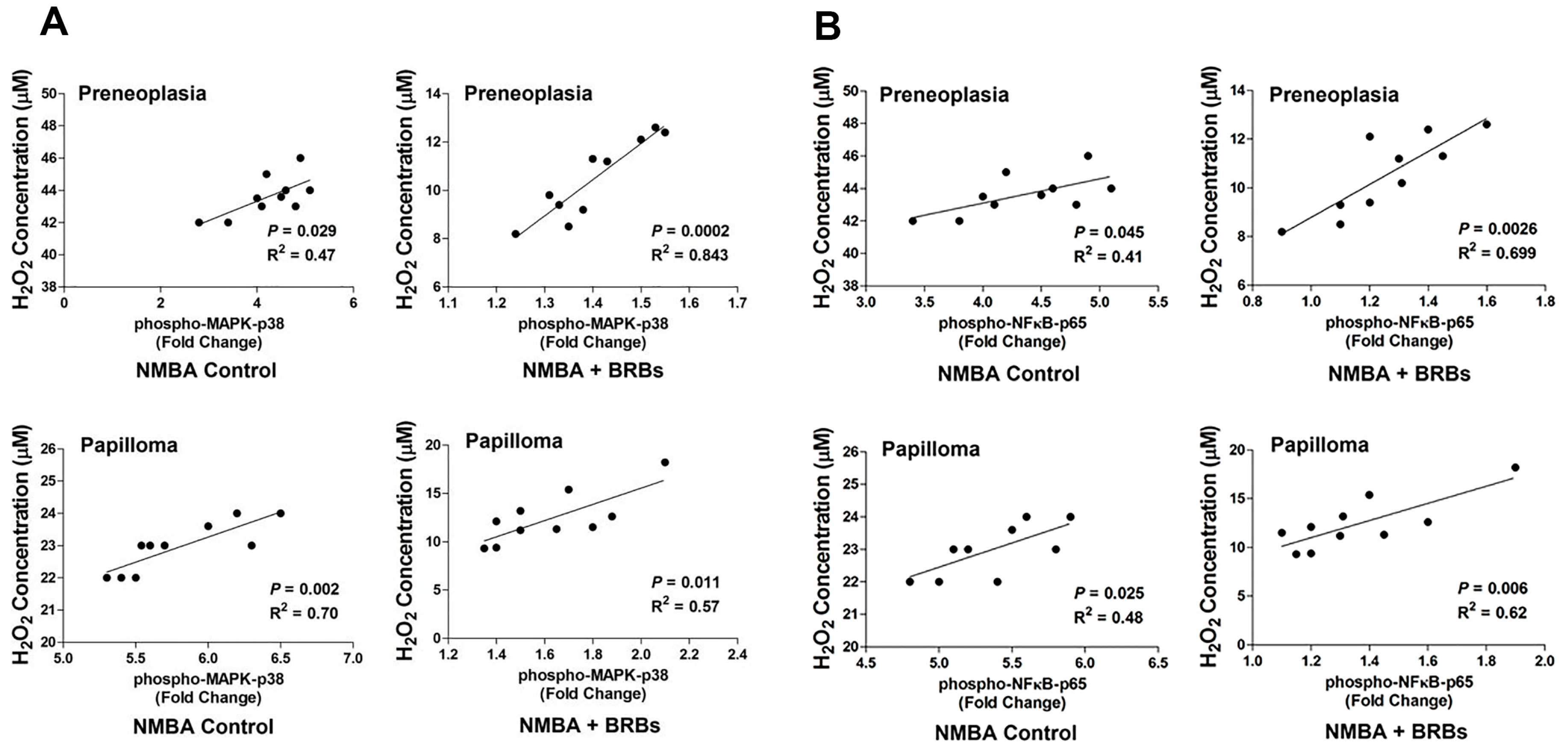
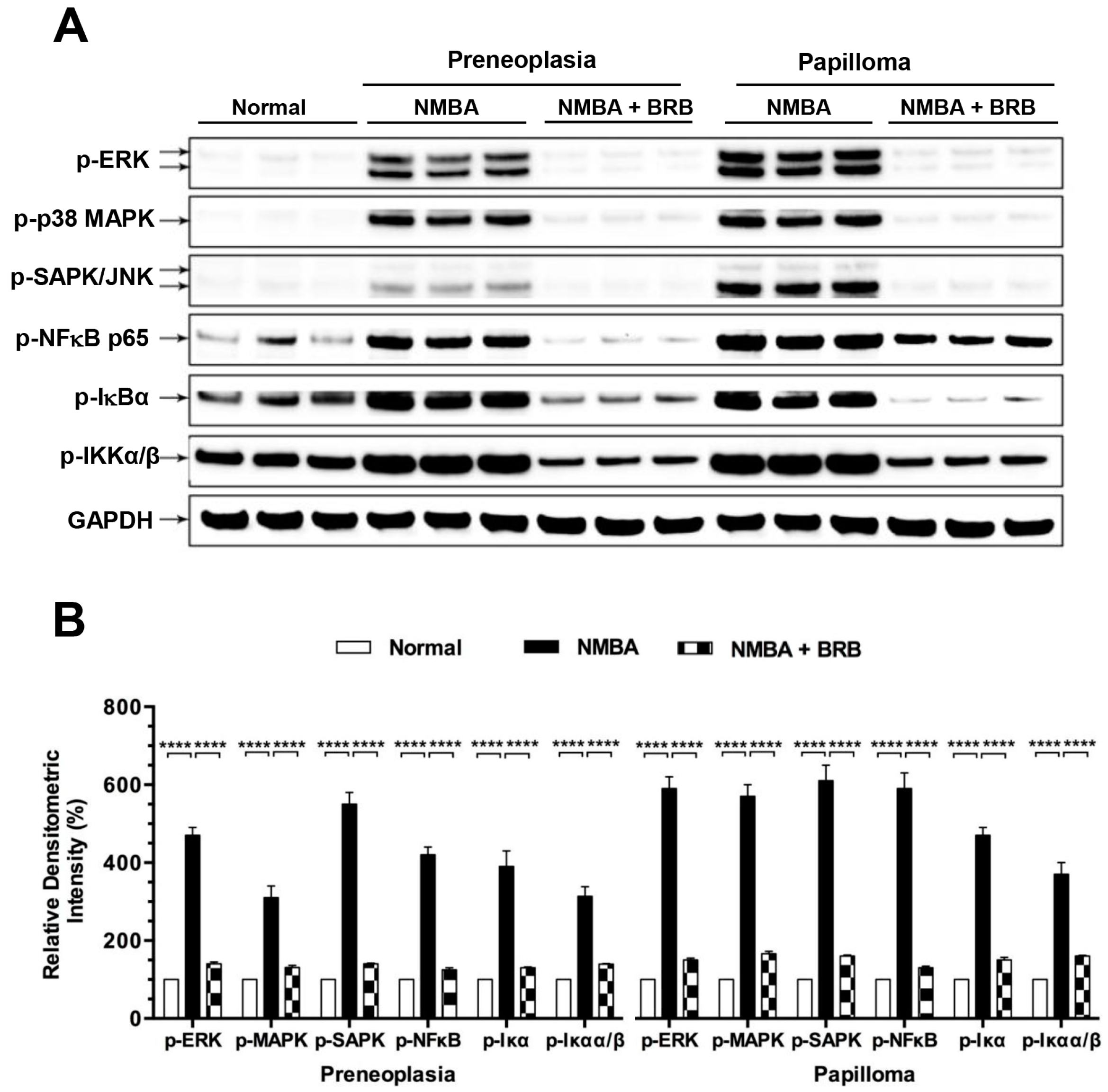
3.7. BRB Suppressed Activation of NFκB and MAPK Signaling
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Trivers, K.F.; Sabatino, S.A.; Stewart, S.L. Trends in esophageal cancer incidence by histology, United States, 1998–2003. Int. J. Cancer 2008, 123, 1422–1428. [Google Scholar] [PubMed]
- Yamashita, K.; Upadhyay, S.; Osada, M.; Hoque, M.O.; Xiao, Y.; Mori, M.; Sato, F.; Meltzer, S.J.; Sidransky, D. Pharmacologic unmasking of epigenetically silenced tumor suppressor genes in esophageal squamous cell carcinoma. Cancer Cell 2002, 2, 485–495. [Google Scholar] [PubMed]
- Prabhu, A.; Obi, K.O.; Rubenstein, J.H. The synergistic effects of alcohol and tobacco consumption on the risk of esophageal squamous cell carcinoma: A meta-analysis. Am. J. Gastroenterol. 2014, 109, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Chestnov, O. Who Report on the Global Tobacco Epidemic, 2013: Enforcing Bans on Tobacco Advertising, Promotion and Sponsorship; World Health Orgnization: Geneva, Switzerland, 2013; pp. 11–89. [Google Scholar]
- Kresty, L.A.; Morse, M.A.; Morgan, C.; Carlton, P.S.; Lu, J.; Gupta, A.; Blackwood, M.; Stoner, G.D. Chemoprevention of esophageal tumorigenesis by dietary administration of lyophilized black raspberries. Cancer Res. 2001, 61, 6112–6119. [Google Scholar] [PubMed]
- Chen, T.; Rose, M.E.; Hwang, H.; Nines, R.G.; Stoner, G.D. Black raspberries inhibit N-nitrosomethylbenzylamine (NMBA)-induced angiogenesis in rat esophagus parallel to the suppression of COX-2 and INOS. Carcinogenesis 2006, 27, 2301–2307. [Google Scholar] [PubMed]
- Chen, T.; Hwang, H.; Rose, M.E.; Nines, R.G.; Stoner, G.D. Chemopreventive properties of black raspberries in N-nitrosomethylbenzylamine-induced rat esophageal tumorigenesis: Down-regulation of cyclooxygenase-2, inducible nitric oxide synthase, and c-jun. Cancer Res. 2006, 66, 2853–2859. [Google Scholar] [CrossRef] [PubMed]
- Stoner, G.D.; Wang, L.S.; Zikri, N.; Chen, T.; Hecht, S.S.; Huang, C.; Sardo, C.; Lechner, J.F. Cancer prevention with freeze-dried berries and berry components. Semin. Cancer Biol. 2007, 17, 403–410. [Google Scholar] [PubMed]
- Shi, N.; Jin, F.; Zhang, X.; Clinton, S.K.; Pan, Z.; Chen, T. Overexpression of human beta-defensin 2 promotes growth and invasion during esophageal carcinogenesis. Oncotarget 2014, 5, 11333–11344. [Google Scholar] [CrossRef] [PubMed]
- Stoner, G.D. Foodstuffs for preventing cancer: The preclinical and clinical development of berries. Cancer Prev. Res. (Phila.) 2009, 2, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.S.; Hecht, S.S.; Carmella, S.G.; Yu, N.; Larue, B.; Henry, C.; McIntyre, C.; Rocha, C.; Lechner, J.F.; Stoner, G.D. Anthocyanins in black raspberries prevent esophageal tumors in rats. Cancer Prev. Res. (Phila.) 2009, 2, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517S–520S. [Google Scholar] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Klaunig, J.E.; Xu, Y.; Isenberg, J.S.; Bachowski, S.; Kolaja, K.L.; Jiang, J.; Stevenson, D.E.; Walborg, E.F., Jr. The role of oxidative stress in chemical carcinogenesis. Environ. Health Perspect. 1998, 106, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Mena, S.; Ortega, A.; Estrela, J.M. Oxidative stress in environmental-induced carcinogenesis. Mutat. Res. 2009, 674, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.; Xie, Q.W. Regulation of biosynthesis of nitric oxide. J. Biol. Chem. 1994, 269, 13725–13728. [Google Scholar] [PubMed]
- Chen, T.; Nines, R.G.; Peschke, S.M.; Kresty, L.A.; Stoner, G.D. Chemopreventive effects of a selective nitric oxide synthase inhibitor on carcinogen-induced rat esophageal tumorigenesis. Cancer Res. 2004, 64, 3714–3717. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.T.; Hsieh, C.C.; Wu, C.T.; Yen, T.C.; Lin, P.Y.; Chen, W.C.; Chen, M.F. 1α,25-dihydroxyvitamin D3 inhibits esophageal squamous cell carcinoma progression by reducing IL6 signaling. Mol. Cancer Ther. 2015, 14, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Kryston, T.B.; Georgiev, A.B.; Pissis, P.; Georgakilas, A.G. Role of oxidative stress and DNA damage in human carcinogenesis. Mutat. Res. 2011, 711, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Altieri, F.; Grillo, C.; Maceroni, M.; Chichiarelli, S. DNA damage and repair: From molecular mechanisms to health implications. Antioxid. Redox Signal. 2008, 10, 891–937. [Google Scholar] [CrossRef] [PubMed]
- Diakowska, D.; Lewandowski, A.; Kopec, W.; Diakowski, W.; Chrzanowska, T. Oxidative DNA damage and total antioxidant status in serum of patients with esophageal squamous cell carcinoma. Hepatogastroenterology 2007, 54, 1701–1704. [Google Scholar] [PubMed]
- Nordberg, J.; Arner, E.S. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic. Biol. Med. 2001, 31, 1287–1312. [Google Scholar] [CrossRef]
- Miguel, F.; Augusto, A.C.; Gurgueira, S.A. Effect of acute vs chronic H2O2-induced oxidative stress on antioxidant enzyme activities. Free Radic. Res. 2009, 43, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Behrend, L.; Henderson, G.; Zwacka, R.M. Reactive oxygen species in oncogenic transformation. Biochem. Soc. Trans. 2003, 31, 1441–1444. [Google Scholar] [CrossRef] [PubMed]
- Klaunig, J.E.; Kamendulis, L.M.; Hocevar, B.A. Oxidative stress and oxidative damage in carcinogenesis. Toxicol. Pathol. 2010, 38, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Yan, F.; Qian, J.; Guo, M.; Zhang, H.; Tang, X.; Chen, F.; Stoner, G.D.; Wang, X. Randomized phase II trial of lyophilized strawberries in patients with dysplastic precancerous lesions of the esophagus. Cancer Prev. Res. (Phila.) 2012, 5, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Ye, W.; Shao, Q.; Qi, Y.; Zhang, M.; Liang, J. Prognostic significance of XIAP and NF-κB expression in esophageal carcinoma with postoperative radiotherapy. World J. Surg. Oncol. 2013, 11, 288. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Stoner, G.D. Inducible nitric oxide synthase expression in n-nitrosomethylbenzylamine (NMBA)-induced rat esophageal tumorigenesis. Mol. Carcinog. 2004, 40, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Yang, W.K.; Li, X.Y.; Lin, T.T.; Lun, Y.N.; Lin, F.; Lv, S.W.; Yan, G.L.; Liu, J.Q.; Shen, J.C.; et al. A trifunctional enzyme with glutathione S-transferase, glutathione peroxidase and superoxide dismutase activity. Biochim. Biophys. Acta 2008, 1780, 869–872. [Google Scholar] [CrossRef] [PubMed]
- Flohe, L.; Otting, F. Superoxide dismutase assays. Methods Enzymol. 1984, 105, 93–104. [Google Scholar] [PubMed]
- Zhang, Z.Q.; Yu, J.; Stanton, R.C. A method for determination of pyridine nucleotides using a single extract. Anal. Biochem. 2000, 285, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Mu, Y.; Yan, G.; Liu, J.; Shen, J.; Luo, G. Antioxidant enzyme mimics with synergism. Mini Rev. Med. Chem. 2010, 10, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Shull, S.; Heintz, N.H.; Periasamy, M.; Manohar, M.; Janssen, Y.M.; Marsh, J.P.; Mossman, B.T. Differential regulation of antioxidant enzymes in response to oxidants. J. Biol. Chem. 1991, 266, 24398–24403. [Google Scholar] [PubMed]
- Jefferies, H.; Coster, J.; Khalil, A.; Bot, J.; McCauley, R.D.; Hall, J.C. Glutathione. ANZ J. Surg. 2003, 73, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Brandes, R.P.; Weissmann, N.; Schroder, K. NADPH oxidases in cardiovascular disease. Free Radic. Biol. Med. 2010, 49, 687–706. [Google Scholar] [CrossRef] [PubMed]
- Yap, L.P.; Sancheti, H.; Ybanez, M.D.; Garcia, J.; Cadenas, E.; Han, D. Determination of GSH, GSSG, and GSNO using HPLC with electrochemical detection. Method Enzymol. 2010, 473, 137–147. [Google Scholar]
- Gu, J.; Ahn-Jarvis, J.H.; Riedl, K.M.; Schwartz, S.J.; Clinton, S.K.; Vodovotz, Y. Characterization of black raspberry functional food products for cancer prevention human clinical trials. J. Agric. Food Chem. 2014, 62, 3997–4006. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.; Riedl, K.M.; Schwartz, S.K.; Zhang, X.; Clinton, S.K.; Chen, T. Efficacy comparison of lyophilised black raspberries and combination of celecoxib and PBIT in prevention of carcinogen-induced oesophageal cancer in rats. J. Funct. Food 2016, 27, 11. [Google Scholar] [CrossRef]
- Kresty, L.A.; Mallery, S.R.; Stoner, G.D. Black raspberries in cancer clinical trials: Past, present and future. J. Berry Res. 2016, 6, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Kula, M.; Krauze-Baranowska, M. Rubus occidentalis: The black raspberry—Its potential in the prevention of cancer. Nutr. Cancer 2016, 68, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Norris, K.M.; Okie, W.; Yakaitis, C.L.; Pazdro, R. The anthocyanin cyanidin-3-O-β-glucoside modulates murine glutathione homeostasis in a manner dependent on genetic background. Redox Biol. 2016, 9, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Del Bo, C.; Martini, D.; Porrini, M.; Klimis-Zacas, D.; Riso, P. Berries and oxidative stress markers: An overview of human intervention studies. Food Funct. 2015, 6, 2890–2917. [Google Scholar] [CrossRef] [PubMed]
- Pratheeshkumar, P.; Son, Y.O.; Wang, X.; Divya, S.P.; Joseph, B.; Hitron, J.A.; Wang, L.; Kim, D.; Yin, Y.; Roy, R.V.; et al. Cyanidin-3-glucoside inhibits UVB-induced oxidative damage and inflammation by regulating map kinase and NF-kappab signaling pathways in SKH-1 hairless mice skin. Toxicol. Appl. Pharmacol. 2014, 280, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Jia, Q.; Wang, Y.; Zhang, Y.; Xia, M. The anthocyanin cyanidin-3-O-β-glucoside, a flavonoid, increases hepatic glutathione synthesis and protects hepatocytes against reactive oxygen species during hyperglycemia: Involvement of a cAMP-PKA-dependent signaling pathway. Free Radic. Biol. Med. 2012, 52, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Benhar, M.; Dalyot, I.; Engelberg, D.; Levitzki, A. Enhanced ROS production in oncogenically transformed cells potentiates c-jun N-terminal kinase and p38 mitogen-activated protein kinase activation and sensitization to genotoxic stress. Mol. Cell. Biol. 2001, 21, 6913–6926. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-κB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.A.; Smith, M.F., Jr.; Sanders, M.K.; Ernst, P.B. Reactive oxygen and nitrogen species differentially regulate toll-like receptor 4-mediated activation of NF-κB and interleukin-8 expression. Infect. Immun. 2004, 72, 2123–2130. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, D.; Resnick, M.B.; Behar, J.; Wands, J.; Cao, W. Signaling in H2O2-induced increase in cell proliferation in barrett's esophageal adenocarcinoma cells. J. Pharmacol. Exp. Ther. 2011, 339, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Li, J.; Qiao, J.; Jain, S.; Mark Evers, B.; Chung, D.H. PKD prevents H2O2-induced apoptosis via NF-κB and p38 MAPK in RIE-1 cells. Biochem. Biophys. Res. Commun. 2009, 378, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Karin, M. NF-κB in cancer development and progression. Nature 2006, 441, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Crusz, S.M.; Balkwill, F.R. Inflammation and cancer: Advances and new agents. Nat. Rev. Clin. Oncol. 2015, 12, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between NRF2 and NF-κB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, Y.; Okumura, H.; Uchikado, Y.; Kita, Y.; Sasaki, K.; Owaki, T.; Ishigami, S.; Natsugoe, S. NRF2 is useful for predicting the effect of chemoradiation therapy on esophageal squamous cell carcinoma. Ann. Surg. Oncol. 2014, 21, 2347–2352. [Google Scholar] [CrossRef] [PubMed]
- Barrett, C.W.; Ning, W.; Chen, X.; Smith, J.J.; Washington, M.K.; Hill, K.E.; Coburn, L.A.; Peek, R.M.; Chaturvedi, R.; Wilson, K.T.; et al. Tumor suppressor function of the plasma glutathione peroxidase GPX3 in colitis-associated carcinoma. Cancer Res. 2013, 73, 1245–1255. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | NMBA a | Diet | No. of Rats | Tumor Incidence (%) | Tumor Multiplicity Mean ± SE b | Tumor Volume (mm3) c Mean ± SE |
|---|---|---|---|---|---|---|
| 1 | − | AIN-76A | 20 | 0 | 0 | 0 |
| 2 | + | AIN-76A | 20 | 100 | 4.73 ± 0.45 | 7.91 ± 2.67 |
| 3 | + | 5% BRB | 20 | 81.5 d | 1.44 ± 0.26 e | 9.12 ± 3.76 |
| Normal | Preneoplasia | Papilloma | |||
|---|---|---|---|---|---|
| NMBA h | NMBA + BRB i | NMBA h | NMBA + BRB i | ||
| H2O2 a | 7.2 ± 0.9 | 44.1 ± 3.8 **** | 9.6 ± 1.2 **** | 23.7 ± 2.3 **** | 10.2 ± 1.1 *** |
| LPO b | 10.5 ± 2.5 | 74.0 ± 7.3 **** | 20.1 ± 5.7 **** | 97.1 ± 4.1 **** | 22.8 ± 4.8 **** |
| GPx (H2O2) c | 85.4 ± 3.5 | 19.4 ± 4.6 **** | 72.6 ± 8.0 **** | 29.8 ± 2.7 **** | 85.9 ± 10.4 **** |
| GPx (CuOOH) d | 213.4 ± 33.5 | 55.4 ± 17.8 **** | 205.9 ± 13.1 **** | 79.9 ± 16.6 **** | 231.5 ± 16.4 **** |
| SOD2 e | 0.39 ± 0.0 | 0.17 ± 0.0 *** | 0.31 ± 0.0 ** | 0.18 ± 0.0 *** | 0.33 ± 0.0 ** |
| GSSG/GSH f | 1.0 ± 0.0 | 7.3 ± 1.0 **** | 2.6 ± 0.8 **** | 7.7 ± 1.4 **** | 2.8 ± 0.6 **** |
| NADP+/NADPH g | 1.0 ± 0.0 | 8.7 ± 0.8 **** | 2.0 ± 0.3 **** | 11.3 ± 1.1 **** | 2.8 ± 0.3 **** |
| NMBA | BRB | |
|---|---|---|
| Tissue oxidative stress index | ||
| 8-OxoG IHC staining | ↑ | ↓ |
| Anti-oxidative enzymes | ||
| GPx | ↓ | ↑ |
| SOD2 | ↓ | ↑ |
| Cell oxidative stress index | ||
| H2O2 and LPO | ↑ | ↓ |
| GSSG/GSH | ↑ | ↓ |
| NADP+/NADPH | ↑ | ↓ |
| NFκB pathway | ||
| p65, IκBα, IKKα/β | ↑ | ↓ |
| MAPK pathway | ||
| p38, ERK and SAPK/JNK | ↑ | ↓ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, N.; Chen, F.; Zhang, X.; Clinton, S.K.; Tang, X.; Sun, Z.; Chen, T. Suppression of Oxidative Stress and NFκB/MAPK Signaling by Lyophilized Black Raspberries for Esophageal Cancer Prevention in Rats. Nutrients 2017, 9, 413. https://doi.org/10.3390/nu9040413
Shi N, Chen F, Zhang X, Clinton SK, Tang X, Sun Z, Chen T. Suppression of Oxidative Stress and NFκB/MAPK Signaling by Lyophilized Black Raspberries for Esophageal Cancer Prevention in Rats. Nutrients. 2017; 9(4):413. https://doi.org/10.3390/nu9040413
Chicago/Turabian StyleShi, Ni, Fang Chen, Xiaoli Zhang, Steven K. Clinton, Xiaofei Tang, Zheng Sun, and Tong Chen. 2017. "Suppression of Oxidative Stress and NFκB/MAPK Signaling by Lyophilized Black Raspberries for Esophageal Cancer Prevention in Rats" Nutrients 9, no. 4: 413. https://doi.org/10.3390/nu9040413
APA StyleShi, N., Chen, F., Zhang, X., Clinton, S. K., Tang, X., Sun, Z., & Chen, T. (2017). Suppression of Oxidative Stress and NFκB/MAPK Signaling by Lyophilized Black Raspberries for Esophageal Cancer Prevention in Rats. Nutrients, 9(4), 413. https://doi.org/10.3390/nu9040413