Body Weight Cycling with Identical Diet Composition Does Not Affect Energy Balance and Has No Adverse Effect on Metabolic Health Parameters


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Induction of Body Weight (BW) Cycling
2.3. Parameters of Interest
2.4. Indirect Calorimetry
2.5. End Point Parameters
2.6. Serum Measurements
2.7. Histology of Epididymal Adipose Tissue
2.8. Data Analysis and Statistics
3. Results
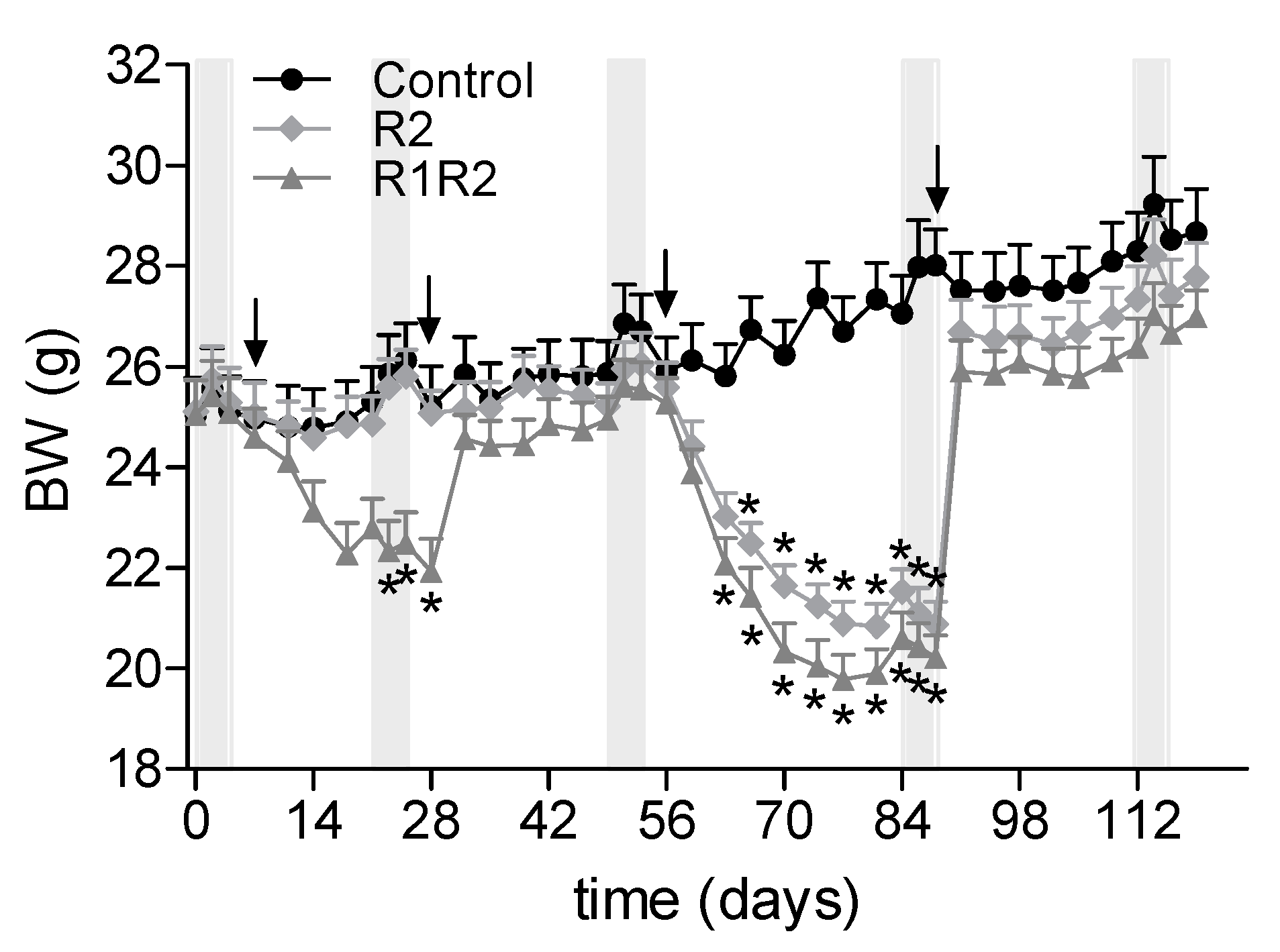
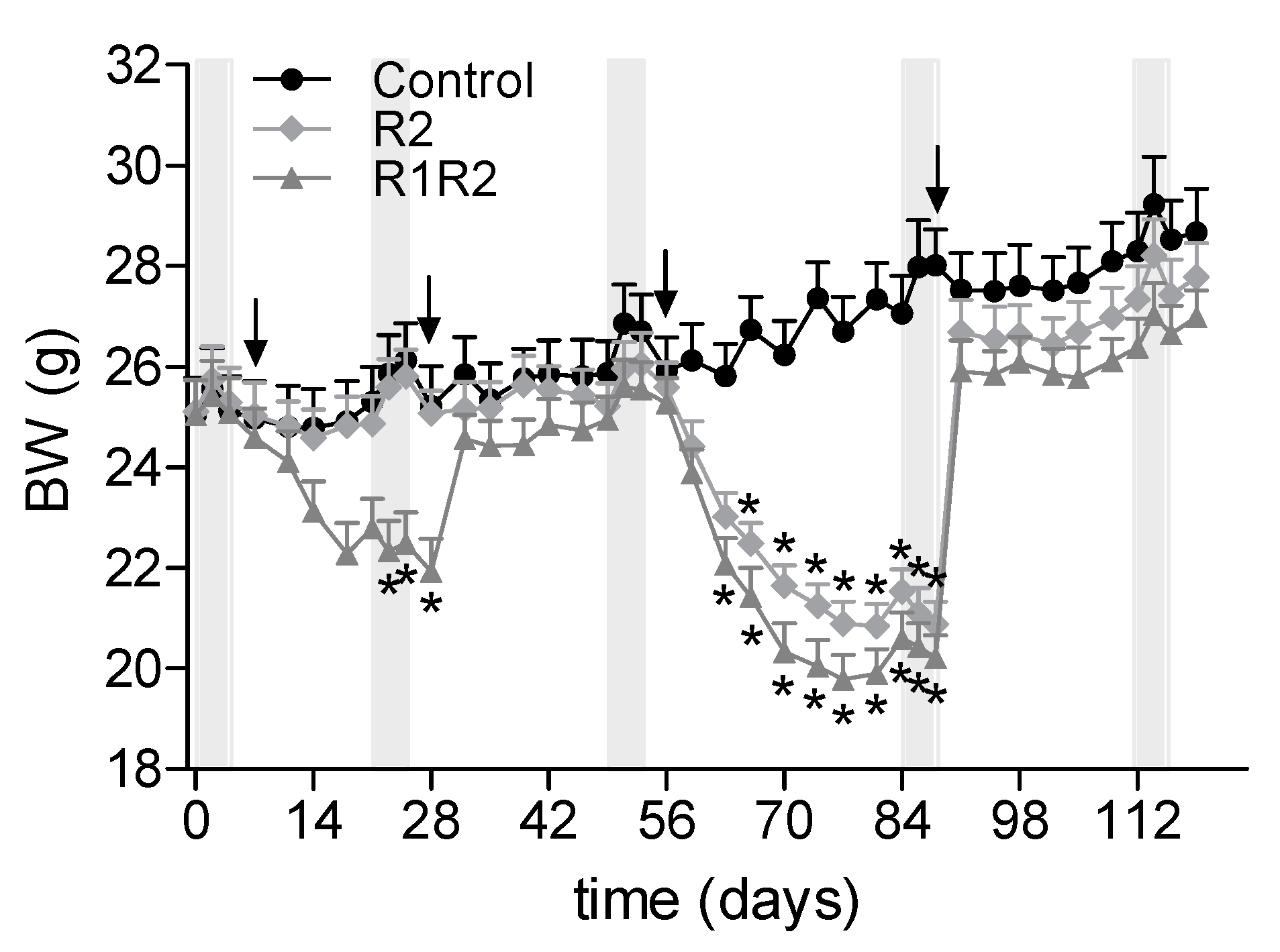
3.1. BW Cycling
3.2. Effects of a History of BW Cycling
3.2.1. Additional BW Cycle: R1R2 vs. R2
3.2.2. BW, Food Intake, and Food Efficiency at the Endpoint
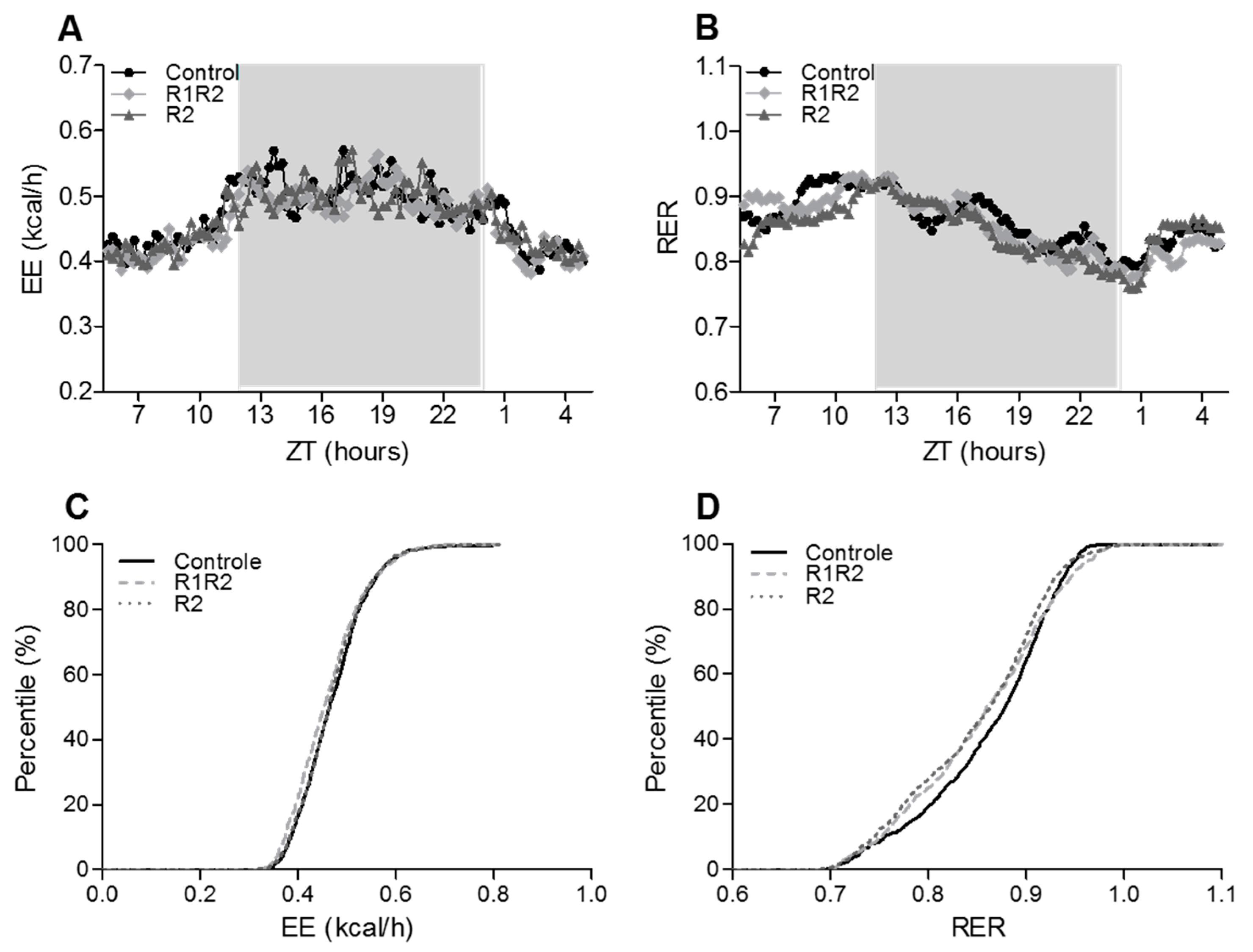
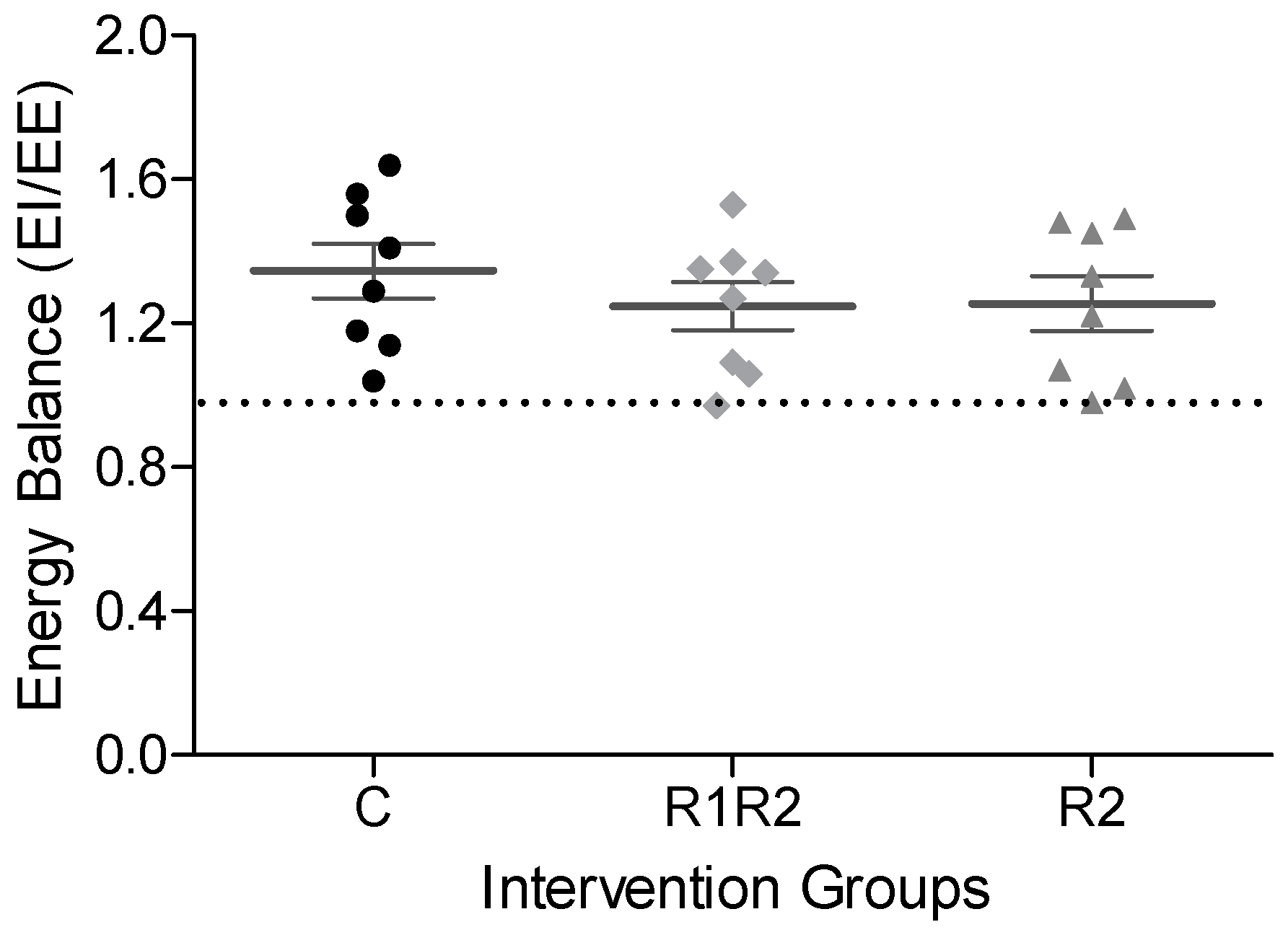
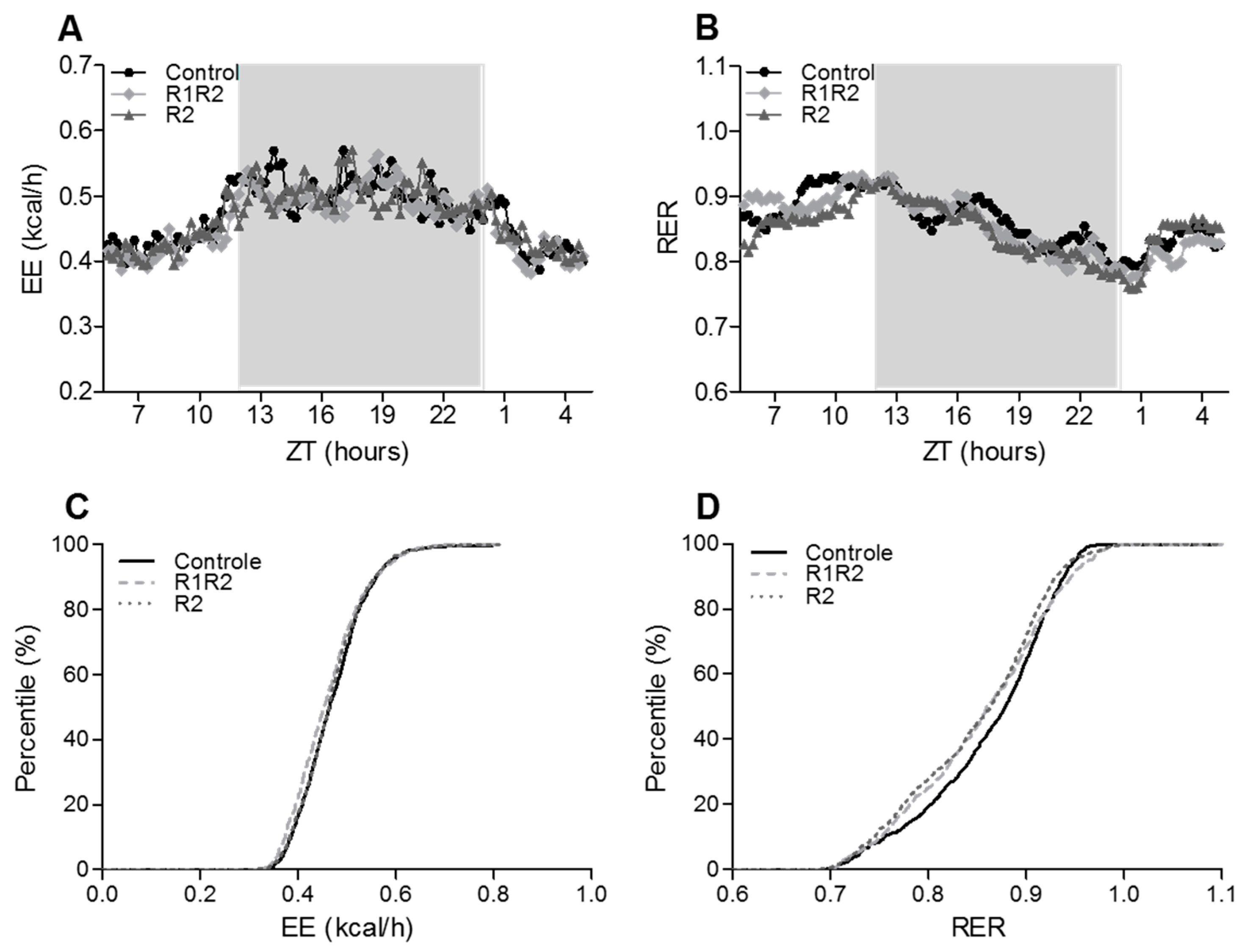
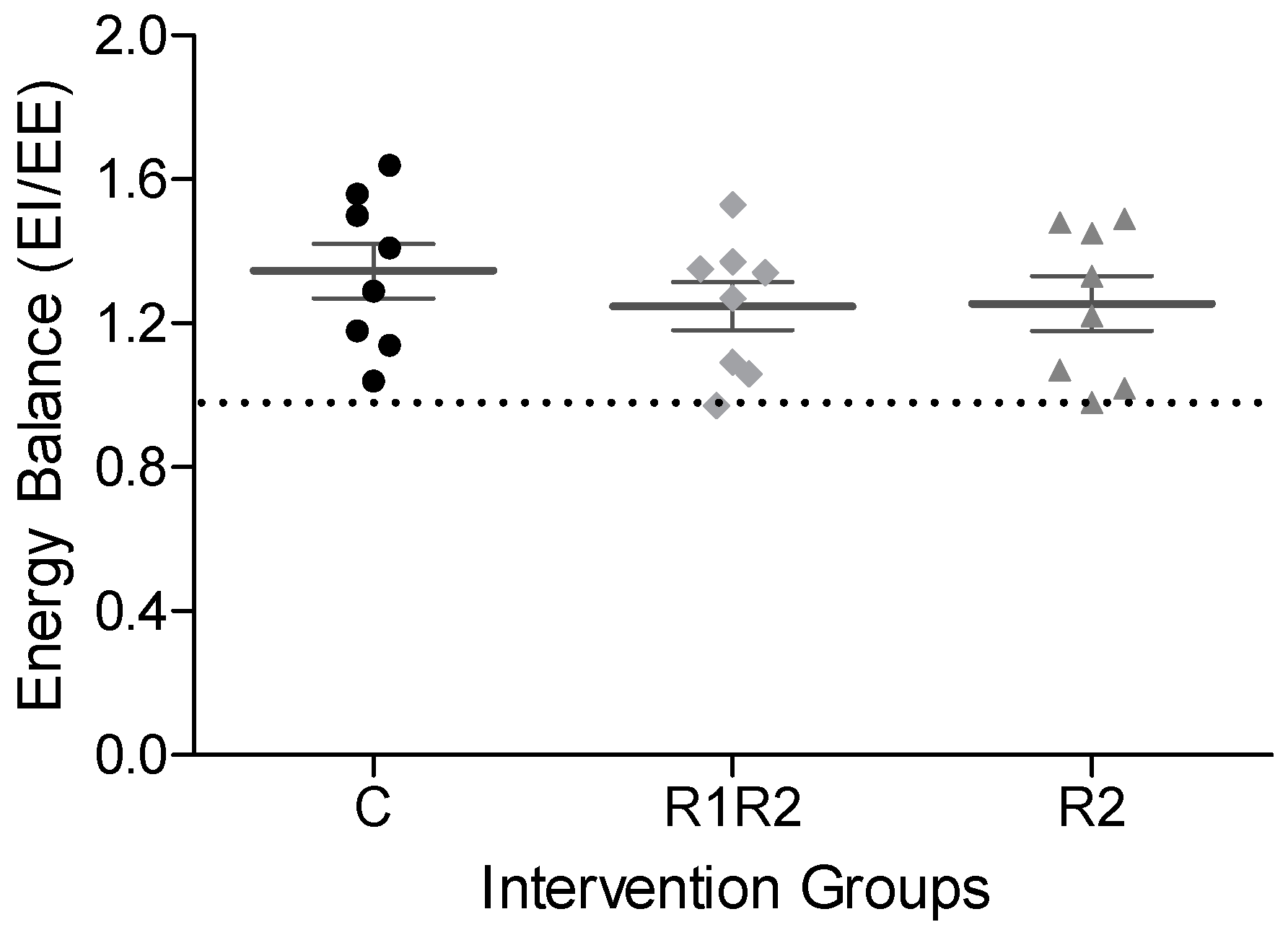
3.2.3. EE, RER, and Energy Balance at the Endpoint
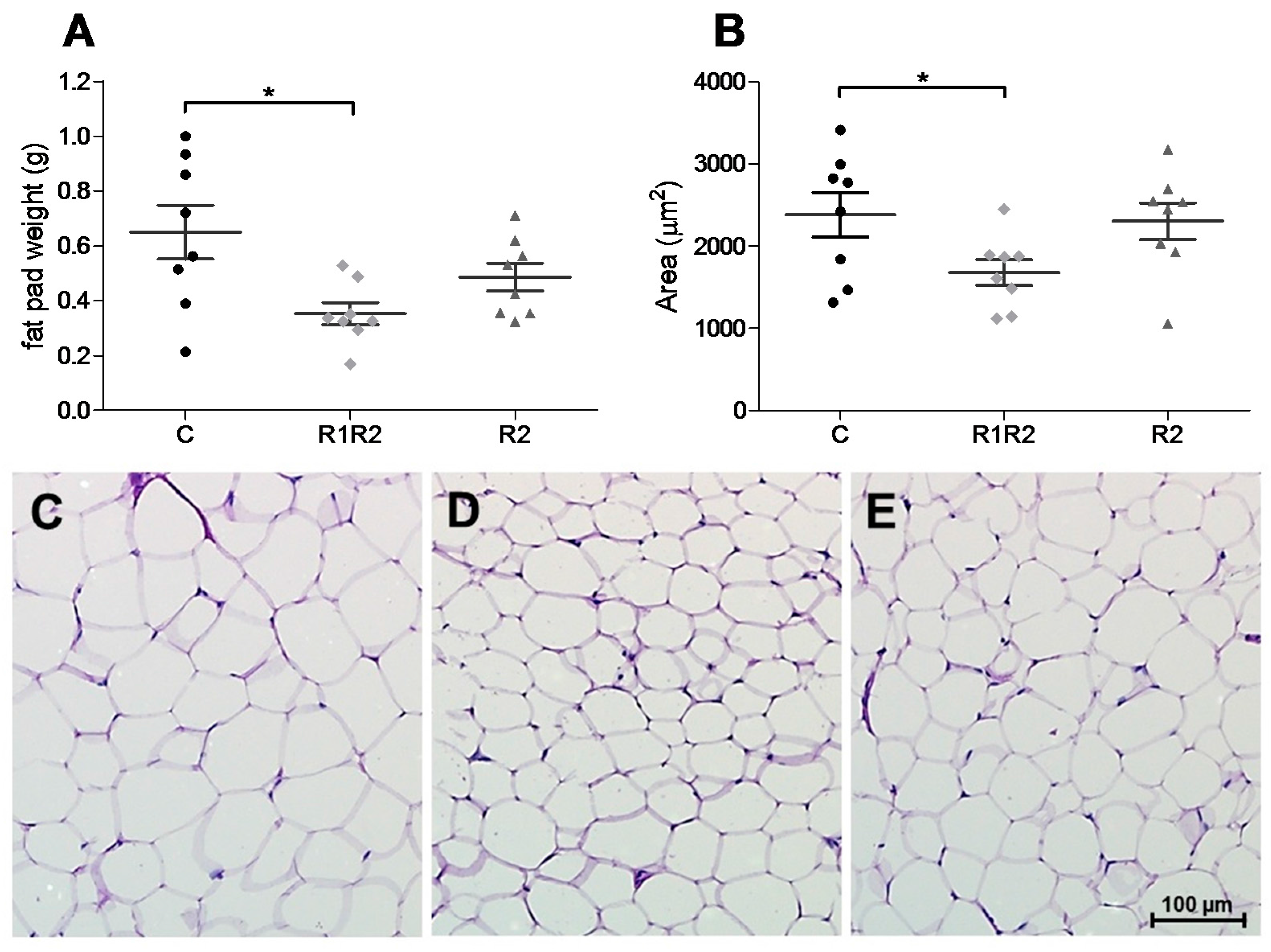
3.2.4. Adipose Tissue Weight and Histology
3.2.5. Serum Concentrations of Lipids and Peptide Hormones
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wing, R.R.; Phelan, S. Long-term weight loss maintenance. Am. J. Clin. Nutr. 2005, 82, 222S–225S. [Google Scholar] [PubMed]
- Strychar, I.; Lavoie, M.E.; Messier, L.; Karelis, A.D.; Doucet, E.; Prud’homme, D.; Fontaine, J.; Rabasa-Lhoret, R. Anthropometric, metabolic, psychosocial, and dietary characteristics of overweight/obese postmenopausal women with a history of weight cycling: A monet (montreal ottawa new emerging team) study. J. Am. Diet. Assoc. 2009, 109, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Weight Cycling. National task force on the prevention and treatment of obesity. JAMA 1994, 272, 1196–1202.
- Reed, G.W.; Hill, J.O. Weight cycling: A review of the animal literature. Obes. Res. 1993, 1, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Brownell, K.D.; Greenwood, M.R.; Stellar, E.; Shrager, E.E. The effects of repeated cycles of weight loss and regain in rats. Physiol. Behav. 1986, 38, 459–464. [Google Scholar] [CrossRef]
- Miller, G.D.; Dimond, A.G.; Stern, J.S. The effect of repeated episodes of dietary restriction and refeeding on systolic blood pressure and food intake in exercise-trained normotensive rats. Obes. Res. 2000, 8, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Ernsberger, P.; Koletsky, R.J.; Baskin, J.S.; Collins, L.A. Consequences of weight cycling in obese spontaneously hypertensive rats. Am. J. Physiol. 1996, 270, R864–R872. [Google Scholar] [PubMed]
- Rozen, R.; Brigant, L.; Apfelbaum, M. Effects of cycles of food restriction followed by ad libitum refeeding on body composition and energy expenditure in obese rats. Am. J. Clin. Nutr. 1994, 59, 560–565. [Google Scholar] [PubMed]
- Duarte, F.O.; Sene-Fiorese, M.; Manzoni, M.S.; de Freitas, L.F.; Cheik, N.C.; Garcia de Oliveira Duarte, A.C.; Nonaka, K.O.; Damaso, A. Caloric restriction and refeeding promoted different metabolic effects in fat depots and impaired dyslipidemic profile in rats. Nutrition 2008, 24, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Lauer, J.B.; Reed, G.W.; Hill, J.O. Effects of weight cycling induced by diet cycling in rats differing in susceptibility to dietary obesity. Obes. Res. 1999, 7, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Buison, A.; Uhley, V.; Jen, K.L. Long-term weight cycling in female wistar rats: Effects on metabolism. Obes. Res. 1995, 3, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Uhley, V.E.; Pellizzon, M.A.; Buison, A.M.; Guo, F.; Djuric, Z.; Jen, K.L. Chronic weight cycling increases oxidative DNA damage levels in mammary gland of female rats fed a high-fat diet. Nutr. Cancer 1997, 29, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Hoevenaars, F.P.M.; Keijer, J.; Swarts, H.J.; Snaas-Alders, S.; Bekkenkamp-Grovenstein, M.; van Schothorst, E.M. Effects of dietary history on energy metabolism and physiological parameters in c57bl/6j mice. Exp. Physiol. 2013, 98, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Schofield, S.E.; Parkinson, J.R.; Henley, A.B.; Sahuri-Arisoylu, M.; Sanchez-Canon, G.J.; Bell, J.D. Metabolic dysfunction following weight cycling in male mice. Int. J. Obes. 2017, 41, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Kochan, Z.; Karbowska, J.; Swierczynski, J. The effects of weight cycling on serum leptin levels and lipogenic enzyme activities in adipose tissue. J. Physiol. Pharmacol. 2006, 57, 115–127. [Google Scholar] [PubMed]
- Desautels, M.; Dulos, R.A. Effects of repeated cycles of fasting-refeeding on brown adipose tissue composition in mice. Am. J. Physiol. 1988, 255, E120–E128. [Google Scholar] [PubMed]
- Bogden, J.D.; Kemp, F.W.; Huang, A.E.; Shapses, S.A.; Ambia-Sobhan, H.; Jagpal, S.; Brown, I.L.; Birkett, A.M. Bone mineral density and content during weight cycling in female rats: Effects of dietary amylase-resistant starch. Nutr. Metab. 2008, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Duivenvoorde, L.P.; van Schothorst, E.M.; Bunschoten, A.; Keijer, J. Dietary restriction of mice on a high-fat diet induces substrate efficiency and improves metabolic health. J. Mol. Endocrinol. 2011, 47, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Riachi, M.; Himms-Hagen, J.; Harper, M.E. Percent relative cumulative frequency analysis in indirect calorimetry: Application to studies of transgenic mice. Can. J. Physiol. Pharmacol. 2004, 82, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Reed, G.W.; Cox, G.; Yakubu, F.; Ding, L.; Hill, J.O. Effects of weight cycling in rats allowed a choice of diet. Am. J. Physiol. 1993, 264, R35–R40. [Google Scholar] [PubMed]
- Sea, M.M.; Fong, W.P.; Huang, Y.; Chen, Z.Y. Weight cycling-induced alteration in fatty acid metabolism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R1145–R1155. [Google Scholar] [PubMed]
- Randall Simpson, J.A.; Wainwright, P.E.; Hoffman-Goetz, L.; Levesque, S. Effects of different weight loss treatments on weight cycling and metabolic measures in male mice. Physiol. Behav. 1994, 56, 197–201. [Google Scholar] [CrossRef]
- Pellizzon, M.A.; Buison, A.M.; Jen, K.L. Short-term weight cycling in aging female rats increases rate of weight gain but not body fat content. Int. J. Obes. Relat. Metab. Disord. 2000, 24, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, J.M.; Yasari, S.; Abdennadher, M.; Paquette, A. Effects of alternations (10 days) of high-fat with normal diet on liver lipid infiltration, fat gain, and plasma metabolic profile in rats. Physiol. Behav. 2005, 86, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Graham, B.; Chang, S.; Lin, D.; Yakubu, F.; Hill, J.O. Effect of weight cycling on susceptibility to dietary obesity. Am. J. Physiol. 1990, 259, R1096–R1102. [Google Scholar] [PubMed]
- Gerardo-Gettens, T.; Miller, G.D.; Horwitz, B.A.; McDonald, R.B.; Brownell, K.D.; Greenwood, M.R.; Rodin, J.; Stern, J.S. Exercise decreases fat selection in female rats during weight cycling. Am. J. Physiol. 1991, 260, R518–R524. [Google Scholar] [PubMed]
- Ernsberger, P.; Koletsky, R.J.; Kilani, A.; Viswan, G.; Bedol, D. Effects of weight cycling on urinary catecholamines: Sympathoadrenal role in refeeding hypertension. J. Hypertens. 1998, 16, 2001–2005. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.; Halaas, J.; Ravussin, E.; Pratley, R.E.; Lee, G.H.; Zhang, Y.; Fei, H.; Kim, S.; Lallone, R.; Ranganathan, S.; et al. Leptin levels in human and rodent: Measurement of plasma leptin and ob rna in obese and weight-reduced subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Arner, P.; Spalding, K.L. Fat cell turnover in humans. Biochem. Biophys. Res. Commun. 2010, 396, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T. Adiponectin and adiponectin receptors. Endocr. Rev. 2005, 26, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Barnea, M.; Madar, Z.; Froy, O. High-fat diet delays and fasting advances the circadian expression of adiponectin signaling components in mouse liver. Endocrinology 2009, 150, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Madsen, E.L.; Rissanen, A.; Bruun, J.M.; Skogstrand, K.; Tonstad, S.; Hougaard, D.M.; Richelsen, B. Weight loss larger than 10% is needed for general improvement of levels of circulating adiponectin and markers of inflammation in obese subjects: A 3-year weight loss study. Eur. J. Endocrinol. 2008, 158, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Durand, J.L.; Hosinking, W.; Jelicks, L.A. Time course of effects of inhalation anesthesia on blood glucose level in male and female c57bl/6 mice. Horm. Metab. Res. 2009, 41, 339–341. [Google Scholar] [CrossRef] [PubMed]
- Yokomichi, H.; Ohde, S.; Takahashi, O.; Mochizuki, M.; Takahashi, A.; Yoda, Y.; Tsuji, M.; Akiyama, Y.; Yamagata, Z. Weight cycling and the subsequent onset of type 2 diabetes mellitus: 10-year cohort studies in urban and rural japan. BMJ Open 2017, 7, e014684. [Google Scholar] [CrossRef] [PubMed]
- Field, A.E.; Malspeis, S.; Willett, W.C. Weight cycling and mortality among middle-aged or older women. Arch. Intern. Med. 2009, 169, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Field, A.E.; Manson, J.E.; Laird, N.; Williamson, D.F.; Willett, W.C.; Colditz, G.A. Weight cycling and the risk of developing type 2 diabetes among adult women in the united states. Obes. Res. 2004, 12, 267–274. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control (n = 8) | R1R2 (n = 8) | R2 (n = 8) |
|---|---|---|---|
| BW change t = 0–119 (g) | 3.63 ± 0.36 a | 1.95 ± 0.37 b | 2.68 ± 0.34 a,b |
| FI cumulative (t = 0–119) (g) | 379.9 ± 4.4 a | 347.0 ± 4.4 b | 361.7± 7.2 a,b |
| BW at t = 119 (g) | 28.68 ± 0.86 a | 26.99 ± 0.52 a | 27.79 ± 0.67 a |
| FI (t = 116–119) (g/day) | 2.87 ± 0.13 a | 2.90 ± 0.07 a | 2.95 ± 0.12 a |
| FE (t = 116–119; BW gain/FI) | 0.014 ± 0.023 a | 0.037 ± 0.026 a | 0.037 ± 0.023 a |
| EE (kcal/24 h) | 11.33 ± 0.08 a | 11.09 ± 0.16 a | 11.17 ± 0.26 a |
| EI (kcal/24 h) | 12.63 ± 1.40 a | 12.25 ± 0.57 a | 12.92 ± 0.81 a |
| Parameter | Control | R1R2 | R2 |
|---|---|---|---|
| Glucose (mmol/L) | 12.11 ± 0.72 a | 11.88 ± 0.60 a | 11.48 ± 0.58 a |
| Leptin (ng/mL) | 2.73 ± 0.64 a | 1.04 ± 0.36 b | 1.09 ± 0.15 b |
| Adiponectin (µg/mL) | 28.15 ± 2.52 a | 29.62 ± 2.77 a | 35.47 ± 1.03 a,# |
| FFA (µmol/L) | 793.9 ± 66.1 a | 667.5 ± 89.6 a | 727.6 ± 67.8 a |
| TG (mg/dL) | 47.45 ± 2.46 a | 43.32 ± 2.27 a | 39.39 ± 1.76 a,# |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palm, I.F.; Schram, R.G.A.E.; Swarts, H.J.M.; Van Schothorst, E.M.; Keijer, J. Body Weight Cycling with Identical Diet Composition Does Not Affect Energy Balance and Has No Adverse Effect on Metabolic Health Parameters. Nutrients 2017, 9, 1149. https://doi.org/10.3390/nu9101149
Palm IF, Schram RGAE, Swarts HJM, Van Schothorst EM, Keijer J. Body Weight Cycling with Identical Diet Composition Does Not Affect Energy Balance and Has No Adverse Effect on Metabolic Health Parameters. Nutrients. 2017; 9(10):1149. https://doi.org/10.3390/nu9101149
Chicago/Turabian StylePalm, Inge F., Rianne G. A. E. Schram, Hans J. M. Swarts, Evert M. Van Schothorst, and Jaap Keijer. 2017. "Body Weight Cycling with Identical Diet Composition Does Not Affect Energy Balance and Has No Adverse Effect on Metabolic Health Parameters" Nutrients 9, no. 10: 1149. https://doi.org/10.3390/nu9101149
APA StylePalm, I. F., Schram, R. G. A. E., Swarts, H. J. M., Van Schothorst, E. M., & Keijer, J. (2017). Body Weight Cycling with Identical Diet Composition Does Not Affect Energy Balance and Has No Adverse Effect on Metabolic Health Parameters. Nutrients, 9(10), 1149. https://doi.org/10.3390/nu9101149