Milk Intake at Midlife and Cognitive Decline over 20 Years. The Atherosclerosis Risk in Communities (ARIC) Study

 ,
,
Abstract
1. Introduction
2. Materials and Methods
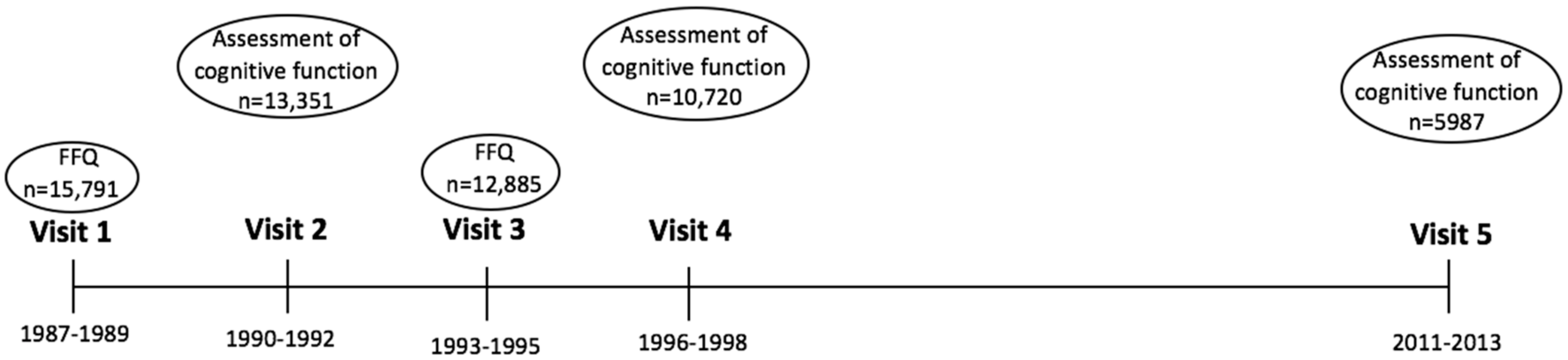
2.1. Study Population
2.2. Assessment of Cognitive Function
2.3. Assessment of Milk Intake
2.4. Diet Quality Score
2.5. Covariates
2.6. Lactase Persistence Genotype
2.7. Statistical Analysis
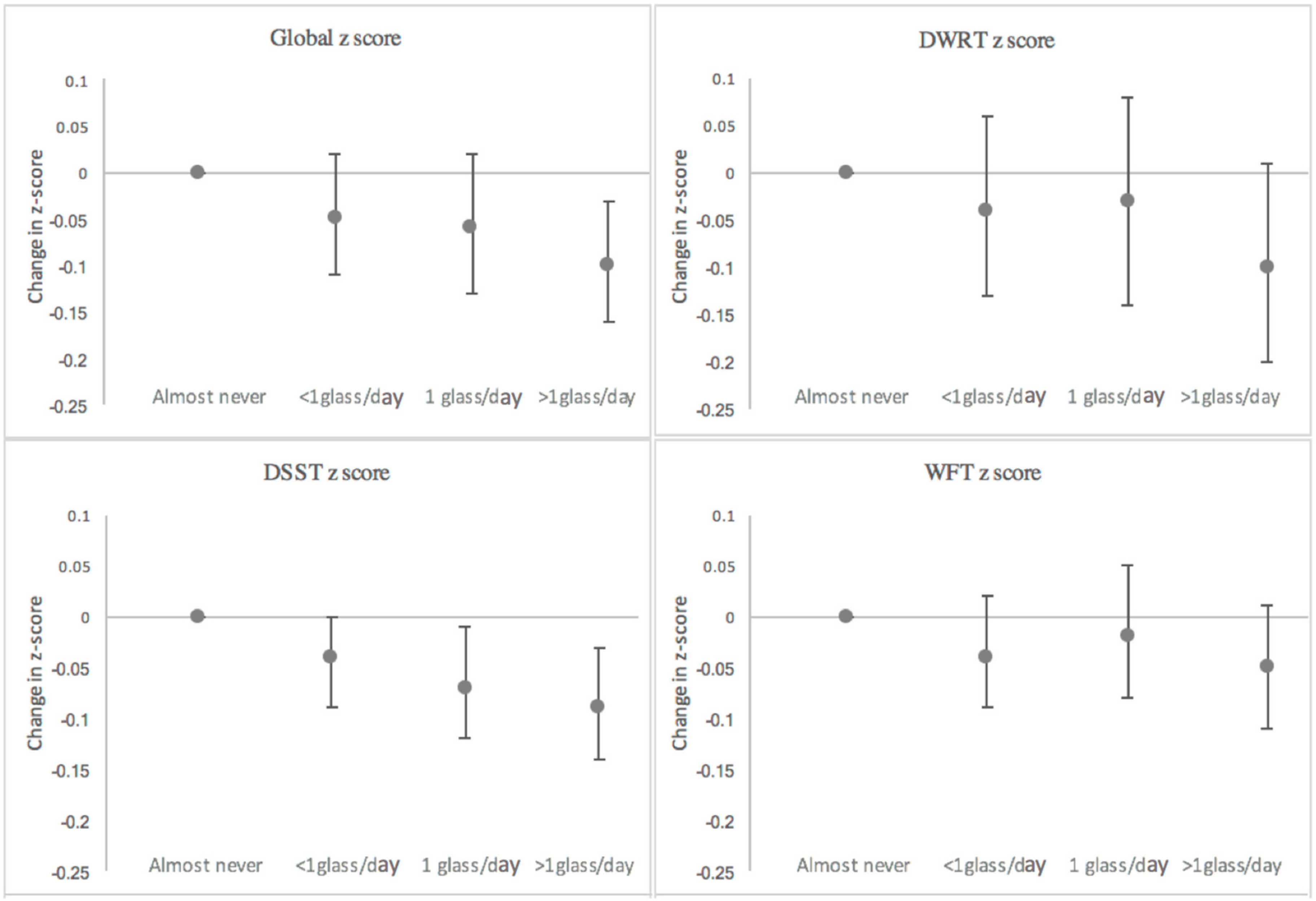
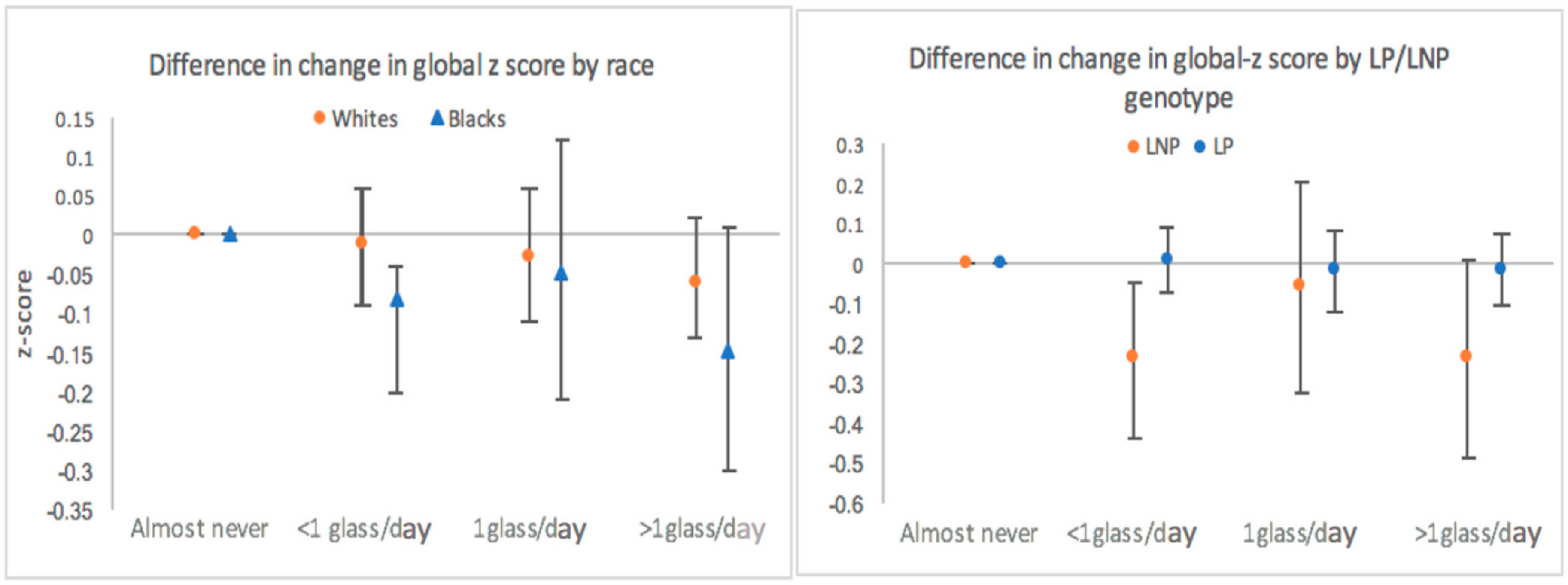
3. Results
3.1. Total Milk Intake
3.2. Lactase Persistence
3.3. Skim/Low-Fat Milk and Total Dairy
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Beydoun, M.A.; Beydoun, H.A.; Gamaldo, A.A.; Teel, A.; Zonderman, A.B.; Wang, Y. Epidemiologic studies of modifiable factors associated with cognition and dementia: Systematic review and meta-analysis. BMC Public Health 2014, 14, 643. [Google Scholar] [CrossRef] [PubMed]
- Schaie, K.W. The Seattle Longitudinal Study: A thirty-five-year inquiry of adult intellectual development. Z. Gerontol. 1993, 26, 129–137. [Google Scholar] [PubMed]
- Sachs-Ericsson, N.; Blazer, D.G. Racial differences in cognitive decline in a sample of community-dwelling older adults: The mediating role of education and literacy. Am. J. Geriatr. Psychiatry 2005, 13, 968–975. [Google Scholar] [CrossRef] [PubMed]
- Comijs, H.C.; Dik, M.G.; Deeg, D.J.; Jonker, C. The course of cognitive decline in older persons: Results from the longitudinal aging study amsterdam. Dement. Geriatr. Cogn. Disord. 2004, 17, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Zelinski, E.M.; Stewart, S.T. Individual differences in 16-year memory changes. Psychol. Aging 1998, 13, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.S.; Boyle, P.A.; Segawa, E.; Yu, L.; Begeny, C.T.; Anagnos, S.E.; Bennett, D.A. The influence of cognitive decline on well-being in old age. Psychol. Aging 2013, 28, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Allerhand, M.; Gale, C.R.; Deary, I.J. The dynamic relationship between cognitive function and positive well-being in older people: A prospective study using the English Longitudinal Study of Aging. Psychol. Aging 2014, 29, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Salthouse, T.A. When does age-related cognitive decline begin? Neurobiol. Aging 2009, 30, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Mecocci, P.; MacGarvey, U.; Kaufman, A.E.; Koontz, D.; Shoffner, J.M.; Wallace, D.C.; Beal, M.F. Oxidative damage to mitochondrial DNA shows marked age-dependent increases in human brain. Ann. Neurol. 1993, 34, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Magnus, T. Ageing and neuronal vulnerability. Nat. Rev. Neurosci. 2006, 7, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Schrag, M.; Mueller, C.; Zabel, M.; Crofton, A.; Kirsch, W.M.; Ghribi, O.; Squitti, R.; Perry, G. Oxidative stress in blood in Alzheimer’s disease and mild cognitive impairment: A meta-analysis. Neurobiol. Dis. 2013, 59, 100–110. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Health and Human Services; U.S. Department of Agriculture. Dietary Guidelines for Americans; U.S. Department of Health and Human Services and U.S. Department of Agriculture: Washington, DC, USA, 2010.
- He, M.; Zhao, L.; Wei, M.J.; Yao, W.F.; Zhao, H.S.; Chen, F.J. Neuroprotective effects of (−)-epigallocatechin-3-gallate on aging mice induced by d-galactose. Biol. Pharm. Bull. 2009, 32, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Almeida, O.P.; Norman, P.; Hankey, G.; Jamrozik, K.; Flicker, L. Successful mental health aging: Results from a longitudinal study of older Australian men. Am. J. Geriatr. Psychiatry 2006, 14, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Salminen, L.E.; Paul, R.H. Oxidative stress and genetic markers of suboptimal antioxidant defense in the aging brain: A theoretical review. Rev. Neurosci. 2014, 25, 805–819. [Google Scholar] [CrossRef] [PubMed]
- Serra, J.A.; Dominguez, R.O.; Marschoff, E.R.; Guareschi, E.M.; Famulari, A.L.; Boveris, A. Systemic oxidative stress associated with the neurological diseases of aging. Neurochem. Res. 2009, 34, 2122–2132. [Google Scholar] [CrossRef] [PubMed]
- Barone, E.; Di Domenico, F.; Cenini, G.; Sultana, R.; Cini, C.; Preziosi, P.; Perluigi, M.; Mancuso, C.; Butterfield, D.A. Biliverdin reductase—A protein levels and activity in the brains of subjects with Alzheimer disease and mild cognitive impairment. Biochim. Biophys. Acta 2011, 1812, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Barone, E.; Di Domenico, F.; Cenini, G.; Sultana, R.; Coccia, R.; Preziosi, P.; Perluigi, M.; Mancuso, C.; Butterfield, D.A. Oxidative and nitrosative modifications of biliverdin reductase-A in the brain of subjects with Alzheimer’s disease and amnestic mild cognitive impairment. J. Alzheimers Dis. 2011, 25, 623–633. [Google Scholar] [PubMed]
- Bonda, D.J.; Wang, X.; Lee, H.G.; Smith, M.A.; Perry, G.; Zhu, X. Neuronal failure in Alzheimer’s disease: A view through the oxidative stress looking-glass. Neurosci. Bull. 2014, 30, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Escudero, V.; Martin-Maestro, P.; Perry, G.; Avila, J. Deconstructing mitochondrial dysfunction in Alzheimer disease. Oxid. Med. Cell. Longev. 2013, 2013, 162152. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.H.; Wang, X.; Zhu, X. Mitochondrial defects and oxidative stress in Alzheimer disease and Parkinson disease. Free Radic. Biol. Med. 2013, 62, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Baierle, M.; Nascimento, S.N.; Moro, A.M.; Brucker, N.; Freitas, F.; Gauer, B.; Durgante, J.; Bordignon, S.; Zibetti, M.; Trentini, C.M.; et al. Relationship between inflammation and oxidative stress and cognitive decline in the institutionalized elderly. Oxid. Med. Cell. Longev. 2015, 2015, 804198. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Prakash, A.; Dogra, S. Centella asiatica Attenuates d-Galactose-Induced Cognitive Impairment, Oxidative and Mitochondrial Dysfunction in Mice. Int. J. Alzheimers Dis. 2011, 2011, 347569. [Google Scholar] [PubMed]
- Wang, W.; Li, S.; Dong, H.P.; Lv, S.; Tang, Y.Y. Differential impairment of spatial and nonspatial cognition in a mouse model of brain aging. Life Sci. 2009, 85, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.Z.; Ge, Q.H.; Qu, R.; Li, Q.; Ma, S.P. Paeonol attenuates neurotoxicity and ameliorates cognitive impairment induced by d-galactose in ICR mice. J. Neurol. Sci. 2009, 277, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Zuo, P.; Zhang, Q.; Li, X.; Hu, Y.; Long, J.; Packer, L.; Liu, J. Chronic systemic d-galactose exposure induces memory loss, neurodegeneration, and oxidative damage in mice: Protective effects of R-alpha-lipoic acid. J. Neurosci. Res. 2006, 84, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Wang, L.; Zuo, P.; Han, Z.; Fang, Z.; Li, W.; Liu, J. d-galactose-caused life shortening in Drosophila melanogaster and Musca domestica is associated with oxidative stress. Biogerontology 2004, 5, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Liaquat, L.; Shahzad, S.; Sadir, S.; Madiha, S.; Batool, Z.; Tabassum, S.; Saleem, S.; Naqvi, F.; Perveen, T. A high dose of short term exogenous d-galactose administration in young male rats produces symptoms simulating the natural aging process. Life Sci. 2015, 124, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.M.; Lu, J.; Zheng, Y.L.; Zhou, Z.; Shan, Q.; Ma, D.F. Purple sweet potato color repairs d-galactose-induced spatial learning and memory impairment by regulating the expression of synaptic proteins. Neurobiol. Learn. Mem. 2008, 90, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Bao, M.; Li, D.; Li, Y.M. Advanced glycation in d-galactose induced mouse aging model. Mech. Ageing Dev. 1999, 108, 239–251. [Google Scholar] [CrossRef]
- Vlassara, H. The AGE-receptor in the pathogenesis of diabetic complications. Diabetes Metab. Res. Rev. 2001, 17, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.M.; Yan, S.D.; Wautier, J.L.; Stern, D. Activation of receptor for advanced glycation end products: A mechanism for chronic vascular dysfunction in diabetic vasculopathy and atherosclerosis. Circ. Res. 1999, 84, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Bierhaus, A.; Humpert, P.M.; Morcos, M.; Wendt, T.; Chavakis, T.; Arnold, B.; Stern, D.M.; Nawroth, P.P. Understanding RAGE, the receptor for advanced glycation end products. J. Mol. Med. 2005, 83, 876–886. [Google Scholar] [CrossRef] [PubMed]
- Michaelsen, K.F.; Nielsen, A.L.; Roos, N.; Friis, H.; Molgaard, C. Cow’s milk in treatment of moderate and severe undernutrition in low-income countries. In Milk and Milk Products in Human Nutrition; Karger AG: Basel, Switzerland, 2011; Volume 67, pp. 99–111. [Google Scholar]
- Campbell, J.R.; Marshall, R.T. The Science of Providing Milk for Man; McGraw-Hill Book Co.: New York, NY, USA, 1975. [Google Scholar]
- Muehlhoff, E.; Bennett, A.; McMahon, D. Milk and Dairy Products in Human Nutrition; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013. [Google Scholar]
- Szilagyi, A. Adaptation to Lactose in Lactase Non Persistent People: Effects on Intolerance and the Relationship between Dairy Food Consumption and Evalution of Diseases. Nutrients 2015, 7, 6751–6779. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.K.; Waud, J.P.; Matthews, S.B. The molecular basis of lactose intolerance. Sci. Prog. 2009, 92, 241–287. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.; Kang, S.A.; Lee, H.O.; Lee, B.H.; Park, J.S.; Kim, J.H.; Jung, I.K.; Park, Y.J.; Lee, J.E. Relationships between dietary intake and cognitive function level in Korean elderly people. Public Health 2001, 115, 133–138. [Google Scholar] [CrossRef]
- Avila-Funes, J.A.; Garant, M.P.; Aguilar-Navarro, S. Relationship between determining factors for depressive symptoms and for dietary habits in older adults in Mexico. Rev. Panam. Salud Publica 2006, 19, 321–330. [Google Scholar] [PubMed]
- Rahman, A.; Sawyer Baker, P.; Allman, R.M.; Zamrini, E. Dietary factors and cognitive impairment in community-dwelling elderly. J. Nutr. Health Aging 2007, 11, 49–54. [Google Scholar] [PubMed]
- Crichton, G.E.; Elias, M.F.; Dore, G.A.; Robbins, M.A. Relation between dairy food intake and cognitive function: The Maine-Syracuse Longitudinal Study. Int. Dairy J. 2012, 22, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Vercambre, M.N.; Boutron-Ruault, M.C.; Ritchie, K.; Clavel-Chapelon, F.; Berr, C. Long-term association of food and nutrient intakes with cognitive and functional decline: A 13-year follow-up study of elderly French women. Br. J. Nutr. 2009, 102, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Laitinen, M.H.; Ngandu, T.; Rovio, S.; Helkala, E.L.; Uusitalo, U.; Viitanen, M.; Nissinen, A.; Tuomilehto, J.; Soininen, H.; Kivipelto, M. Fat intake at midlife and risk of dementia and Alzheimer’s disease: A population-based study. Dement. Geriatr. Cogn. Disord. 2006, 22, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Eskelinen, M.H.; Ngandu, T.; Helkala, E.L.; Tuomilehto, J.; Nissinen, A.; Soininen, H.; Kivipelto, M. Fat intake at midlife and cognitive impairment later in life: A population-based CAIDE study. Int. J. Geriatr. Psychiatry 2008, 23, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Kasagi, F.; Sasaki, H.; Masunari, N.; Mimori, Y.; Suzuki, G. Association between dementia and midlife risk factors: The Radiation Effects Research Foundation Adult Health Study. J. Am. Geriatr. Soc. 2003, 51, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Kesse-Guyot, E.; Assmann, K.E.; Andreeva, V.A.; Ferry, M.; Hercberg, S.; Galan, P.; Group SVMR. Consumption of Dairy Products and Cognitive Functioning: Findings from the SU.VI.MAX 2 Study. J. Nutr. Health Aging 2016, 20, 128–137. [Google Scholar] [CrossRef] [PubMed]
- The ARIC investigators. The Atherosclerosis Risk in Communities (ARIC) Study: Design and objectives. Am. J. Epidemiol. 1989, 129, 687–702. [Google Scholar]
- Knopman, D.S.; Ryberg, S. A verbal memory test with high predictive accuracy for dementia of the Alzheimer type. Arch. Neurol. 1989, 46, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Wechsler, D. Manual for the Wechsler Adult Intelligence Scale-Revised; Psycological Corp: Ney York, NY, USA, 1981. [Google Scholar]
- Benton, A.L.; Eslinger, P.J.; Damasio, A.R. Normative observations on neuropsychological test performances in old age. J. Clin. Neuropsychol. 1981, 3, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, A.M.; Sharrett, A.R.; Schneider, A.L.; Coresh, J.; Albert, M.; Couper, D.; Griswold, M.; Gottesman, R.F.; Wagenknecht, L.E.; Windham, B.G.; et al. Diabetes in midlife and cognitive change over 20 years: A cohort study. Ann. Intern. Med. 2014, 161, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, R.F.; Rawlings, A.M.; Sharrett, A.R.; Albert, M.; Alonso, A.; Bandeen-Roche, K.; Coker, L.H.; Coresh, J.; Couper, D.J.; Griswold, M.E.; et al. Impact of differential attrition on the association of education with cognitive change over 20 years of follow-up: The ARIC neurocognitive study. Am. J. Epidemiol. 2014, 179, 956–966. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.A.; Schneider, J.A.; Buchman, A.S.; Barnes, L.L.; Boyle, P.A.; Wilson, R.S. Overview and findings from the rush Memory and Aging Project. Curr. Alzheimer Res. 2012, 9, 646–663. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.S.; Segawa, E.; Boyle, P.A.; Bennett, D.A. Influence of late-life cognitive activity on cognitive health. Neurology 2012, 78, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- Willett, W.C.; Sampson, L.; Stampfer, M.J.; Rosner, B.; Bain, C.; Witschi, J.; Hennekens, C.H.; Speizer, F.E. Reproducibility and validity of a semiquantitative food frequency questionnaire. Am. J. Epidemiol. 1985, 122, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Steffen, L.M.; Kroenke, C.H.; Yu, X.; Pereira, M.A.; Slattery, M.L.; Van Horn, L.; Gross, M.D.; Jacobs, D.R., Jr. Associations of plant food, dairy product, and meat intakes with 15-year incidence of elevated blood pressure in young black and white adults: The Coronary Artery Risk Development in Young Adults (CARDIA) Study. Am. J. Clin. Nutr. 2005, 82, 1169–1177. [Google Scholar] [PubMed]
- Weng, L.C.; Steffen, L.M.; Szklo, M.; Nettleton, J.; Chambless, L.; Folsom, A.R. A diet pattern with more dairy and nuts, but less meat is related to lower risk of developing hypertension in middle-aged adults: The Atherosclerosis Risk in Communities (ARIC) study. Nutrients 2013, 5, 1719–1733. [Google Scholar] [CrossRef] [PubMed]
- Swallow, D.M. Genetics of lactase persistence and lactose intolerance. Annu. Rev. Genet. 2003, 37, 197–219. [Google Scholar] [CrossRef] [PubMed]
- Tishkoff, S.A.; Reed, F.A.; Ranciaro, A.; Voight, B.F.; Babbitt, C.C.; Silverman, J.S.; Powell, K.; Mortensen, H.M.; Hirbo, J.B.; Osman, M.; et al. Convergent adaptation of human lactase persistence in Africa and Europe. Nat. Genet. 2007, 39, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Mattar, R.; de Campos Mazo, D.F.; Carrilho, F.J. Lactose intolerance: Diagnosis, genetic, and clinical factors. Clin. Exp. Gastroenterol. 2012, 5, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Azur, M.J.; Stuart, E.A.; Frangakis, C.; Leaf, P.J. Multiple imputation by chained equations: What is it and how does it work? Int. J. Methods Psychiatr. Res. 2011, 20, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, A.M.; Sang, Y.; Sharrett, A.R.; Coresh, J.; Griswold, M.; Kucharska-Newton, A.M.; Palta, P.; Wruck, L.M.; Gross, A.L.; Deal, J.A.; et al. Multiple imputation of cognitive performance as a repeatedly measured outcome. Eur. J. Epidemiol. 2016, 32, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Colditz, G.A.; Willett, W.C.; Stampfer, M.J.; Sampson, L.; Rosner, B.; Hennekens, C.H.; Speizer, F.E. The influence of age, relative weight, smoking, and alcohol intake on the reproducibility of a dietary questionnaire. Int. J. Epidemiol. 1987, 16, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Crichton, G.E.; Bryan, J.; Murphy, K.J.; Buckley, J. Review of dairy consumption and cognitive performance in adults: Findings and methodological issues. Dement. Geriatr. Cogn. Disord. 2010, 30, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi, A.; Leighton, H.; Burstein, B.; Xue, X. Latitude, sunshine, and human lactase phenotype distributions may contribute to geographic patterns of modern disease: The inflammatory bowel disease model. Clin. Epidemiol. 2014, 6, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.E.; Coltell, O.; Sorli, J.V.; Estruch, R.; Martinez-Gonzalez, M.A.; Salas-Salvado, J.; Fito, M.; Aros, F.; Dashti, H.S.; Lai, C.Q.; et al. Associations of the MCM6-rs3754686 proxy for milk intake in Mediterranean and American populations with cardiovascular biomarkers, disease and mortality: Mendelian randomization. Sci. Rep. 2016, 6, 33188. [Google Scholar] [CrossRef] [PubMed]
- Willett, W.C. Food Frequency Method Nutritional Epidemiology, 3rd ed.; Oxford University Press: New York, NY, USA, 2012; pp. 70–141. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Covariates | Milk Intake Group | |||
|---|---|---|---|---|
| Almost Never | <1 Glass/Day | 1 Glass/Day | >1 Glass/Day | |
| n = 1554 | n = 6872 | n = 2036 | n = 3290 | |
| Age, mean (SD) | 56.7 (5.6) | 57.2 (5.6) | 58.5 (5.7) | 57.9 (5.8) |
| Black, % | 530 (34.1%) | 1833 (26.7%) | 360 (17.7%) | 542 (16.5%) |
| Female, % | 1023 (65.8%) | 3879 (56.4%) | 1096 (53.8%) | 1664 (50.6%) |
| Study site, % | ||||
| Forsyth County, NC | 328 (21.1%) | 1894 (27.6%) | 584 (28.7%) | 760 (23.1%) |
| Jackson, MS | 484 (31.1%) | 1641 (23.9%) | 320 (15.7%) | 469 (14.3%) |
| Minneapolis, MN | 335 (21.6%) | 1499 (21.8%) | 601 (29.5%) | 1293 (39.3%) |
| Washington County, MD | 407 (26.2%) | 1838 (26.7%) | 531 (26.1%) | 768 (23.3%) |
| Education, % <High School | 415 (26.8%) | 1474 (21.5%) | 390 (19.2%) | 627 (19.1%) |
| Smoking, % Never | 564 (36.3%) | 2771 (40.3%) | 839 (41.2%) | 1301 (39.6%) |
| Drinking, % Never | 366 (23.6%) | 1582 (23.0%) | 476 (23.4%) | 654 (19.9%) |
| BMI (kg/m2), mean (SD) | 27.9 (5.7) | 28.1 (5.5) | 27.7 (5.1) | 27.9 (5.1) |
| Diabetes, % | 220 (14.3%) | 945 (13.8%) | 311 (15.3%) | 555 (16.9%) |
| Hypertension, % | 622 (40.2%) | 2451 (35.7%) | 721 (35.6%) | 1076 (32.8%) |
| Diet score, mean (SD) | 19.3 (4.9) | 20.7 (4.7) | 22.0 (4.7) | 22.1(4.8) |
| Lactose intake (g), mean (SD) | 2.3 (3.0) | 7.7 (5.5) | 14.9 (3.5) | 27.8 (15.4) |
| Physical activity (met-min/week) | 500 (647) | 674 (825) | 822 (907) | 728 (782) |
| APOEe4 allele, % present | 565 (33.8%) | 2218 (30.2%) | 669 (30.7%) | 1071 (30.3%) |
| Lactase persistence (Whites) | ||||
| CC (Lactase non-persistent) | 149 (17.0%) | 444 (10.1%) | 96 (6.5%) | 139 (5.8%) |
| CT (Lactase persistent) | 326 (37.1%) | 1722 (39.2%) | 589 (39.5%) | 922 (38.2%) |
| TT (Lactase persistent) | 403 (45.9%) | 2224 (50.7%) | 803 (54.0%) | 1355 (56.1%) |
| Cognitive test scores | ||||
| DWRT, mean (SD) | 6.6 (1.5) | 6.6 (1.5) | 6.5 (1.5) | 6.6 (1.5) |
| DSST, mean (SD) | 42.6 (15.2) | 44.6 (14.4) | 45.2 (13.7) | 45.6 (13.6) |
| WFT, mean (SD) | 31.2 (12.9) | 33.4 (12.4) | 33.5 (12.6) | 33.8 (12.3) |
| Test | 20-Year Decline | Difference | Percent |
|---|---|---|---|
| Global z | |||
| Almost never | −0.94 (−1.00, −0.88) | ref | ref |
| <1 glass/day | −0.99 (−1.01, −0.96) | −0.05 (−0.11, 0.02) | 5% |
| 1 glass/day | −1.00 (−1.05, −0.95) | −0.06 (−0.13, 0.02) | 6% |
| >1 glass/day | −1.04 (−1.08, −1.01) | −0.10 (−0.16, −0.03) | 11% |
| DWRT z | |||
| Almost never | −1.15 (−1.23, −1.06) | ref | ref |
| <1 glass/day | −1.19 (−1.23, −1.15) | −0.04 (−0.13, 0.06) | 3% |
| 1 glass/day | −1.18 (−1.26, −1.11) | −0.03 (−0.14, 0.08) | 3% |
| >1 glass/day | −1.25 (−1.31, −1.19) | −0.10 (−0.20, 0.00) | 9% |
| DSST z | |||
| Almost never | −0.78 (−0.82, −0.74) | ref | ref |
| <1 glass/day | −0.82 (−0.84, −0.80) | −0.04 (−0.09, 0.00) | 5% |
| 1 glass/day | −0.85 (−0.89, −0.81) | −0.07 (−0.12, −0.01) | 9% |
| >1 glass/day | −0.87 (−0.89, −0.84) | −0.09 (−0.14, −0.03) | 12% |
| WFT z | |||
| Almost never | −0.24 (−0.29, −0.19) | ref | ref |
| <1 glass/day | −0.28 (−0.30, −0.26) | −0.04 (−0.09, 0.02) | 16% |
| 1 glass/day | −0.26 (−0.30, −0.22) | −0.02 (−0.08, 0.05) | 8% |
| >1 glass/day | −0.29 (−0.33, −0.26) | −0.05 (−0.11, 0.01) | 21% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petruski-Ivleva, N.; Kucharska-Newton, A.; Palta, P.; Couper, D.; Meyer, K.; Graff, M.; Haring, B.; Sharrett, R.; Heiss, G. Milk Intake at Midlife and Cognitive Decline over 20 Years. The Atherosclerosis Risk in Communities (ARIC) Study. Nutrients 2017, 9, 1134. https://doi.org/10.3390/nu9101134
Petruski-Ivleva N, Kucharska-Newton A, Palta P, Couper D, Meyer K, Graff M, Haring B, Sharrett R, Heiss G. Milk Intake at Midlife and Cognitive Decline over 20 Years. The Atherosclerosis Risk in Communities (ARIC) Study. Nutrients. 2017; 9(10):1134. https://doi.org/10.3390/nu9101134
Chicago/Turabian StylePetruski-Ivleva, Natalia, Anna Kucharska-Newton, Priya Palta, David Couper, Katie Meyer, Misa Graff, Bernhard Haring, Richey Sharrett, and Gerardo Heiss. 2017. "Milk Intake at Midlife and Cognitive Decline over 20 Years. The Atherosclerosis Risk in Communities (ARIC) Study" Nutrients 9, no. 10: 1134. https://doi.org/10.3390/nu9101134
APA StylePetruski-Ivleva, N., Kucharska-Newton, A., Palta, P., Couper, D., Meyer, K., Graff, M., Haring, B., Sharrett, R., & Heiss, G. (2017). Milk Intake at Midlife and Cognitive Decline over 20 Years. The Atherosclerosis Risk in Communities (ARIC) Study. Nutrients, 9(10), 1134. https://doi.org/10.3390/nu9101134