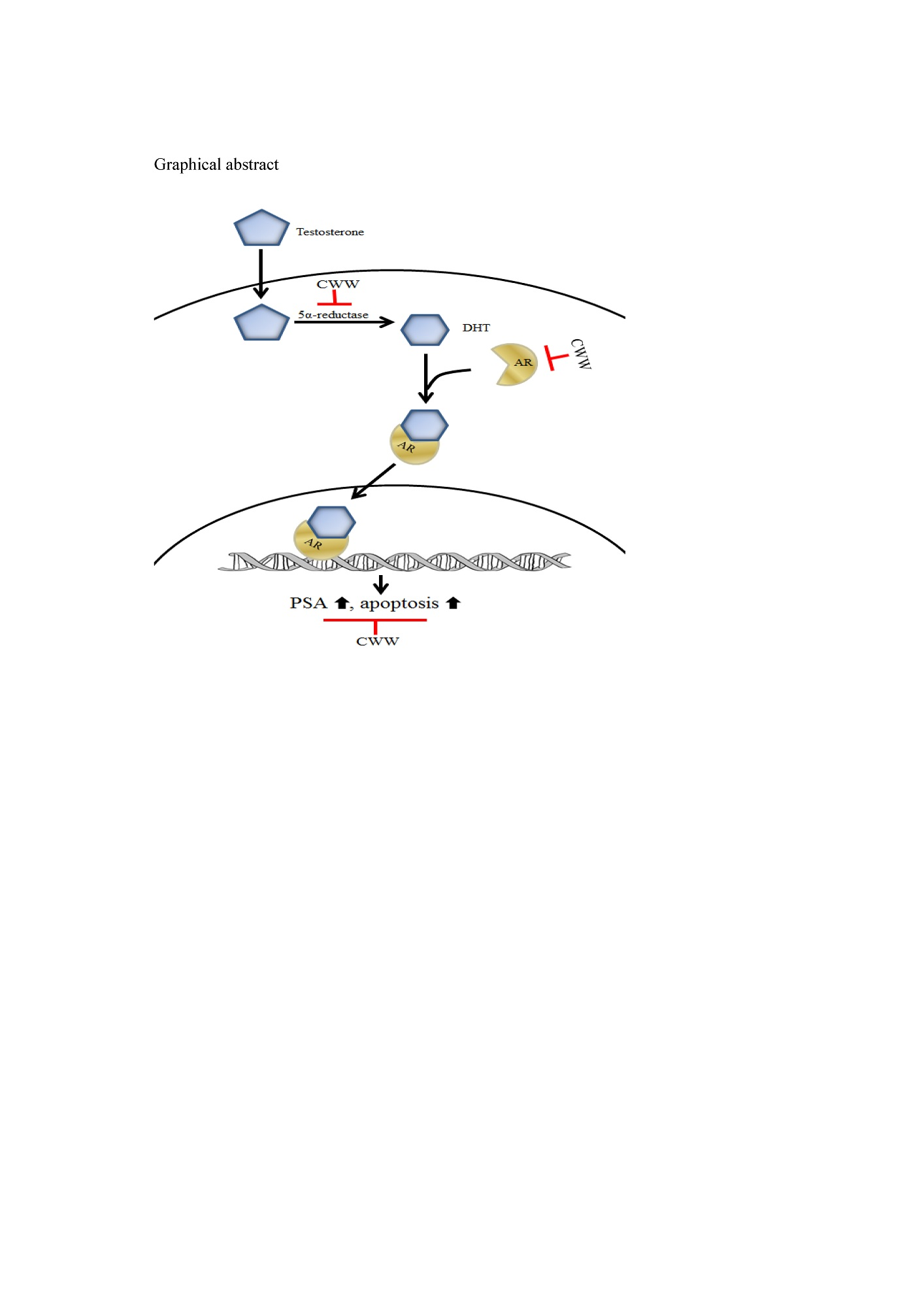
Cynanchum wilfordii Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia by Regulating 5α-Reductase and Androgen Receptor Activities in a Rat Model

Abstract

1. Introduction
2. Materials and Methods
2.1. Sample Procurement
2.2. Experimental Animals and Maintenance Conditions
2.3. Orchiectomy Procedure
2.4. Induction of BPH
2.5. Sample Administration Groups
2.6. Prostate Weight
2.7. Determination of Testosterone and DHT Levels in Serum and Prostate
2.8. Determination of Prostate-Specific Antigen (PSA) Levels in Serum
2.9. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis of AR, 5α-Reductase, and Bcl-2
2.10. Histological Examination
2.11. Measurement of Serum Alanine Aminotransferase (ALT) and Aspartate Aminotransferase (AST) Levels
2.12. Standardization of CWW
2.13. Statistical Analysis
3. Results
3.1. Effects of CWW on Body and Prostate Weights
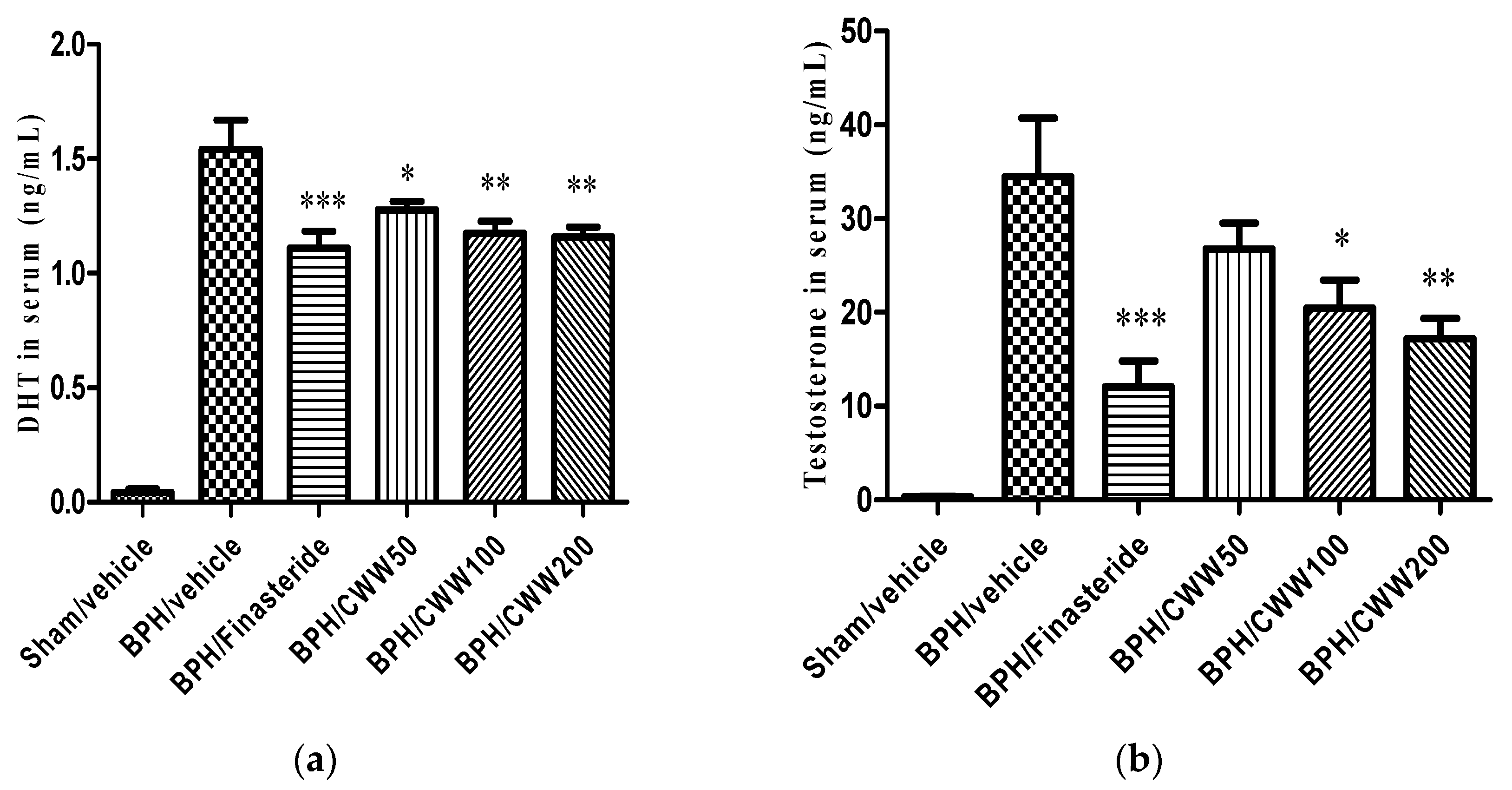
3.2. Effects of CWW on Serum Testosterone and DHT Levels
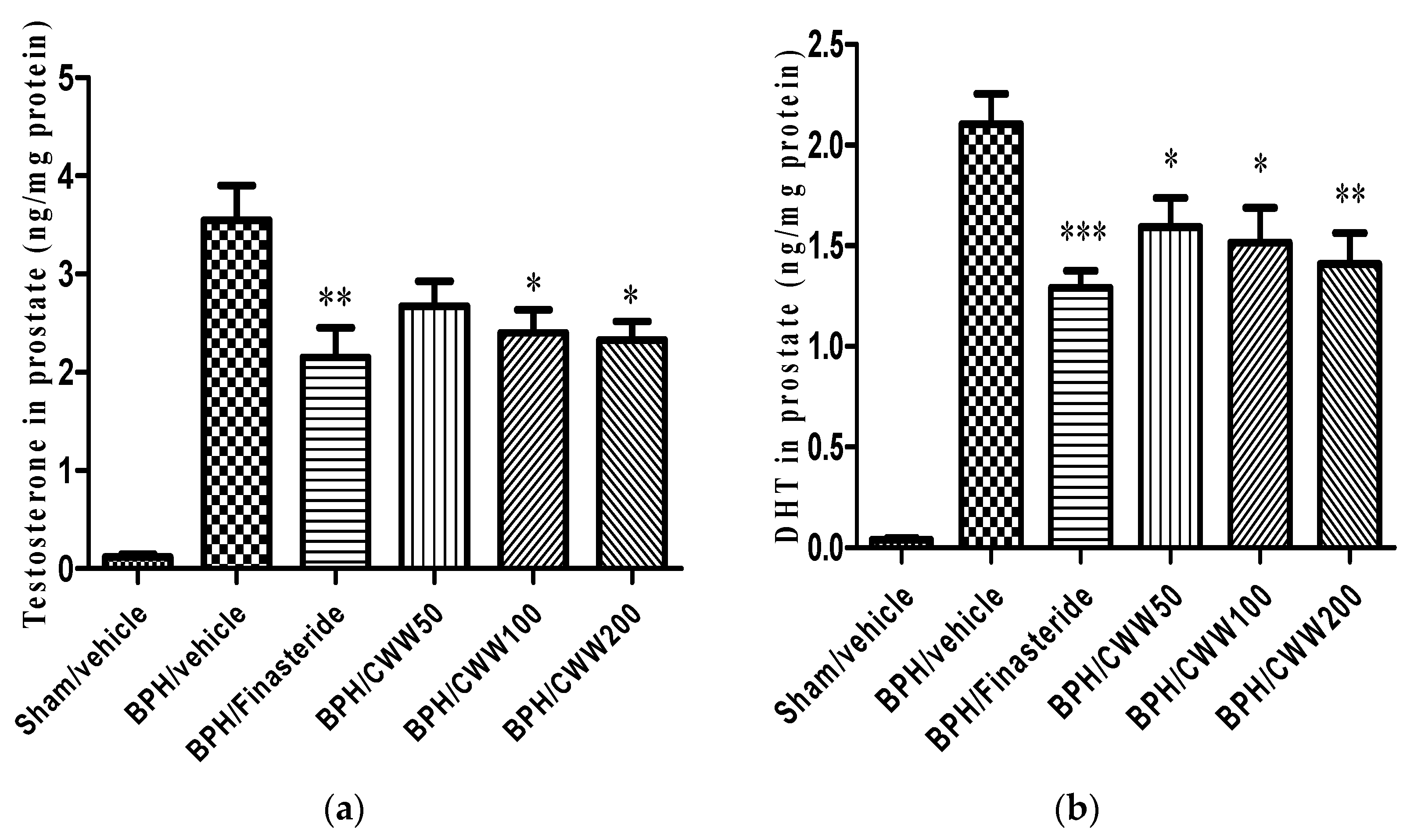
3.3. Effects of CWW on Prostate Testosterone and DHT Levels
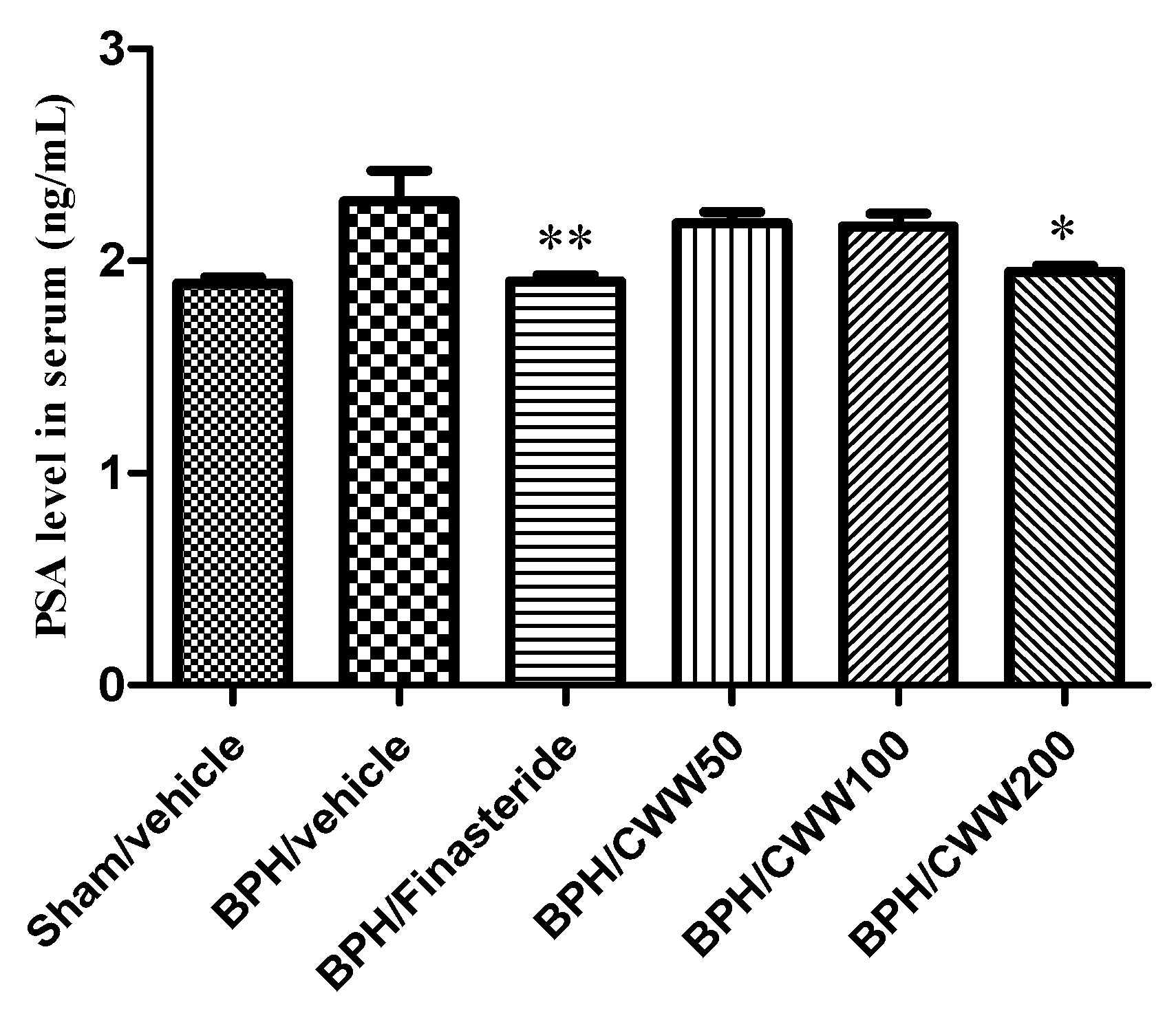
3.4. Effects of CWW on Serum PSA Levels
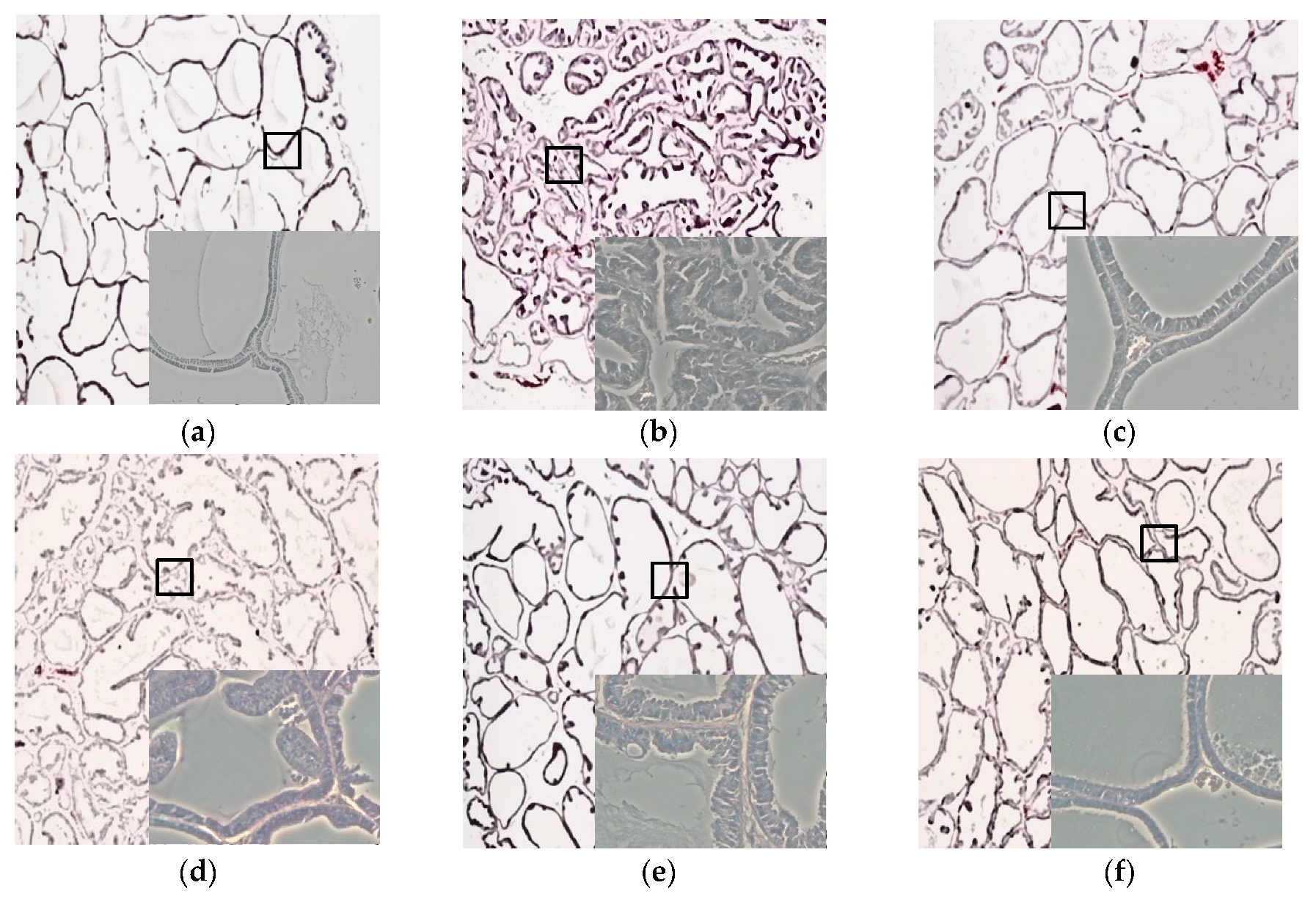
3.5. Effects of CWW on Prostate Tissue Histology
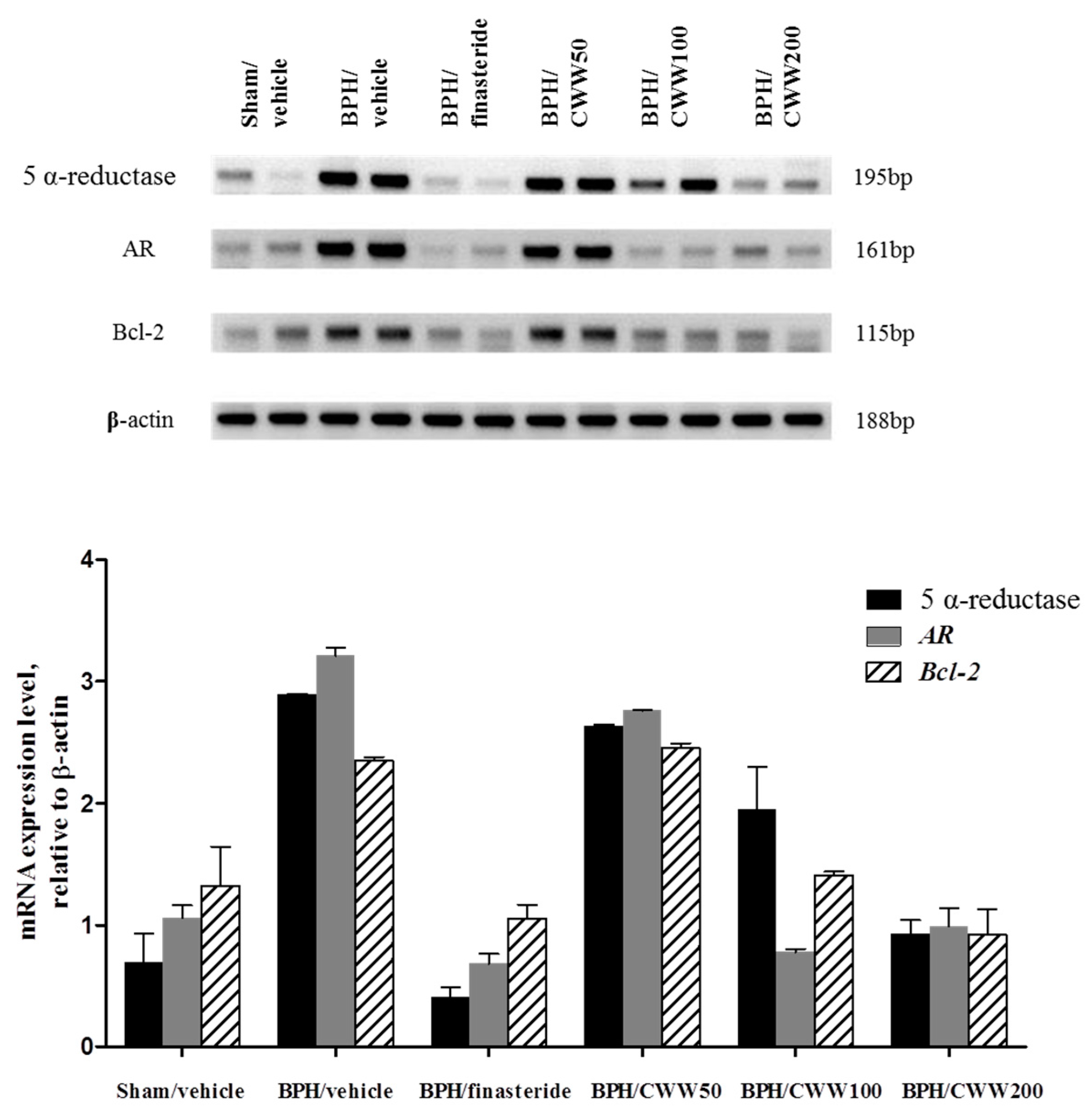
3.6. Assessment of AR, 5α-Reductase, and Bcl-2 mRNA Expression
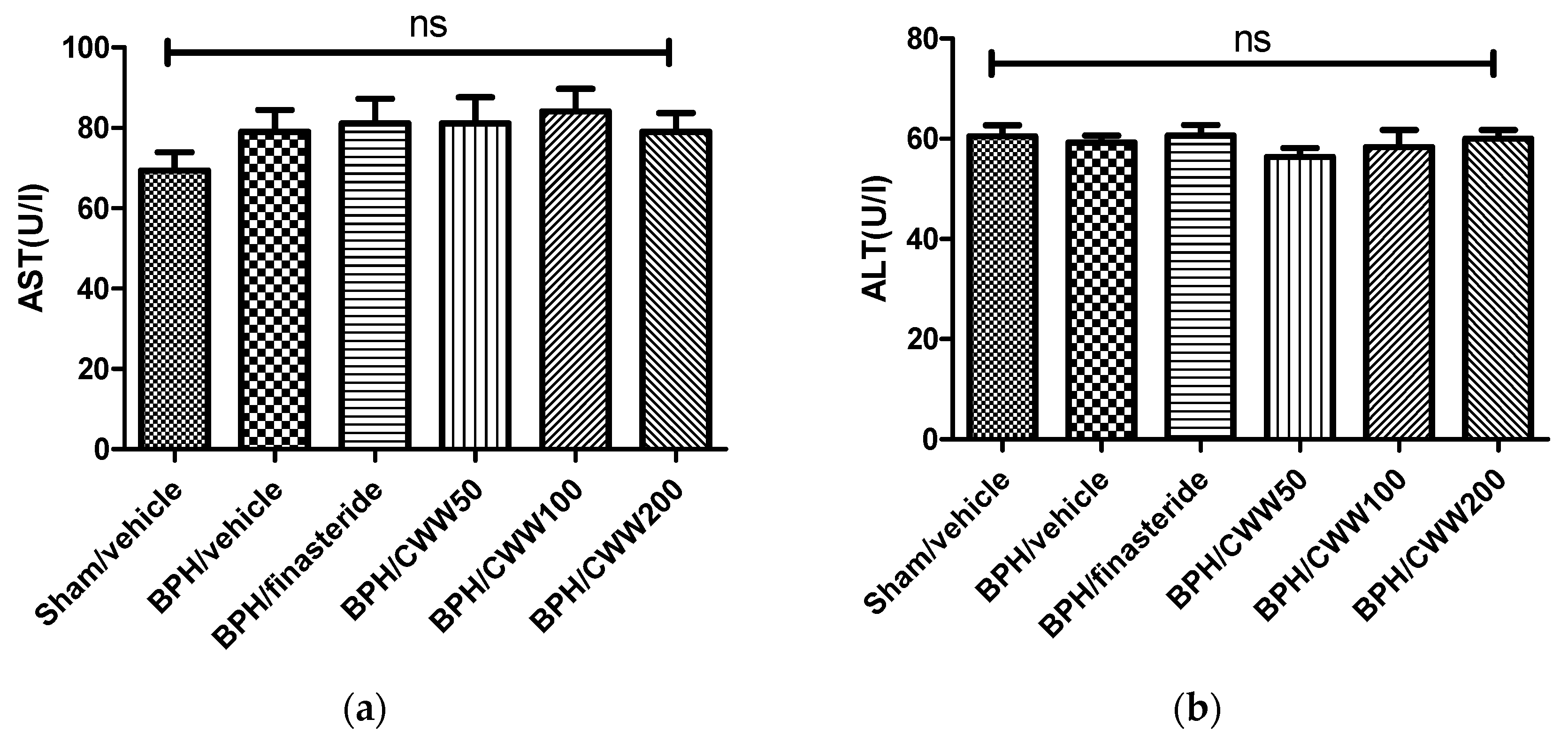
3.7. Toxicity of CWW in Rat BPH Model
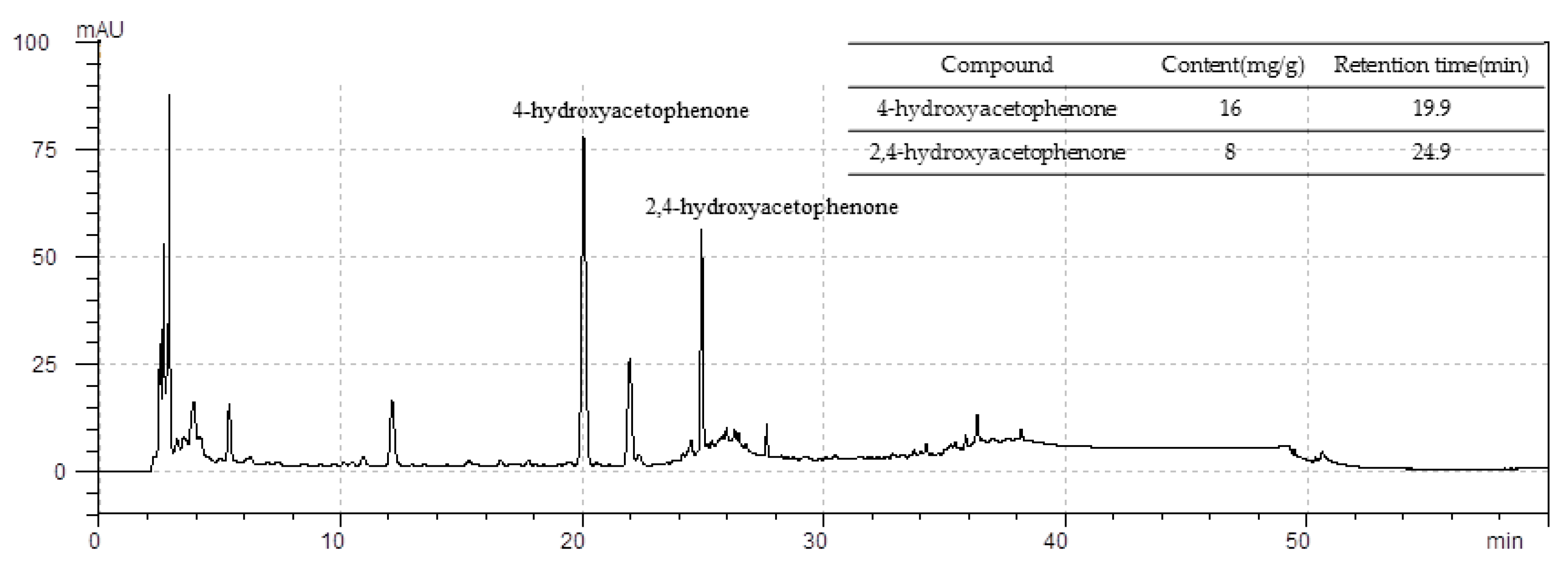
3.8. Standardization of CWW
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Garraway, W.M.; Collins, G.N.; Lee, R.J. High prevalence of benign prostatic hypertrophy in the community. Lancet 1991, 338, 469–471. [Google Scholar] [CrossRef]
- Chute, C.G.; Panser, L.A.; Girman, C.J.; Osterling, J.E.; Guess, H.A.; Jacobsen, S.J.; Lieber, M.M. The prevalence of prostatism: A population-based survey of urinary symptoms. J. Urol. 1993, 150, 85–89. [Google Scholar] [CrossRef]
- Foster, C.S. Pathology of benign prostatic hyperplasia. Prostate Suppl. 2000, 9, 4–14. [Google Scholar] [CrossRef]
- Schauer, I.G.; Rowley, D.R. The functional role of reactive stroma in benign prostatic hyperplasia. Differentiation 2011, 8, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Shoskes, D.A. Phytotherapy in chronic proststitis. Urology 2002, 60, 35–37. [Google Scholar] [CrossRef]
- Huhtaniemi, I.; Forti, G. Male late-onset hypogonadism: Pathogenesis, diagnosis and treatment. Nat. Rev. Urol. 2011, 8, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Huhtaniemi, I. Late-onset hypogonadism: Current concepts and controversies of pathogenesis, diagnosis and treatment. Asian J. Androl. 2014, 16, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, G.; Rittmaster, R.S.; Klocker, H. Dihydrotestosterone and the concept of 5α-reductase inhibition in human benign prostatic hyperplasia. Eur. Urol. 2000, 37, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Roehrborn, C.G. Male lower urinary tract symptoms (LUTS) and benign prostatic hyperplasia (BPH). Med. Clin. N. Am. 2011, 95, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Speakman, M.; Kirby, R.; Doyle, S.; Ioannou, C. Burden of male lower urinary tract symptoms (LUTS) suggestive of benign prostatic hyperplasia (BPH)-Focus on the UK. Br. J. Urol. 2014, 115, 508–519. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.L.; Wahane, V.D. Current status of 5α-reductase inhibitors in the treatment of benign hyperplasia of prostate. Indian J. Med. Sci. 2008, 62, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Gravas, S.; Oelke, M. Current status of 5α-reductase inhibitors in the management of lower urinary tract symptoms and BPH. World J. Urol. 2010, 28, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Bullock, T.L.; Andriole, G.L. Emerging drug therapies for benign prostatic hyperplasia. Expert Opin. Emerg. Drugs 2006, 11, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Paba, S.; Frau, R.; Godar, S.S.; Devot, P.; Marrosu, F.; Bortolato, M. Steroid 5α-reductase as a novel therapeutic target for schizophrenia and other neuropsychiatric disorders. Curr. Pharm. Des. 2011, 17, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Berges, R.R.; Furuya, Y.; Remington, L.; English, H.F.; Jacks, T.; Isaacs, J.T. Cell proliferation, DNA repair, and p53 function are not required for programmed death of prostatic glandular cells induced by androgen ablation. Proc. Natl. Acad. Sci. USA 1993, 90, 8910–8914. [Google Scholar] [CrossRef] [PubMed]
- English, H.F.; Drago, J.R.; Santen, R.J. Cellular response to androgen depletion and repletion in the rat ventral prostate: Autoradiography and morphometric analysis. Prostate 1985, 7, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Sandford, N.L.; Searle, J.W.; Kerr, J.F.R. Successive waves of apoptosis in the rat prostate after repeated withdrawal of testosterone stimulation. Pathology 1984, 16, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Buttyan, R.; Shabsigh, A.; Perlman, H.; Colombel, M. Regulation of apoptosis in the prostate gland by androgenic steroids. Trends Endocrinol. Metab. 1999, 10, 47–54. [Google Scholar] [CrossRef]
- Yang, S.B.; Lee, S.M.; Park, J.; Lee, T.H.; Baek, N.; Park, H.; Lee, H.; Kim, J. Cynandione A from Cynanchum wilfordii attenuates the production of inflammatory mediators in LPS-induced BV-2 microglial cells via NF-κB inactivation. Biol. Pharm. Bull. 2014, 37, 1390–1396. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.; Kwak, B.Y.; Yi, K.; Kim, J.S. The effect of herbal extract (EstroG-100) on pre-, peri- and post-menopausal women: A randomized double-blind placebo-controlled study. Phytother. Res. 2012, 26, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.O.; Kim, J.Y.; Pan, S.O.; Kim, M.; Jun, W.J.; Choi, C.Y. Aphrodisiac property of the aqueous extract of Cynanchum wilfordii. J. Food Nutr. Res. 2016, 4, 713–719. [Google Scholar] [CrossRef]
- Atawia, R.T.; Tadros, M.G.; Khalifa, A.E.; Mosli, H.A.; Abdel-Naim, A.B. Role of the phytoestrogenic, pro-apoptotic and anti-oxidative properties of silymarin in inhibiting experimental benign prostatic hyperplasia in rats. Toxicol. Lett. 2013, 219, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Huynh, H. Induction of apoptosis in rat ventral prostate by finasteride is associated with alteration in MAP kinase pathways and Bcl-2 related family of proteins. Int. J. Oncol. 2002, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Donnell, R.F. Benign prostate hyperplasia: A review of the year’s progress from bench to clinic. Curr. Opin. Urol. 2011, 21, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Culig, Z.; Hobisch, A.; Cronauer, M.V.; Radmayr, C.; Hittmair, A.; Zhang, J.; Thurnher, M.; Bartsch, G.; Klocker, H. Regulation of prostatic growth and function by peptide growth factors. Prostate 1996, 28, 392–405. [Google Scholar] [CrossRef]
- Jacobsen, S.J.; Girman, C.J.; Lieber, M.M. Natural history of benign prostatic hyperplasia. Urology 2001, 58, 5–16. [Google Scholar] [CrossRef]
- Shin, I.S.; Lee, M.Y.; Jung, D.Y.; Seo, C.S.; Ha, H.K.; Shin, H.K. Ursolic acid reduces prostate size and dihydrotestosterone level in a rat model of benign prostatic hyperplasia. Food Chem. Toxicol. 2012, 50, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Andriole, G.; Bruchovsky, N.; Chung, L.W.; Matsumoto, A.M.; Rittmaster, R.; Roehrborn, C.; Russell, D.; Tindall, D. Dihydrotestosterone and the prostate: The scientific rationale for 5α-reductase inhibitors in the treatment of benign prostatic hyperplasia. J. Urol. 2004, 172, 1399–1403. [Google Scholar] [CrossRef] [PubMed]
- Martikainen, P.; Kyprianou, N.; Isaacs, J.T. Effect of transforming growth factor-β on proliferation and death of rat prostatic cells. Endocrinology 1990, 127, 2963–2968. [Google Scholar] [CrossRef] [PubMed]
- Arias, E. United States life tables, 2007. Natl. Vital Stat. Rep. 2011, 59, 1–60. [Google Scholar] [PubMed]
- Roehrborn, C.G. Pathology of benign prostatic hyperplasia. Int. J. Impot. Res. 2008, 20, S11–S18. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Tarter, T.H. Combination therapy with dutasteride and tamsulosin for the treatment of symptomatic enlarged prostate. Clin. Interv. Aging 2009, 4, 251–258. [Google Scholar] [PubMed]
- Nickel, J.C.; Gilling, P.; Tammela, T.L.; Morrill, B.; Wilson, T.H.; Rittmaster, R.S. Comparison of dutasteride and finasteride for treating benign prostatic hyperplasia the enlarged prostate international comparator study (EPICS). BJU Int. 2011, 108, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Bent, S.; Kane, C.; Shinohara, K.; Neuhaus, J.; Hudes, E.S.; Goldberg, H.; Avins, A.L. Saw palmetto for benign prostatic hyperplasia. N. Engl. J. Med. 2006, 354, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Okazaki, H.; Ono, Y.; Kurokawa, K.; Suzuki, T.; Onuma, E.; Takanashi, H.; Mamiya, Y.; Yamanaka, H. Effect of dual inhibition of 5α-reductase and aromatase on spontaneously developed canine prostatic hypertrophy. Prostate 1998, 37, 70–76. [Google Scholar] [CrossRef]
- Izumi, K.; Mizokami, A.; Lin, W.J.; Lai, K.P.; Chang, C. Androgen receptor roles in the development of benign prostate hyperplasia. Am. J. Pathol. 2013, 182, 1942–1949. [Google Scholar] [CrossRef] [PubMed]
- Sciarra, A.; Mariotti, G.; Salciccia, S.; Autran, G.A.; Monti, S.; Toscano, V.; Di, S.F. Prostate growth and inflammation. J. Steroid Biochem. Mol. Biol. 2008, 108, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Luo, F.; Zhou, Y.; Du, X.; Shi, J.; Zhao, X.; Xu, Y.; Zhu, Y.; Hong, W.; Zhang, J. The therapeutic effects of docosahexaenoic acid on oestrogen/androgen-induced benign prostatic hyperplasia in rats. Exp. Cell Res. 2016, 345, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Lilja, H. Biology of prostate-specific antigen. Urology 2003, 62, 27–33. [Google Scholar] [CrossRef]
- Levitt, J.M.; Slawin, K.M. Prostate-specific antigen and prostate-specific antigen derivatives as predictors of benign prostatic hyperplasia progress. Curr. Urol. Rep. 2007, 8, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Rick, F.G.; Schally, A.V.; Block, N.L.; Halmos, G.; Perez, R.; Fernandez, J.B.; Vidaurre, I.; Szalontay, L. LHRH antagonist Cetrorelix reduces prostate size and gene expression of proinflammatory cytokines and growth factors in a rat model of benign prostatic hyperplasia. Prostate 2011, 71, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.S.; Lim, H.; Moon, S.; Oh, H.L.; Kim, Y.T.; Kim, M.K.; Lee, C.H. Benzyldihydroxyoctenone, a novel anticancer agent, induces apoptosis via mitochondrial-mediated pathway in androgen-sensitive LNCap prostate cancer cells. Bioorg. Med. Chem. Lett. 2009, 19, 742–744. [Google Scholar] [CrossRef] [PubMed]
- Buttyan, R.; Zakeri, Z.; Lockshin, R.; Wolgemuth, D. Cascade induction of c-fos, c-myc, and heat shock 70K transcripts during regression of the rat ventral prostate gland. Mol. Endocrinol. 1988, 2, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Marti, A.; Jehn, B.; Costello, E.; Keon, N.; Ke, G.; Martin, F.; Jaqqi, R. Protein kinase A and AP-1 (c-Fos/JunD)are induced during apoptosis of mouse mammary epithelial cells. Oncogene 1994, 9, 1213–1223. [Google Scholar] [PubMed]
- Colombel, M.; Radvanyi, F.; Blanche, M.; Abbou, C.; Buttyan, R.; Donehower, L.A.; Chopin, D.; Thiery, J.P. Androgen suppressed apoptosis is modified in p53 deficient mice. Oncogene 1995, 10, 1269–1274. [Google Scholar] [PubMed]
- Perlman, H.; Zhang, X.; Chen, M.W.; Walsh, K.; Buttyan, R. An elevated bax/bcl-2 ratio corresponds with the onset of prostate epithelial cell apoptosis. Cell Death Differ. 1999, 6, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Matsuzawa, A.; Iguchi, T. Down regulation of Bcl-2 is the first step on Fas-mediated apoptosis of male reproductive tract. Oncogene 1996, 13, 31–37. [Google Scholar] [PubMed]
- Dimmeler, S.; Breitschopf, K.; Haendeler, J.; Zeiher, A.M. Dephosphorylation targets Bcl-2 for ubiquitin-dependent degradation: A link between the apoptosome and the proteasome pathway. J. Exp. Med. 1999, 189, 1815–1822. [Google Scholar] [CrossRef] [PubMed]
- Majno, G.; Joris, I. Apoptosis, oncosis, and necrosis. An overview of cell death. Am. J. Pathol. 1995, 146, 3–15. [Google Scholar] [PubMed]
- Kyprianou, N.; Tu, H.; Jacobs, S.C. Apoptotic versus proliferative activities in human benign prostatic hyperplasia. Hum. Pathol. 1996, 27, 668–675. [Google Scholar] [CrossRef]
- Shariat, S.F.; Ashfaq, R.; Roehrborn, C.G.; Slawin, K.M.; Lotan, Y. Expression of survivin and apoptotic biomarkers in benign prostatic hyperplasia. J. Urol. 2005, 174, 2046–2050. [Google Scholar] [CrossRef] [PubMed]
- Yeo, H.S.; Kim, J.W. A benzoquinone from Cynanchum wilfordii. Phytochemistry 1997, 46, 1103–1105. [Google Scholar] [CrossRef]
- Kapoor, A. Benign prostatic hyperplasia (BPH) management in the primary care setting. Can. J. Urol. 2012, 19 (Suppl. 1), 10–17. [Google Scholar] [PubMed]
- Lepor, H.; Kazzazi, A.; Djavan, B. Alpha-Blockers for benign prostatic hyperplasia: The new era. Curr. Opin. Urol. 2012, 22, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Chughtai, B.; Forde, J.C.; Thomas, D.D.; Laor, L.; Hossack, T.; Woo, H.H.; Te, A.E.; Kaplan, S.A. Benign prostatic hyperplasia. Nat. Rev. Dis. Prim. 2016, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Emberton, M.; Cornel, E.B.; Bassi, P.F.; Fourcade, R.O.; Gomez, J.M.F.; Castro, R. Benign prostatic hyperplasia as a progressive disease: A guide to the risk factors and options for medical management. Int. J. Clin. Pract. 2008, 62, 1076–1086. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Body Weight Gain (g) | Prostate Weight (g) | Growth Inhibition (%) |
|---|---|---|---|
| Sham/vehicle | 118.5 ± 5.0 | 0.82 ± 0.07 | -- |
| BPH/vehicle | 41.5 ± 4.4 | 1.39 ± 0.07 | -- |
| BPH/finasteride | 38.8 ± 4.2 | 1.06 ± 0.06 ** | 58.1 ± 10.7 ** |
| BPH/CWW 50 | 33.6 ± 5.0 | 1.08 ± 0.07 * | 54.5 ± 11.7 * |
| BPH/CWW 100 | 39.9 ± 6.6 | 1.10 ± 0.05 * | 51.8 ± 9.7 * |
| BPH/CWW 200 | 41.9 ± 6.0 | 1.11 ± 0.08 * | 50.1 ± 14.4 * |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, G.; Shin, J.; Choi, H.; Jo, A.; Pan, S.; Bae, D.; Lee, Y.; Choi, C. Cynanchum wilfordii Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia by Regulating 5α-Reductase and Androgen Receptor Activities in a Rat Model. Nutrients 2017, 9, 1070. https://doi.org/10.3390/nu9101070
Lee G, Shin J, Choi H, Jo A, Pan S, Bae D, Lee Y, Choi C. Cynanchum wilfordii Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia by Regulating 5α-Reductase and Androgen Receptor Activities in a Rat Model. Nutrients. 2017; 9(10):1070. https://doi.org/10.3390/nu9101070
Chicago/Turabian StyleLee, Gyuok, Jawon Shin, Hakjoon Choi, Ara Jo, SangO Pan, Donghyuck Bae, Yongwook Lee, and Chulyung Choi. 2017. "Cynanchum wilfordii Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia by Regulating 5α-Reductase and Androgen Receptor Activities in a Rat Model" Nutrients 9, no. 10: 1070. https://doi.org/10.3390/nu9101070
APA StyleLee, G., Shin, J., Choi, H., Jo, A., Pan, S., Bae, D., Lee, Y., & Choi, C. (2017). Cynanchum wilfordii Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia by Regulating 5α-Reductase and Androgen Receptor Activities in a Rat Model. Nutrients, 9(10), 1070. https://doi.org/10.3390/nu9101070